
Intelligentes Design der Lebewesen, Konvergenzen (unabhängige Merkmalsentstehung) & Grundtypenbiologie
04.02.21 Wie fliegt ein Schmetterling?
Die flatternden Sommervögel haben unter den Insekten einen ganz eigenen Flugstil. Mit Ihren großflächigen Flügeln scheinen sie eher ziellos und zufällig hin und her zu flattern. Wie aber funktioniert der Flug der Schmetterlinge? Wie erzeugen die Vor- und Auftrieb? Interessante Experimente geben Einblicke in einige der zugrundeliegenden aerodynamischen Prozesse.
Ein Schwalbenschwanz (Papilio machaon) der eben noch auf einer Blüte ruhte, hat sich mit wenigen Flügelschlägen erhoben und flattert mit eingestreuten Segeleinlagen dem Wegrain entlang. Wie erheben sich diese filigranen Insekten in die Luft und nach welchen Prinzipien funktioniert diese zufällig scheinende Luftfahrt? Diese Frage kann man nach derzeitigem Wissenstand nur ansatzweise beantworten; hier werden einige Beobachtungen und Einsichten zusammengestellt, die zum Nachdenken anregen und zu eigenem Beobachten ermutigen sollen.

Abb. 1 Schwalbenschwanz (Papilio machaon). (Wikimedia: hamon jp, CC BY-SA 3.0)
Otto Liliental, einer der Flugpioniere, hat gemeinsam mit seinem Bruder Gustav bereits als Schüler mit intensiven Studien des Vogelflugs begonnen. Daraus haben sie Modelle zum Schlagflug (Fliegen mit sich auf und ab bewegenden Flügeln) entwickelt und dazu auch experimentelle Untersuchungen durchgeführt. Schließlich entwickelten sie erste Flugapparate mit starren und gewölbten Tragflächen, mit denen sie den Gleitflug großer Vögel nachahmen konnten (Lilienthal 1889). Bis heute ist der Vogelflug Gegenstand wissenschaftlicher Untersuchungen, weil man sich noch viele spannende und technisch nutzbare Entdeckungen verspricht.
Der Insektenflug unterscheidet sich vom Vogelflug nicht nur aufgrund der unterschiedlichen Dimensionen an Masse und der Größe sowie an den eingesetzten Materialien; dabei kommen auch andere aerodynamische Prinzipen ins Blickfeld und gewinnen an Einfluss. Die hauptsächlich aus Chitin1 bestehenden flexiblen Insektenflügel werden typischerweise mit hoher Frequenz geschlagen und damit Vor- und Auftrieb erzeugt. Schwebfliegen und Libellen sind mit ihren nicht durch Muskelkraft aktiv veränderbaren Flügeln zu ganz erstaunlichen und faszinierenden Flugmanövern fähig.
Die meisten Schmetterlinge weichen in ihrem typischen Körperbau von dem anderer Insekten ab. Ein schmaler kleiner Körper trägt vier großflächige Flügel, die in der Regel auf jeder Seite paarweise synchron auf und ab bewegt werden. Die aus einer Chitin-Doppelmembran bestehenden Flügel sind mit Adern durchzogen und mit einer unüberschaubar großen Zahl winziger Schuppen bedeckt. Diese sind nur lose an der Chitin-Membran angeheftet. Im Flug nutzen Schmetterlinge ihre Flügel typischerweise mit einer niedrigen Frequenz von z. B. 15 Hz. Das Taubenschwänzchen (Macroglossum stellatarum), das im Schwirrflug, wie ein Kolibri an einer Blüte steht, schlägt die Flügel mit ca. 80 Hz. Die meisten Schmetterlinge zeigen auch einen typisch schaukelnden Flug, der auf den ersten Blick wenig zielgerichtet und effizient erscheint (obwohl Wanderfalter, wie z. B. der Monarchfalter [Danaus plexippus] große Strecken mit extremer Effizienz zurücklegen können).
Wie also fliegt ein Schmetterling, was sind die dabei genutzten Mechanismen? Mit großem experimentellem Aufwand untersuchten Fuchiwaki et al. (2013) mit an den Beinen auf einem kleinen Träger fixierte Distelfalter (Cynthia cardui) und Weiße Baumnymphen (Idea leuconoe) im Windkanal. Dabei kamen Hochgeschwindigkeitskameras, Laserlichtquellen und sehr leichte, kleinste Kunststoffkügelchen (10µm Durchmesser) zum Einsatz, um Luftwirbel dokumentieren zu können. Die an technischen Instituten in Japan arbeitenden Wissenschaftler wollten Informationen zur Verbesserung technischer Kleinstfluggeräte wie Mini-Drohnen gewinnen. Sie konnten – mit erheblichem mathematischem Aufwand – dokumentieren, dass sich beim Auf- und Abschlag der Schmetterlingsflügel an den Flügelkanten ringförmig geschlossene, schlauchartige Wirbel ausbildeten, die sich an den Umkehrpunkten der Flügel von denselben ablösten. Das war bei beiden Schmetterlingsarten gleich, die Form der Wirbelschläuche war aber entsprechend der unterschiedlichen Flügelgröße und -umrisse verändert. Diese Luftwirbelschläuche an den Flügelkanten tragen nach den Erkenntnissen der Autoren mit zum Flugverhalten der Falter bei.
Johansson & Henningsson, zwei schwedische Biowissenschaftler eines Ökologischen Instituts, untersuchten in einem vergleichbaren Experiment frei fliegende Exemplare des Kaisermantels (Argynnis paphia) im Windkanal. Dabei kamen mehrere Hochgeschwindigkeitskameras zum Einsatz, was einen entsprechend höheren Rechenaufwand erforderte. Bei dieser Studie (Johansson & Henningsson 2021) starteten die Schmetterlinge im Windkanal von einer Startplattform aus gegen einen Luftstrom von ca. 2 m s-1. Beim Aufschlag der Flügel bilden die flexiblen Flügelflächen, kurz bevor sie oben zusammenschlagen, eine tassenförmige Struktur, deren Öffnung nach hinten gerichtet ist. Wenn die Flügelflächen aufeinander klatschen, ergibt sich ein nach hinten gerichteter Luftstrom, der für den Schmetterling einen Vortrieb bewirkt. Die Untersuchung ergab, dass der Flügelaufschlag bei Kaisermantel in der allerletzten Phase, kurz vor dem Zusammenklatschen der Flügel, den Vortrieb erzeugt. Der Abschlag dagegen dient vor allem als Gegenkraft zur Gravitation, d.h. die Körpermasse des Schmetterlings wird dabei ausgeglichen. Die Autoren betonen, dass damit der schaukelnde und scheinbar wenig effiziente Flug des Kaisermantels in einem neuen Licht erscheint. Im Vergleich zu Modellen mit starren Flügeln bringt die aufgrund der Flexibilität der Flügel erzeugte tassenförmige Struktur einen um 22 % höheren Impuls im Vortrieb und eine um 28 % gesteigerte Effektivität beim Fliegen. Auch diese Autoren bemerken abschließend in ihrer Veröffentlichung, dass diese neu gewonnenen Erkenntnisse zur Verbesserung von Flugrobotern genutzt werden können.
Die hier angeführten Einsichten könnten uns dazu herausfordern, Schmetterlingen nicht nur aufgrund ihrer vielfältigen Farbenpracht oder ihres Nutzens durch ihre Bestäubertätigkeit aufmerksam und genau zu beobachten. Es wird deutlich, wie viel uns bei den bisherigen Beobachtungen verborgen geblieben und deshalb noch zu entdecken ist. Es kann uns aber auch zum Staunen bringen und nachdenklich machen, wenn wir hier – wie auch an vielen anderen Beispielen – erkennen, wieviel Detailwissen und vielfältige Prinzipien in Lebewesen verwirklicht sind. Wer dies wahrnimmt, kann auch bedenken, was ein alter Prophet in Israel sagt: Er ist es, der die Erde gemacht hat durch seine Kraft, der den Erdkreis gegründet durch seine Weisheit und den Himmel ausgespannt durch seine Einsicht (Jer. 10,12).
Anmerkung
1 Ein Polysaccharid ähnlich der Cellulose; der Monomerbaustein besteht anstelle von Glucose aus N-Acetylglukosamin.
Literatur
Fuchiwaki M, Kuroki T, Tanaka K & Tababa T (2013) Dynamic behavior of the vortex ring formed on a butterfly wing. Exp. Fluids 54, 1450–1461.
Johansson LC & Henningsson P (2021) Butterflies fly using efficient propulsive clap mechanism owing to flexible wings. J. R. Soc. Interface 18: 20200854; doi.org/10.1098/rsif.2020.0854
Lilienthal O (1889) Der Vogelflug als Grundlage der Fliegekunst. Ein Beitrag zur Systematik der Flugtechnik. Berlin. https://www.deutschestextarchiv.de/book/show/lilienthal_vogelflug_1889
Autor dieser News: Harald Binder
© 2021, http://www.genesisnet.info/schoepfung_evolution/n286.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
25.11.20 Die Optimalität des genetischen Codes – ein klarer Beleg für Intelligentes Design
Der genetische Code wurde in den 1960er-Jahren entschlüsselt und ist der Code, durch den die Information zur Herstellung von Proteinen in der DNA übermittelt wird. Eine bemerkenswerte Tatsache ist, dass alle Organismen praktisch denselben Code besitzen, während theoretisch Milliarden solcher Codes auftreten könnten. Noch immer gilt dies als Beweis für die gemeinsame Abstammung aller Organismen. Neue Entdeckungen zeigen jedoch, dass dieser Code optimal ist, um die Wirkung von Mutationen abzupuffern. Der genetische Code ist daher „antievolutiv“! Wer hätte gedacht, dass der universelle genetische Code heute einer der besten Beweise für intelligentes Design ist?
Die Wissenschaft hat im Universum außer mit Energie und Materie ganz offensichtlich auch mit Information zu tun. In Atomen und Molekülen erkennen wir zum einen insofern Information, als sie sich in den Naturgesetzen, im unsichtbaren atomaren und molekularen Verbindungsverhalten der Atome und Moleküle manifestiert. In der Biologie ist sie zum anderen in Entwicklungsabläufen erkennbar, nach denen sich einzelne Keimzellen gesetzmäßig zu Organismen entwickeln. Die immer wiederkehrende Choreographie der frühen Entwicklung des Fadenwurms, die die Entfaltung von genau 671 Zellen umfasst, und der Zelltod von 111 Zellen (oder 113, je nach dem Geschlecht des Fadenwurms) ist für die in biologischen Systemen vorhandene Information beispielhaft. Auch der vorprogrammierte Tod der nicht mehr benötigten Zellen wird durch Information bestimmt. Die Organisation und der Selbstaufbau, zwei typische Merkmale des sich entfaltenden Lebens, sind durch Information gesteuerte Phänomene.
Information wurde von den materialistischen Philosophen des 19. Jahrhunderts, den Naturalisten, nicht beachtet, da das Informationskonzept, wie wir es heutzutage kennen, noch nicht entwickelt war. Ein Paradebeispiel ist das Erbmolekül DNA. Die Abfolge der Bausteine der DNA ist aperiodisch, d. h. sie existiert als eine schier unendliche Reihe von augenscheinlich ungeordneten Abfolgen chemischer Verbindungen (den Nukleotiden Adenin, Thymin, Cytosin und Guanin, die mit den Symbolen A, C, T und G abgekürzt werden). Die Nicht-Periodizität ermöglicht es, ein Informationsträger zu sein, wie das auch bei unserer nicht-periodischen Sprache der Fall ist. Eine DNA-Sequenz ist dementsprechend eine Symbolisierung funktionaler Merkmale, die wir in Lebewesen vorfinden.
Die Information einer DNA-Sequenz kann ihre Funktion allerdings nur dann entfalten, wenn sie von Decodierungs-Maschinen abgelesen wird. Diese Maschinen sind ebenso wie die digitalisierte Information in der DNA der Zellen vorhanden. Die codierte Information wird benötigt, damit sich eine einzige Zelle zuerst zu einem Embryo, weiter zu einem Fetus, zu einem Baby und schließlich zu einem erwachsenen Individuum entwickelt.
Das Genom des Menschen (also sein komplettes Erbgut) und weitere epigenetische Informationssysteme enthalten zum Beispiel die Information für den Aufbau einer Hand mit fünf Fingern und eines Fußes mit fünf Zehen, und für die Stellen, wo die Sehnen befestigt werden müssen. Ebenso gibt es Information, wie durch den Stoffwechsel in den Zellen Aminosäuren auf- und abzubauen sind und wie daraus Proteine hergestellt werden können. Die Information im Genom bestimmt, wie mehrere hundert verschiedene Zelltypen gebildet werden, und auch die Moleküle, die dafür sorgen, dass die verschiedenen Zelltypen als Ganzes funktionieren. Information wird auch für den Bau und die Entwicklung von Magen, Leber, Nieren, Lungen und Herz und alle anderen Organe benötigt. Die Form bzw. Ausprägung von Organen wie Augen oder Nase ist in der genetischen Information festgelegt. Alle Eigenschaften eines Organismus vererben sich als Informationspakete. Das Geheimnis des Lebens ist, dass Lebewesen auf Information beruhen.
Der genetische Code. Die Tatsache, dass es – mit wenigen Ausnahmen – nur einen einzigen genetischen Code bei allen Organismen gibt, wurde im vergangenen Jahrhundert als ein überzeugender Beweis für den Grundsatz der universellen Abstammung gefeiert. Durch den Code ist die Zuordnung der Abfolgen von Nukleotiden (A, C, G, T) zu entsprechenden Abfolgen von Aminosäuren in Proteinen festgelegt. Jeweils drei Nukleotide stehen für eine bestimmte Aminosäure (und einige stehen für Start und Stopp), und diese Entsprechung ist bei fast allen Organismen gleich. Doch diese Einheitlichkeit des Codes hat noch eine andere Seite und diese beinhaltet einen guten Grund, zu einer anderen Schlussfolgerung als „gemeinsame Abstammung“ zu gelangen. Zur Erklärung müssen wir etwas ausholen.
Mit den vier digitalisierten Symbolen (A, T, C und G) können durch Dreiergruppen (s. o.) insgesamt 64 verschiedene Kombinationen zusammengestellt werden. Diese sogenannten Tripletts – oder Codons – verschlüsseln also die Bausteine der Proteine, die Aminosäuren, wovon es es nur 20 unterschiedliche in den Lebewesen gibt (auch hier mit wenigen Ausnahmen). Es gibt also viel mehr Codons (64) als zu codierende Proteinbausteine (20 Aminosäuren). Zusammen mit dem Codon, das als Stoppsignal fungiert und anzeigt, wo das Protein endet, benötigen die Lebewesen nicht mehr als 21 Codons, um alle beliebigen Proteine codieren zu können. Wozu dienen die restlichen 43 Kombinationen?
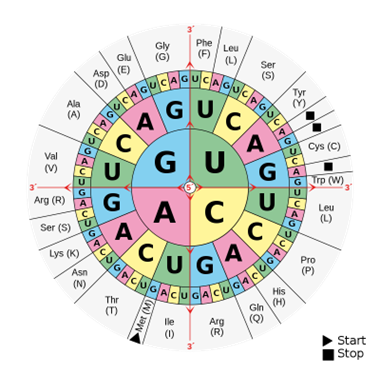
Abb. 1: Die Codesonne zeigt, welche Basensequenz welche Aminosäure ergibt. (Wikimedia: Mouagip, Gemeinfrei)
Genau genommen gibt es drei unterschiedliche Stoppsignale, die übrigen Codons erweisen sich als alternative Tripletts, die für dieselben 20 Aminosäuren codieren. Es heißt, der Proteincode ist ein degenerierter Code. Damit ist nicht gemeint, dass er irgendwie defekt sei, sondern dass eine bestimmte semantische Einheit (eine bestimmte Aminosäure) durch mehrere unterschiedliche syntaktische Abfolgen (die Codons) codiert wird. Den meisten Aminosäuren entsprechen mehrere Codons. Die Aminosäuren Serin, Arginin und Leucin werden beispielsweise von jeweils sechs unterschiedlichen Codons codiert. Vier verschiedene Codons stehen für jeweils fünf andere Aminosäuren, nämlich Alanin, Glycin, Prolin, Threonin und Valin. Die übrigen Aminosäuren werden durch einen, zwei oder drei Codons codiert; Aminosäuren, die von fünf Codons dargestellt werden, kommen nicht vor. Wozu gibt es diese seltsame Verteilung? Wäre ein Code, in dem alle 20 Aminosäuren dreifach codiert wurden, zuzüglich der vier Stoppsignale, nicht ebenso funktionsfähig? Diese seltsame, ungleiche Verknüpfung von Codons und Aminosäuren war lange Zeit rätselhaft. Im 21. Jahrhundert ist dieses Rätsel dank unseres detaillierten Wissens bezüglich der molekularen Grundlagen des Lebens gelöst worden: Der Code ist in verschiedener Hinsicht optimal, insbesondere in Bezug auf Fehlertoleranz (Freeland et al. 2000).
Mit 64 verschiedenen Kombinationen von drei Symbolen können wie erwähnt 20 Aminosäuren auf viele Weisen zugeordnet werden. Theoretisch sind Milliarden solcher Zuordnungen vorstellbar, welche als alternative genetische Codes fungieren könnten. Mittlerweile ist man in der Lage, diese alternativen Möglichkeiten zu erproben und zu bewerten. Dabei hat sich der tatsächlich vorhandene universelle Code als optimal herausgestellt, um zufällige genetische Veränderungen (Mutationen) so ausgleichen zu können, dass die Proteine sich in ihrer Funktion möglichst nicht ändern (Freeland et al. 2000). Der genetische Code ist gerade so mit der Synthese von Proteinen verbunden, dass Mutationen im Erbgut das Endprodukt (Protein) möglichst wenig beeinträchtigen können. Anders gesagt, der Code ist daraufhin eingerichtet, Veränderungen optimal vorzubeugen und den Status Quo zu bewahren. Der Code ist optimal in allen Lebewesen und es gibt keinen Grund für die Annahme, dass es jemals anders war. Das ist bemerkenswert.
Wenn man ein Team von Wissenschaftlern herausfordern wurde, den für das Leben passendsten genetischen Code zu entwerfen, und vorausgesetzt, sie wüssten alles über die molekulare Biologie, die Biochemie und die Biophysik (was sie nicht tun), dann würden sie genau diesen Code (oder einen sehr ähnlichen, gleichwertigen Code) entwerfen, den alle Lebewesen aufweisen. Ist das nicht Grund zum Nachdenken? Zeugt diese Feinabstimmung des universellen Codes nicht eher von intelligentem Design als von universeller Abstammung?
2013 beschrieb ein russisches Team aus Bio-Informatikern, dass der biologische Code, wie er in allen Organismen vorkommt, alle Indizien eines intelligenten Ursprungs beinhaltet (shCherbak & Makukov 2013). Sie äußerten, dass das eigentliche Szenario für die Herkunft des irdischen Lebens bei Weitem noch nicht geklärt sei, und dass die Idee, dass das Leben planvoll von einer Intelligenz „ausgesät“ wurde, nicht a priori ausgeschlossen werden dürfe. Ein statistisch starkes intelligentes Signal im genetischen Code ist eine prüfbare Folge eines solchen Szenarios. Laut diesen Forschern weist der Aminosäure-Code eine ausgefeilte Feinabstimmung auf, die den Kriterien entspricht, die als Informationssignal zu betrachten sind. Genau und systematisch erscheinen die zugrundeliegenden Design-Muster als Produkt der Präzisionslogik und Feinabstimmung. Die Bio-Informatiker sahen sich gezwungen, die Nullhypothese, wonach der Code ein Produkt des Zufalls sei, zu verwerfen. Die Idee, dass der genetische Code auf natürliche Weise entstanden ist, wurde entschieden in Frage gestellt. Die Design-Muster des Codes waren so stark, dass die Code-Zuordnung selbst eindeutig aus ihrer algebraischen Darstellung abgeleitet werden könnte. Das dahinterliegende Signal zeigt leicht erkennbare Merkmale der Künstlichkeit, darunter das Symbol Null, die privilegierte Dezimalsyntax und semantische Symmetrien. Von dem intelligenten Design des Codes beeindruckt nannten sie ihren Artikel The “Wow! signal” of the terrestrial genetic code. Als „Wow!-Signal“ bezeichnen Astronomen, die den Kosmos mit Radiotelekopen absuchen, ein Signal, das alle Kennzeichen eines interstellaren Kommunikationsversuchs von intelligentem außeridischem Leben aufweist. Anders gesagt, das Signal kann nur von intelligenten Urhebern herrühren. Genau ein solches Signal fanden die russischen Forscher in der DNA der lebenden Zellen!
Meinte Nobelpreisträger Francis Crick 1968 noch, der Proteincode sei ein eingefrorener Zufall, stellt die moderne Forschung ihn als einen außerordentlich zuverlässigen Informationsspeicher mit eindeutiger intelligenten Signatur dar. Diese neuen wissenschftlichen Erkenntnisse erfordern eine Umkehr des Denkens: Die Frage nach dem Ursprung des Lebens kann besser durch die Annahme eines Schöpfers beantwortet werden.
Literatur
Freeland SJ, Knight RD, Landweber LF & Hurst LD (2000) Early Fixation of an Optimal Genetic Code. Molecular Biology and Evolution 17, 511–518. https://academic.oup.com/mbe/article/17/4/511/1127636
shCherbak V & Makukov MA (2013) The “Wow! Signal” of the terrestrial genetic code. Icarus 224, 228–242. https://www.sciencedirect.com/science/article/abs/pii/S0019103513000791
Autor dieser News: Peter Borger
© 2020, http://www.genesisnet.info/schoepfung_evolution/n282.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.03.20 Rätselhafte Verteilung der Wurzelknöllchensymbiose
Die Wurzelknöllchensymbiose bei Leguminosen (z. B. Bohnen oder Erbsen) ist eine fein ausbalancierte Symbiose zwischen Pflanze und Bakterien zur Fixierung von Luftstickstoff und von erheblicher wirtschaftlicher Bedeutung. Überraschenderweise muss in evolutionstheoretischen Modellierungen eine mehrfache Entstehung und ein mehrfacher Verlust angekommen werden.
Ein wichtiger „Rohstoff“ für das Pflanzenwachstum ist Stickstoff, er wird z. B. für die Synthese der Proteine und Nukleinsäuren benötigt. Elementarer, zweiatomiger Stickstoff (N2) ist mit 78 % Anteil zwar reichlich in der Luft vorhanden, kann aber von den Pflanzen nicht direkt genutzt werden, sondern nur in Form von Ammonium (NH4+) oder Nitrat (NO3-). Nur einige Bodenbakterien sind in der Lage, Luftstickstoff durch Reduktion in eine für Pflanzen verfügbare Form zu überführen; das von ihnen produzierte Nitrat kann von den Pflanzen über die Wurzeln aufgenommen werden. Diesen Vorgang nennt man Stickstoffbindung (Nitrifizierung).
Es gibt jedoch vier Pflanzenordnungen, in denen einige Arten in bestimmten Bereichen der Wurzeln eine Symbiose mit stickstoffbindenden Bakterien der Gattungen Rhizobium oder Frankia eingehen können. Diese Symbiose findet in sogenannten Wurzelknöllchen statt. Die Bakterien in den Wurzelknöllchen sind als Bakterioide ausgebildet, das heißt, sie besitzen keine Zellwände. Die Bakterioide werden von den infizierten Pflanzenzellen in Membranen eingehüllt und bilden dadurch gleichsam Zellorganellen; sie besitzen die für die Stickstoffbindung erforderlichen Enzyme. Das wichtigste darunter ist die Nitrogenase. Dieses Enzym wird allerdings bei Anwesenheit einer bereits geringen Menge an Sauerstoff geschädigt, andererseits kommen die Bakterioiden nicht ohne Sauerstoff aus. Die Sauerstoffkonzentration in den Wurzelknöllchen muss daher genau ausbalanciert sein. Dafür sorgt die Pflanze, indem sie ein eisenhaltiges Protein, das Leghämoglobin, in den Knöllchen bildet, das überschüssigen Sauerstoff bindet, dessen Niveau konstant hält und somit eine Sauerstoffpuffer-Funktion erfüllt. Außerdem sorgt es für einen optimalen Sauerstofffluss zu den Bakterioiden. Das stickstofffixierende System benötigt also sehr anspruchsvolle Rahmenbedingungen. Das Leghämoglobin ähnelt übrigens in seinem Aufbau stark dem Hämoglobin des Blutes der Wirbeltiere, dessen Funktion ebenfalls darin besteht, Sauerstoff zu binden.
Beide Partner profitieren von dieser Symbiose, denn während die Pflanze mit Stickstoff versorgt wird, erhalten die Bakterien Kohlenhydrate und andere organische Verbindungen von der Pflanze. Die Symbiose in den Wurzelknöllchen ist von großer wirtschaftlicher Bedeutung, da überschüssige Ammonium-Ionen ausgeschieden werden und dadurch der Stickstoffgehalt des Bodens zunimmt, was den Ertrag verbessert. Daher werden wurzelknöllchenbildende Leguminosen angebaut und untergepflügt, um dadurch die Stickstoffdüngung zu verbessern.
Die stickstofffixierenden Wurzelknöllchen (nitrogen fixing root nodule, NFN) sind bei Arten von vier Pflanzenordnungen nachgewiesen, die als NFN-Klade zusammengefasst werden (Fabales, Fagales, Cucurbitales und Rosales), darunter sind viele landwirtschaftlich wichtige Arten wie Bohnen, Erbsen und Soja. Die meisten Arten dieser vier Ordnungen besitzen allerdings keine NFN; die Arten mit NFN sind vielmehr sporadisch verteilt und auch innerhalb der vier NFN-Ordnungen meist nicht näher verwandt. Nur bei zehn der 28 Familien des NFN-Klades kommen Arten mit NFN vor, und bei neun von diesen Familien haben die meisten Arten wiederum kein NFN (Griesmann 2018, 1f.). Die NFN zeigen dabei in ihrer Ausprägung große Vielfalt. Diese unsystematische Verteilung erfordert in evolutionstheoretischer Deutung eine mindestens vierfach unabhängige Entstehung von NFN. Das an sich ist angesichts der Komplexität und anspruchsvollen Ausbalanciertheit des Systems erstaunlich und eines von mittlerweile ungezählten Beispielen einer Konvergenz von Komplexmerkmalen, wie man sie auf der Basis ungerichteter Evolutionsmechanismen nicht erwarten kann (zur Problematik von Konvergenzen siehe den Grundsatzartikel von Braun 2012). Nagy (2018) bezeichnet diese Konvergenz als „uraltes Mysterium“ („an age-old mystery“).
Überraschende Ergebnisse einer neuen Studie. Eine neuere genetische Studie offenbarte nun dazu noch weitere Überraschungen. Griesmann et al. (2018) sequenzierten die Genome von zehn Pflanzenarten mit unterschiedlichen Knöllchentypen und bakteriellen Symbionten und bezogen sie anschließend in einen genomweiten Vergleich mit insgesamt 37 Pflanzenspezies ein.
Es stellte sich heraus, dass den Symbiosen der nicht näher verwandten Arten ähnliche Gen-Garnituren zugrunde liegen. Neben zwei Symbiose-Schlüsselgenen (NIN und RPG) werden weitere 290 gemeinsame Gene in den Knöllchen von Medicago truncatula (Ordnung Fabales) und Parasponia andersonii (Ordnung Rosales) hochreguliert, obwohl sie die Fähigkeit zur Knöllchenbildung unabhängig erworben hätten und nach evolutionstheoretischen Modellierungen seit über 100 Millionen Jahren getrennte Wege gehen sollen (Nagy 2018). „Wie kann dieses Ausmaß genetischer Ähnlichkeit mit der phylogenetischen Unregelmäßigkeit der Knöllchenbildung in Einklang gebracht werden?“ fragt Nagy verständlicherweise. Neue Modelle zur Erklärung konvergenter Evolution seien erforderlich, so Nagy (2018). Es müsse ein Szenario zwischen den beiden Extremen „16 malige unabhängige Entstehung“ und „einmalige Entstehung mit vielfachem Verlust geben. Plausibel ist keines dieser Szenarien, sie sind nur logische Schlussfolgerungen bei Voraussetzung von Evolution.

Abb. 1: Medicago truncatula aus der Gattung Schneckenklee geht eine Symbiose mit Knöllchenbakterien ein. (Wikimedia: Ninjatacoshell, CC BY-SA 3.0)
Die Forscher vermuten für die mehrfache Entstehung des NFN-Komplexes eine Prädisposition bei dem gemeinsamen Vorfahren des NFN-Klades, die die weitere unabhängige Evolution der NFN-Symbiose ermöglicht habe. Diese gemeinsame Prädisposition ist aber eine bloße ad-hoc-Mutmaßung, und die weiteren anzunehmenden unabhängigen Schritte zu den NFN sind komplex. Dasselbe gilt für die Annahme einer Co-option (Einbau, Übernahme) passender vorhandener Gene (Griesmann et al. 2019, 5). Für komplexere Konstellationen ist ein solcher Vorgang experimentell nicht nachgewiesen und theoretisch unplausibel außer beim Vorliegen einer programmierten Situation.
Rätselhafter Verlust. Die Daten über die Verteilung von Arten mit NFN legen überraschenderweise nahe, dass die Fähigkeit zur Stickstofffixierung öfter verlorengegangen sei. Dass es zu mehrfachen Verlusten der Symbiose gekommen sein soll, ist angesichts des großen Nutzens dieser Fähigkeit überraschend und recht unverständlich – auch Kulturpflanzen wie Erdbeeren, Brombeeren oder Äpfel sind davon betroffen. Die Forscher stellten fest, dass das für die Wurzelknöllchensymbiose essenzielle NIN-Gen in verschiedenen Linien mehrfach unabhängig voneinander mutiert ist. Die Gründe für den Verlust sind unklar und es können nur Mutmaßungen über mögliche Selektionsdrücke zuungunsten der Symbiose getroffen werden. Diskutiert werden Befall mit parasitischen Bakterien, ein dauerndes Überangebot an Stickstoff oder eine begrenzte Verfügbarkeit von Wasser oder Phosphat für das Pflanzenwachstum.
Fazit. Insgesamt erscheint die Verteilung der Pflanzen mit dem NHN-Komplex für jedes Ursprungsmodell rätselhaft. Die anscheinend unsystematische Verteilung dieses Komplexmerkmals im System der Blütenpflanzen ist für alle Ursprungsmodelle eine Herausforderung. Angesichts der komplexen Abhängigkeiten von Wirt und Symbiont ist eine Entstehung durch ungerichtete evolutionäre Prozesse unplausibel. Das gilt erst recht bei mehrmaliger unabhängiger Entstehung.
Literatur
Braun HB (2012) Warten auf einen neuen Einstein.
Griesmann M, Chang Y, et al. (2018) Phylogenomics reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 361, eaat1743; doi:10.1126/science.aat1743.
Nagy LG (2018) Many roads to convergence. Science 361, 125-126.
Autor dieser News: Reinhard Junker
© 2020, http://www.genesisnet.info/schoepfung_evolution/n275.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.11.19 Superschnelle Wüstenameise
Die Wüstenameise Cataglyphis bombycina schafft in einer Sekunde eine Strecke, die dem 100-fachen ihrer Körperlänge entspricht. Dies gelingt mit einer sehr hohen Schrittfrequenz und einer extrem schnellen Bewegung beim Schwingen der Beine. Die Beine berühren den Untergrund nur sehr kurze Zeit, so dass die Ameise beim Laufen gleichsam auch „fliegt“.
Ameisen genießen große Aufmerksamkeit von Biowissenschaftlern, z. B. werden auffällige und außergewöhnliche Verhaltensweisen an diesen sozial lebenden Insekten untersucht. Ameisen besiedeln die unterschiedlichsten Lebensräume und weisen eine enorme Vielfalt an dafür geeigneten Ausstattungen auf. Wüstenameisen leben in einem extremen Lebensraum, z. B. in Bezug auf Temperatur, Wasser, Nahrung etc. So lebt Cataglyphis bombycina in den Sanddünen der Nordafrikanischen Sahara und zeigt nach Untersuchungen von Pfeffer et al. (2019) mit 855 mm s-1 die höchste Geschwindigkeit, die bisher für solche Tiere gemessen wurde. Die Ameise legt bei diesem Tempo in einer Sekunde eine Strecke zurück, die dem 100-fachen ihrer Körperlänge entspricht. Übertragen auf einen Menschen mit einer Körpergröße von 1,80 m würde das einer Geschwindigkeit von 180 m s-1 entsprechen. Usain Bolt, der 2009 in Berlin den aktuellen Weltrekord über 100 m in 9,58 s aufstellte, könnte da nur staunen; er musste aber auch eine sehr viel größere Masse beschleunigen.

Abb. 1: Silberameisen (Cataglyphis bombycina) beim Fressen einer Kamelzecke. (Wikimedia: Bjørn Christian Tørrissen, http://bjornfree.com/galleries.html, CC BY-SA 3.0)
Diese extrem hohe Geschwindigkeit erreichte C. bombycina mit kürzeren Beinen als die bisher geführte Rekordhalterin C. fortis. Die Analyse von Videos aus Hochgeschwindigkeitsaufnahmen lieferte Hinweise auf eine sehr hohe Schrittfrequenz (< 40 Hz) und extrem schnelle Bewegung beim Schwingen der Beine (bis zu 1400 mm s-1). Bei diesen schnellen Bewegungen sind die Beine sehr gut synchronisiert, und zwar so, dass immer drei Beine aus Vorder- und Hinterbein einer Seite mit dem mittleren Bein der anderen Seite einen Dreibeinstand zeigen. Die Beine berühren den Untergrund nur sehr kurze Zeit (7 ms) und die Ameise befindet sich vor dem folgenden Dreibeinstand ohne Bodenberührung in der Luft, sie „fliegt“ quasi. Diese „Flugphasen“ treten bereits auf, bevor die Höchstgeschwindigkeit erreicht wird.
Für die Wüstenameise, die tagaktiv ist und in der Mittagshitze bei wenig Verkehr im Wüstensand auf Nahrungssuche geht, sind diese Fähigkeiten sehr hilfreich: so ist sie nicht unnötig lange der lebensfeindlichen Hitze ausgesetzt. Im Übrigen scheint sie die hohen Temperaturen für ihre Höchstgeschwindigkeit zu benötigen, im Labor bei kühlen 10 °C erreichten die Ameisen nur Geschwindigkeiten von 57 mm s-1.
Die beiden von Pfeffer et al. verglichenen Wüstenameisen C. bombycina und C. fortis erreichen die auffällig hohen Geschwindigkeiten bei ihrer Fortbewegung auf unterschiedliche Weise (Beinlänge, Schrittfrequenz und -länge) und sind doch beide für ihren Lebensraum gut ausgerüstet.
Literatur
Pfeffer SE, Wahl VL, Wittlinger M & Wolf H (2019) High-speed locomotion in the Saharan silver ant, Cataglyphis bombycina. J. Exp. Biol. 222, doi:10.1242/jeb.198705
Autor dieser News: Harald Binder
© 2019, http://www.genesisnet.info/schoepfung_evolution/n272.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
04.05.18 Fledermäuse mit „Long Life“-Garantie
Kann man den Alterungsprozess verzögern? Man kann, wenn „man“ eine Fledermaus der Art Myotis myotis (Großes Mausohr) ist. Sie kann ungefähr zehnmal so alt werden im Vergleich zu Säugetieren mit vergleichbarem Gewicht. Wissenschaftler haben einige wahrscheinliche Ursachen dafür herausgefunden. Liegt hier ein Ansatz für eine Erklärung der hohen Alter der biblischen Stammväter?
Eines der rätselhaften Phänomene des Lebens ist das Altern. Die Hoffnung ist, dass durch ein Verständnis der Alterungsprozesse auch Einfluss auf deren Geschwindigkeit genommen werden könnte. Möglichst lange jung zu bleiben, ist ein uralter Traum der Menschheit. Bibelleser kennen die hohen „biblischen“ Alter der Urväter mit dem Altersrekord von 969 Jahren des auch sprichwörtlich genutzten Methusalem. Könnten solche hohen Alter realistisch sein oder sind sie eine biologische Unmöglichkeit?
Von überraschender Seite wurden zu dieser Frage kürzlich hochinteressante Befunde publiziert. Eine Forschergruppe (Foley et al. 2018) berichtete über Untersuchungen des Alterns bei verschiedenen Arten von Fledermäusen und fand dabei heraus, dass der Vorgang des Alterns beim Großen Mausohr (Myotis myotis) sehr viel langsamer als bei anderen Fledermausarten verläuft. Bekanntlich gibt es bei Säugetieren einen ungefähren Zusammenhang zwischen Körpergewicht und Lebenserwartung. Je geringer das Körpergewicht, desto geringer das Lebensalter. So haben Mäuse eine Lebenserwartung von wenigen Jahren, Hunde werden bis zu ca. 15 Jahre alt, große Säugetiere ca. 50 Jahre. Der Mensch fällt somit deutlich aus dem Rahmen. Aber im Vergleich zum Körpergewicht ist das noch nichts gegen das Große Mausohr. Obwohl es nur wenige Gramm schwer wird und entsprechend nur wenige Lebensjahre zu erwarten hätte, erreichen die Tiere oft ein Lebensalter von 35 Jahren, ungefähr zehnmal so viel wie aufgrund ihres Gewichts zu erwarten wäre.
Was ermöglicht diesen enormen Ausreißer? Schon lange wird ein Zusammenhang zwischen dem Altern und der Länge der Endstücke der Chromosomen, der Telomere, vermutet. Bei jeder Zellteilung werden die Telomere verkürzt, bis schließlich keine Zellteilungen mehr möglich sind. Allerdings verzögert die Tätigkeit des Enzyms Telomerase die Verkürzung, indem es die Telomere in beschränktem Maße wiederherstellen kann, was jedoch mit zunehmendem Alter nachlässt. Entsprechend können Mutationen der Telomerase zu vorzeitigem Altern führen. Beim berühmten Klon-Schaf Dolly waren die Telomere schon bei der Geburt verkürzt. Die geringe Lebensspanne und der früh einsetzende Altersprozess von Dolly dürften damit zusammenhängen und dieser Befund wird auch als Stütze für die Telomer-Hypothese des Alterns angesehen.
Die Verhältnisse bei den Fledermäusen erwiesen sich allerdings als komplizierter. Zwar konnte ein Zusammenhang mit dem Ausbleiben der Telomerverkürzung bestätigt werden, doch die Ursachen dafür sind vielfältig und bestehen nicht nur in der Tätigkeit der Telomerase. Vielmehr erwiesen sich 21 andere Gene als bedeutsam, die an der DNA-Reparatur und an der Wartung der Telomere beteiligt sind. Diese Gene sind spezifisch bei Myotis myotis aktiv. Die Aktivität zweier Gene, ATM und SETX, ist besonders wichtig. Offenbar spielen also neben Telomerschutzprogrammen auch besondere Vorgänge in der DNA-Reparatur bei der erheblichen Verzögerung der Alterung eine Rolle.
Weshalb gerade das Große Mausohr der normalen Alterung entkommt, ist nicht bekannt. Jedenfalls zeigt dieses Beispiel, dass es bezüglich des Alterns sehr weit reichende Ausreißer geben kann. Vor diesem Hintergrund scheint auch ein sehr viel größeres individuelles Alter von Menschen nicht im Widerspruch zu biologischen Sachverhalten zu stehen.
Literatur
Foley NM, Hughes GM et al. (2018) Growing old, yet staying young: The role of telomeres in bats’ exceptional longevity. Sci. Adv. 2018;4: eaao0926
Autor dieser News: Reinhard Junker
© 2018, http://www.genesisnet.info/schoepfung_evolution/n259.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.04.17 „Wunderwerk Feder“
Neue Untersuchungen eröffnen neue Einblicke in den komplizierten Feinbau von Vogelfedern. Eine ausgeklügelte Anordnung von Keratinfasern ermöglicht die besonderen Eigenschaften der Federn – die Kombination von Leichtheit, Stabilität, Flexibilität und Robustheit. In den Beschreibungen wird die Genialität des Federfeinbaus hervorgehoben. Für die ebenfalls geäußerte Auffassung, es handle sich um eine Perfektionierung im Laufe einer langen Evolution, werden dagegen keinerlei Gründe genannt.
„Wunderwerk Feder“ – diese Überschrift stellt Verena Dietrich-Bischoff über eine Zusammenfassung neuer Studien zum Feinbau von Vogelfedern. Im Text ist dann von „Wunderwerken der Evolution“ die Rede; wenn man sich aber die erforschten Details vor Augen hält, kommen Zweifel, ob die genialen Strukturen ein Evolutionsprodukt sein können.
Nimmt man eine Vogelfeder in die Hand, überrascht immer wieder, wie leicht dieses Gebilde ist. Es heißt nicht umsonst „federleicht“. Gleichzeitig sind Vogelfedern ausgesprochen robust und dennoch biegsam und kombinieren damit sehr unterschiedliche Eigenschaften. Diese Kombination stellt besondere Anforderungen an den Feinbau der Federn. Es wird geeignetes Baumaterial benötigt – lange Proteinfasern aus beta-Keratin –, außerdem gibt es anspruchsvolle Anforderungen an die Art und Weise, wie das Material in Federschaft, -ästen und -strahlen „verbaut“ wird, und schließlich sind auch die Anforderungen an die Struktur der reißverschlussartig ineinandergreifenden Federstrahlen (Bogen- und Hakenstrahlen) alles andere als trivial. Denn bei aller Robustheit müssen die Federn auch kontrolliert nachgeben können, wenn starke Kräfte auf sie wirken, damit irreparable Risse vermieden werden; dafür ist der „Reißverschluss“ sehr praktisch. Materialeigenschaften, Feinbau und Federstrukturen sind also genau aufeinander abgestimmt. Es ist leicht einzusehen, dass diese besonderen Eigenschaften der Federn zum einen komplett vorhanden sein müssen, damit Federn flugtauglich sind, und dass sie zum anderen nicht oder nur in sehr viel einfacherer Ausprägung benötigt würden, wenn Federn nur irgendwelche anderen Funktionen wie Wärme- und Kälteschutz, Signalgebung oder Schutz vor Verletzungen erfüllen müssten, nicht aber zum Fliegen gebraucht würden.
Tatsächlich ist der Feinbau von Schaft und Federästen so kompliziert, dass bis vor kurzem viele Details nicht bekannt waren. Die Fasern sind nämlich mit einer Matrix zementartig verbunden, was die Aufklärung ihrer Anordnung erschwert, aber zugleich zu ihrer Stabilität beiträgt (vgl. Lingham-Soliar 2017, 1). Erst in jüngerer Zeit wurden durch neue Untersuchungstechniken hier größere Fortschritte erzielt. Durch Computertomographie (Laurent et al. 2014) und mithilfe federzersetzender Pilze (Lingham-Soliar et al. 2010), durch die die Keratinfasern und Faserbündel freigelegt werden können, wurden ganz neue Einblicke in die Anordnung der Fasern und damit in den Feinbau gewonnen (Lingham-Soliar 2014; 2017).
Dabei hat sich Folgendes gezeigt: Die beta-Keratin-Fasern bestehen aus feineren Fäserchen und sind selbst in Bündeln angeordnet. Diese können verschiedene Schichten bilden, in denen die Fasern unterschiedlich orientiert sein können. Dietrich-Bischoff (2014) fasst die weiteren Befunde wie folgt zusammen: „Nun sind zwei verschiedene Fasersysteme mit unterschiedlicher Orientierung nachgewiesen worden, die unterschiedliche mechanische Eigenschaften verleihen. Im größten Teil des Federschafts laufen die Keratinfasern in Längsrichtung, wodurch die Feder verdreht werden kann, ohne zu brechen. In den Seitenwänden des Schafts hingegen liegen die Fasern über Kreuz, was der Feder größere Steifheit verleiht. … Die Keratinfasern weisen zudem in gewissen Abständen Knötchen auf, die zu denen benachbarter Fasern versetzt angeordnet sind. Diese Anordnung trägt dazu bei, dass sich ein Riss im Federschaft schlecht ausbreiten kann. Zudem konnte gezeigt werden, dass Federschaft und -äste keineswegs hohl sind, sondern ein ‚schaumähnliches‘ Inneres aufweisen – ein poröses Netzwerk von Fasern, die mit einem Polymer beschichtet sind, das Gase bindet. So steht die Feder unter schwachem Druck, was dazu beiträgt, dass sie weniger leicht einknickt. Der Schaum wirkt wie ein Energieabsorber und sorgt vermutlich dafür, dass die Feder nach Verbiegen oder Verdrehen wieder in ihre ursprüngliche Position zurückspringt.“
In einer neueren Arbeit legt Lingham-Soliar (2017) weitere interessante Befunde zur Anordnung der Keratinfasern in den sich verjüngenden Federschäften vor. Er konnte zeigen, wie die Verjüngung erfolgt, ohne dass die Faserverläufe in den Wänden (Cortex) des Schafts auslaufen. Die Keratinfasern biegen vielmehr links und rechts in die Wände der Federäste ab, wodurch die Äste tief im Schaft gleichsam „verwurzelt“ sind. Das ermöglicht Reißfestigkeit und verringert gleichzeitig die Gefahr von Brüchen oder Knicken im Schaft; die Kräfte, die beim Flug auf die Federn wirken, werden so auf die Federn gleichmäßig verteilt. Ein Auslaufen (bzw. ein Abbruch) der Fasern im Cortex würde – so Lingham-Soliar – aus mechanischen Gründen die Gefahr von Beschädigungen erhöhen. Diese Grundstruktur des Abzweigens in die Federäste erwies sich bei den verschiedenen untersuchten Arten als sehr ähnlich (S. 4). Lingham-Soliar (2017, 7f.) lobt diese Lösung als „biomechanisch genial“ („ingenious“); durch diese neu entdeckte Architektur des Cortex der Schaftes und der Äste, wo Hunderte von Fasern in die Äste abzweigen statt beendet zu werden, würden auf einen Schlag viele Sollbruchstellen vermieden.
Kommentar
Diese Details machen deutlich, wie fein abgestimmt verschiedene Aspekte der Federfeinstruktur sein müssen, damit die Feder die für ihre Funktion erforderlichen Eigenschaften besitzt. Angesichts dieser Tatsache überrascht es, wenn Verena Dietrich-Bischoff (2014) abschließend zu den damals bereits bekannten Details schreibt, dass die vorgestellten Befunde zur Feinstruktur von Federn möglicherweise dazu beitragen würden, ein evolutionäres Rätsel, die Entstehung des Vogelflugs, zu lösen. Oder dass Lingham-Soliar (2017, 6) meint, es sei klar, dass die außergewöhnliche Mikrostruktur des Wand des Schafts und der Äste über die Millionen Jahre der Vogelevolution perfektioniert worden sei. Jedenfalls zeigen die neuen Befunde, dass das Rätsel größer ist als je zuvor, denn die Federn erweisen sich als komplizierter gebaut als bisher bekannt. Und man kann durchaus nachvollziehen, dass die Details auch allesamt in der verwirklichten Form ausgeprägt sein müssen, damit die Federn ihren Teil zur Ermöglichung des Fluges leisten können. Die dafür erforderlichen höchst anspruchsvollen Materialeigenschaften sind notwendige Voraussetzungen. Es handelt sich auch um weit mehr als um „Perfektionierung“, sondern vielmehr um eine ausgeklügelte, anspruchsvolle Konstruktion.
Zudem ermöglichen auch die allerbesten Federn noch keinen Flug. Es wird auch eine zweckmäßige Verankerung im Körper benötigt, ein Muskelgeflecht an den Federspulen zur Bewegung der Federn, Blutgefäße, Nervenbahnen und Sinnesorgane, die die Positionen der einzelnen Federn registrieren und ans Gehirn melden, und die passenden Reaktionen darauf, die Einbettung der Federn in ein hydraulisches System von Fett- und Bindegewebe und weitere anatomische Details. Die Anforderungen an die Federn und ihre Steuerung sind wiederum nur ein Teil der Voraussetzungen für die Flugfähigkeit, denn es muss auch ein passendes Federkleid ausgebildet sein, weiterhin dessen koordinierte Steuerung; zahlreiche Muskelpartien sind beim Flug beteiligt; es gibt Anforderungen an den Skelettbau; die Federn bedürfen ständiger Pflege usw.
Es ist daher alles andere als klar, dass und wie die Federstrukturen evolutiv entwickelt und „perfektioniert“ wurden. Das Problem besteht neben zahlreichen Details vor allem auch in der Notwendigkeit vielfacher Abstimmung verschiedenster Aspekte. Dietrich-Bischoff (2014) kommentiert einen kurzen Überblick zum groben Aufbau einer Feder wie folgt: „All diese einzelnen Elemente sind hervorragend an ihre jeweilige Funktion angepasst und wirken zusammen, um der Feder die notwendigen Gesamteigenschaften zu verleihen.“ Zukunftsblinde Prozesse wie Mutation und Selektion sind nach allem, was wir wissen, völlig überfordert, solche Abstimmungen über mehrere Ebenen der Organisation zu ermöglichen.
Natur und Technik. Lingham-Soliar (2017, 10) zieht einen Vergleich mit technischen Materialien; diese seien vergleichsweise einfach strukturiert und die technischen Strukturen seien monofunktional. Ganz anders dagegen die biologischen Strukturen, die viele Funktionen zugleich erfüllen und so kompliziert aufgebaut sind, dass es schwierig ist, sie in die einzelnen Komponenten aufzuspalten, um daraus etwas für die technische Anwendung zu lernen. Von einer Nachahmung in der Technik ist man daher weit entfernt. Auch dieser Sachverhalt spricht zusätzlich mehr dafür, dass die Entstehung von Vogelfedern eines enormen geistigen Inputs bedurfte, während Komplexität und Multifunktionalität sicher nicht für eine natürlich-evolutive Entstehung sprechen.
Vergleich mit anderen Strukturen. Interessant ist auch der Befund, dass es Ähnlichkeiten im Aufbau der Federn mit anderen biologischen Strukturen gibt. Dietrich-Bischoff (2014) weist darauf hin, dass auch der Chitinpanzer von Insekten aus mehreren Schichten besteht, in denen die Chitinfasern auf verschiedene Weise orientiert sein können, wodurch der Panzer an manchen Stellen hart, an anderen biegsam wird. Was das schaumartige Innere des Federschafts betrifft, so sind ähnliche Strukturen auch von Pflanzenstängeln und Stacheln von Tieren bekannt. Lingham-Soliar (2017, 8f.) sieht weiterhin Ähnlichkeiten in der Anordnung von Fasern in der Rücken- und Schwanzflosse des Weißen Hais (Carcharodon carcharias) und im Verlauf von Leitungssystem und Fasern bei der Verbindung von Ästen mit Stämmen von Pflanzen, die ähnlichen Kräften ausgesetzt seien wie die Federäste. Auch hier stellt sich die Frage: Sind zukunftsblinde Prozesse in der Lage, unabhängig voneinander in ganz verschiedenen Systemen dieselbe anspruchsvolle Lösung für schwierige Aufgaben zu finden?
Literatur
Dietrich-Bischoff V (2014) Wunderwerk Feder. Die Vogelwarte 52, 59.
Laurent CM, Palmer C, Boardman RP, Dyke G & Cook RB (2014) Nanomechanical properties of bird feather rachises: exploring naturally occurring fibre reinforced laminar composites. J. R. Soc. Interface 11: 20140961; doi:10.1098/rsif.2014.0961
Lingham-Soliar T (2014) Feather structure, biomechanics and biomimetics: the incredible lightness of being. J. Ornithol. doi:10.1007/s10336-013-1038-0.
Lingham-Soliar T (2017) Microstructural tissue-engineering in the rachis and barbs of bird feathers. Sci. Rep. 7:45162; doi:10.1038/srep45162.
Lingham-Soliar T, Bonser RHC & Wesley-Smith J (2010) Selective biodegradation of keratin matrix in feather rachis reveals classic bioengineering. Proc. R. Soc. 277B, 1161-1168.
Autor dieser News: Reinhard Junker
© 2017, http://www.genesisnet.info/schoepfung_evolution/n248.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.12.16 Kurzflügler als „Ameisenkäfer“ – eine erstaunliche Konvergenz
Mindestens 12-mal unabhängig ist eine ausgefeilte Strategie mancher Kurzflügler-Käfer entstanden. Diese Käfer sehen aus wie Ameisen und riechen und verhalten sich wie Ameisen, um unerkannt in Nestern von Wanderameisen auf Beutezug gehen zu können. Da „Design“ als Erklärungsform aufgrund weltanschaulicher Vorentscheidungen ausgeschlossen wird, gerät man bei evolutionären Erklärungsversuchen in Zirkelschlüsse.

Abb. 1: Ameisenkäfer Scydmaenus tarsatus. (Wikimedia: Siga, CC BY-SA 3.0)
Ist es eine Überraschung, dass Merkmale oder Fähigkeiten von Lebewesen mehrfach unabhängig – konvergent – entstehen? Aus einer Designer-Perspektive ist ein zahlreiches Auftreten von Konvergenzen durchaus zu erwarten. Denn ein Designer kann planen, die Zukunft gedanklich vorwegnehmen und entsprechende Schritte einleiten, um ein Ziel zu erreichen. Es ist nicht überraschend, wenn er sich dabei öfter ähnlicher Mittel bedient. Natürliche Selektion kann das alles nicht, denn sie ist genauso zukunftsblind wie die Mutationen, die ihr das Rohmaterial liefern. Daher haben Evolutionstheoretiker, für die das Wechselspiel von Mutation und Selektion der hauptsächliche Motor des Artenwandels ist, allen Grund sich zu wundern, wenn Konvergenzen auftreten, besonders wenn es sich um Komplexmerkmale handelt und wenn sie sich häufen. Und sie wundern sich ja tatsächlich häufig. Denn wenn man zweimal oder gar mehrfach denselben Punkt erreicht, ohne ihn je angesteuert zu haben, stellt sich die Frage, ob es doch eine unerkannte Strategie oder Steuerung gibt. Evolutionstheoretiker verweisen hierzu bei Konvergenzen auf gleiche Selektionsdrücke, Präadaptionen (Voranpassungen) oder auf konstruktive Zwänge. Damit ist gemeint, dass evolutive Veränderungen immer an den bereits vorhandenen Konstruktionen ansetzen müssen und daher nur umbauen können. Die Vorkonstruktionen erlauben „Umbauten“ nur in bestimmte Richtungen – setzen also Konstruktionszwänge.
Alle drei Faktoren – Selektionsdrücke, Präadaptionen und Konstruktionszwänge – können aber nicht als Ursachen für Veränderungen gelten, sondern nur als Begleiterscheinungen. In vielen Fällen ist nicht einmal klar, warum sie als Rahmenbedingungen eine kanalisierende Wirkung haben sollen. Das trifft insbesondere bei Verhaltenskonvergenzen zu. Ein besonders eindrucksvolles Beispiel einer Konvergenz eines ausgefallenen Verhaltens untersuchten Maruyama & Parker (2016) bei parasitisch lebenden kleinen Käfern (vgl. die Zusammenfassung bei Pennisi 2016). Mindestens 12-mal unabhängig sollen Kurzflügler-Käfer aus der Unterfamilie der Aleocharinae die Fähigkeit erworben haben, in Nester von Wanderameisen eindringen und dort auf Beutezug gehen zu können; dabei vergreifen sie sich auch der Brut der Ameisen. Das hört sich auf den ersten Blick vielleicht einfacher an als es ist. Doch um gegen Wanderameisen ankommen zu können, muss man sich einiges einfallen lassen. Nicht umsonst werden diese Ameisen auch als Heeresameisen bezeichnet (engl. „army ants“). Denn sie sind darauf spezialisiert, in ganzen Gruppen auszuschwärmen und auch Eindringlinge zu attackieren.
Um hier überhaupt eine Chance zu haben, in die Nester zu gelangen und dort eine Zeitlang zu verbleiben, müssen ungebetene Besucher sich tarnen können. Den Kurzflüglern gelingt dies dadurch, dass sie den „Käfer-Look“ gegen ein ameisenartiges Aussehen getauscht haben: Sie besitzen eine verengte Taille, verlängerte Beine, Antennen mit ameisenartigen Ellenbogengelenken und anderen Besonderheiten im Körperbau. Aber das genügt nicht: Die flohgroßen Käfer müssen auch durch ihr Verhalten – z. B. einen ameisenartigen Gang – und durch ihren Geruch vortäuschen, dass sie Artgenossen der Ameisen seien. Damit gelingt es ihnen, ungestört ihre Raubzüge in den Ameisennestern durchzuführen. Als Präadaption dafür betrachten die Forscher den Besitz einer Drüse am Ende des Hinterleibs, durch die unangenehm riechende Stoffe gegen Angreifer abgegeben werden können. Doch einige der parasitisch lebenden Käfer besitzen neue Drüsen und neue Funktionen, was so wenig wie die veränderte Morphologie und das veränderte Verhalten als Folge einer Präadaption gewertet werden kann.
In jahrelanger mühevoller Forschungsarbeit gelang es Maruyama & Parker (2016), eine größere Anzahl verschiedener Arten dieser sozialparasitisch leben Käfer und ihrer nichtparasitischen Verwandten zu sammeln und genetisch zu untersuchen. Dabei fanden sie das höchst erstaunliche Ergebnis, dass die „Ameisenkäfer“-Arten so unsystematisch vorkommen, dass ihr Aussehen und Verhalten mindestens 12-mal unabhängig entstanden sein muss, wahrscheinlich noch sehr viel öfter. Dabei ist jede unabhängige „Ameisenkäfer“-Linie auf eine bestimmte geographische Region beschränkt und auf eine bestimmte Wanderameisen-Art spezialisiert.
Man könnte nun annehmen, dass der gemeinsame Vorfahr vor noch nicht allzu langer Zeit gelebt hat und dass aufgrund der noch nahen genetischen Verwandtschaft ein gemeinsames Potential zum „Ameisen-Look“ in diesem Vorfahren vorhanden war und in verschiedenen Linien abgerufen werden konnte. Doch die genetischen Daten weisen unter evolutionstheoretischen Voraussetzungen darauf hin, dass der gemeinsame Vorfahr bereits vor 105 Millionen Jahren gelebt hat. Außerdem habe sich das außergewöhnliche Aussehen und Verhalten der Käfer erst sehr spät – Dutzende von Millionen Jahren nach der Aufspaltung in die verschiedenen Linien – entwickelt – und wurde somit nicht auf der Basis eines gemeinsamen genetischen Potentials abgerufen. Evolutionstheoretisch muss man daher annehmen, dass die Nachahmung im Bau und Verhalten nicht nur jeweils unabhängig ausgeprägt wurde, sondern im Wesentlichen vielfach unabhängig ganz neu entstand.
Dass dieser Befund sehr unerwartet ist, geht aus den Worten von Pennisi (2016) hervor (in Übersetzung): „Man mag denken, dass die Anpassungen dieser Kurzflügler, wie sie nun bekannt sind, eine unwahrscheinliche Meisterleistung der Evolution bedeuten, die niemals wiederholt wurde. Doch damit würde man falsch liegen, …“ (Hervorhebung nicht im Original). Die Überraschung ist verständlich, da Evolution wie erwähnt ein zukunftsblinder Prozess ist. Daher kann man festhalten: Das Auftreten komplexer Konvergenzen widerspricht einer Entstehung auf natürlich-evolutivem Wege.
Doch Maruyama & Parker (2016) drehen den Spieß um: Aus der vielfachen Konvergenz folgern sie kurzerhand, es gebe in der Kurzflügler-Linie der Aleocharinae eine inhärente Fähigkeit zum Erwerb des ameisenartigen Körperbaus und Verhaltens. Daten zum Beleg dafür legen sie allerdings nicht vor, vielmehr ist die stillschweigende Voraussetzung von Evolution leitend für ihre Argumentation. Denn nur wenn man Evolution voraussetzt, kann man die Konvergenzen nachträglich als „vorhersehbar“ behaupten. Doch wären diese Konvergenzen keinesfalls vorhergesagt worden, wenn man sie nicht kennen würde! Die Vorhersage war eine andere: Wenn überhaupt kann sich ein solches System nur einmal evolutiv herausbilden (s. o. g. Zitat von Pennisi).
Dass hier (wie allgemein auch sonst) nur einer von zwei grundsätzlich möglichen Deutungsansätzen verfolgt wird – nämlich der naturalistische – wird auch durch Äußerungen zweier Wissenschaftler unterstrichen, die Pennisi am Schluss der Zusammenfassung zitiert. Diese Forscher legen – ohne weitere Begründung – das kreative Potential, das sich in der Flexibilität der Gestalt und des Verhaltens der Käfer zeigt, in die Evolution: Es zeige sich die „Virtuosität der Evolution“ einmal mehr (Eldredge) und „man spürt, wie einem die Kraft der Evolution – der natürlichen Selektion – in deine Augen starrt“ (Kronauer). Selektion ist zwar ein Thema der Arbeit von Maruyama & Parker (2016), aber ohne, dass auch nur ansatzweise erklärt wird, wie Selektion im Einzelnen zur Herausbildung der Ameisenkäfer beigetragen haben soll. Die Charakterisierungen von Eldredge und Kronauer sind Glaubensbekenntnisse über die Kraft natürliche Prozesse, für die es keine empirischen Belege gibt. Die vorliegenden Indizien passen dagegen viel besser zu einem Design-Ansatz. Wenn dieser jedoch von vornherein als Erklärungsform ausgeschlossen wird, gerät man bei den Erklärungsversuchen in Zirkelschlüsse und ist damit nicht ergebnisoffen unterwegs auf der Suche nach der plausibelsten Antwort.
Quellen
Maruyama M & Parker J (2016) Deep-time parallel evolution of myrmecoid syndrome in rove beetle symbionts of army ants. bioRxiv preprint first posted online Sep. 20, 2016; doi: http://dx.doi.org/10.1101/076315.
Pennisi E (2016) A new evolutionary classic. Science 354, 813.
Autor dieser News: Reinhard Junker
© 2016, http://www.genesisnet.info/schoepfung_evolution/n241.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
22.01.15 Schockierende Organe
Die Entstehung von elektrischen Organen in Fischen war schon zu Darwins Zeiten ein Thema, das Wissenschaftler faszinierte und herausforderte. Die Forscher konnten nicht erklären, warum bei Fischen aus unterschiedlichen, nicht näher verwandten Fischfamilien sehr ähnliche elektrische Organe vorkommen. Darwin rechnete damit, dass weiterführende Forschungen zu einer Lösung dieses Problems führen würden. Heute können auch genetische Daten zu Rate gezogen werden. Neue Studien darüber zeigen aber, dass das Rätsel der unabhängigen Entstehung elektrischer Organe eher noch größer wird.

Abb. 1: Zitteraal (Electrophorus electricus). (Wikimedia: FakirNL, CC BY-SA 4.0)
Eine Reihe von Fischen ist in der Lage, Elektrizität zu erzeugen, die sie für Orientierung, Kommunikation, Jagd und Verteidigung nutzen. Das geschieht mit Hilfe umgewandelter Muskelzellen, so genannten Elektrozyten, die sich unter der Haut befinden. Durch Hintereinanderschalten der Elektrozyten (vergleichbar mit einer Batterie) summiert sich die elektrische Ladung, und Fische wie der Zitteraal erreichen so Spannungen bis zu 600 Volt. Lindsay Traeger von der Michigan State University in East Lansing erklärt: „Die Fähigkeit zur Kontraktion* ging bei den Elektrozyten verloren und Proteine in den Zellmembranen änderten sich, so dass Ionen die Seiten wechseln, was zu einer elektrischen Ladung führt“ (nach Vieweg 2014). Dieser Weg ist ideal, um biogene Elektrizität zu erzeugen. Der Zitterrochen (Torpedinidae), der Zitteraal (Gymnotidae) oder der Nilhecht (Mormyridae) sind Beispiele dafür.
*Kontraktion (= Zusammenziehen von Muskeln)
Charles Darwin, der Begründer der modernen Evolutionsbiologie, befasste sich auch mit unabhängig entstandenen ähnlichen Organen wie beispielweise elektrische Organe. Er beobachtete, dass nicht verwandte Fische ähnliche Elektro-Organe nutzen. Er konnte keine angemessene Erklärung dafür finden und vertrat folgende zwei Argumente:
- Solange wir nur wenig über die Entstehung von elektrischen Organen wissen, kann nicht widerlegt werden, dass sie sich aus einer Vorläuferform entwickelt haben. „Darüber hinaus können wir momentan keine Erklärung geben, aber weil wir so wenig über den Gebrauch dieser Organe wissen, und nichts über ihre Verwendung und Anordnung bei den Stammeltern dieser Fische sagen können, wäre es extrem gewagt zu behaupten, dass keine sinnvolle Veränderung möglich gewesen wäre, durch die sich diese Organe schrittweise entwickelt haben.1
- Die elektrischen Organe seien in den verschiedenen Fischen nicht besonders ähnlich und können daher nicht als gleichartig betrachtet werden. Sie sind höchstens verhältnismäßig ähnlich in ihrer Funktion und müssen demnach nicht von einem gemeinsamen Vorfahren abstammen.2 Er schreibt: „Wenn wir das Thema noch näher betrachten, fällt auf, dass bei vielen Fischen mit elektrischen Organen diese sich in unterschiedlichen Bereichen des Körpers befinden, sie sich in ihrer Konstruktion unterscheiden sowie in der Anordnung der Anoden, in der Art und Weise, wie Elektrizität erzeugt wird und schlussendlich inwiefern sie mit Nervenzellen aus verschiedenen Quellen bestückt sind, was vielleicht der wichtigste aller Unterschiede ist. Daher können die Fische mit elektrischen Organen nicht als homolog, sondern nur analog betrachtet werden. Folglich gibt es keinen Grund anzunehmen, sie kämen von einem gemeinsamen Vorfahren.“
Darwin erwartete, dass weitere und verbesserte Forschung später zu einer Erklärung durch natürliche Prozesse führen würde. Doch durch die mittlerweile möglich gewordenen Erkenntnisse über die genetischen Grundlagen der elektrischen Organe wurde das Rätsel noch komplizierter. Denn Darwins zweites Argument über die nicht vorhandene Gleichartigkeit (Homologie) wurde dadurch hinfällig. Studien von Gallant et al. (2014) haben nämlich gezeigt, dass allen elektrischen Organen die gleichen Gene und gleiche zelluläre Entwicklungsabläufe zugrunde liegen. Der Leiter der betreffenden Studie, Jason Gallant von der Michigan State University, drückte diese Entdeckung mit folgenden Worten aus: „Wir haben herausgefunden, dass alle Elektro-Fische die gleiche genetische Werkzeugkiste nutzen, um ihre elektrischen Organe auszubilden“ (nach Vieweg 2014).
Wie aber kommt es, dass elektrische Organe in Fischen sechsmal unabhängig voneinander evolutiv, auf der Basis zukunftsblinder natürlicher Prozesse durch gleichartige Nutzung des genetischen Baukastens entstehen? Die Fische befinden sich in den unterschiedlichsten Lebensräumen wie dem Amazonas, Meeresgewässern oder Flüssen in Afrika. Man kann also davon ausgehen, dass es unterschiedliche Selektionsbedingungen gab, und trotzdem sind die Resultate auf der genetischen Ebene erstaunlich ähnlich. Das plötzliche Auftreten der Organe im Fossilbericht und die Tatsache, dass sie nur in sechs nichtverwandten Fischarten vorkommen und gleichzeitig bei deren näheren Verwandten keine Elektro-Organe vorhanden sind, sperren sich ebenfalls gegen eine Erklärung mittels natürlicher, blinder evolutionärer Prozesse. Schließlich sind die Anatomie und Physiologie, die die Nutzung dieser Organe ermöglichen, reichlich ausgeklügelt. Das bedeutet: Der evolutive Weg von Vorläuferstrukturen zu elektrischen Organen muss weit gewesen sein und viele Schritte erfordert haben. Wie aber kann ohne Zielvorgabe, die es in einer natürlichen Evolution nicht geben kann, dennoch ein derart genetisch ähnliches Ergebnis erreicht werden?
Erkenntnisse in diesem Forschungsgebiet könnten zukünftig auch dem Menschen nützlich sein. Die Herstellung von künstlichen Elektrozyten erlaubt vielleicht die Erzeugung von Strom in Körpern, was zu einer Reihe von Innovationen führen könnte. Co-Autor Michael Sussman von der University of Wisconsin-Madison drückte das so aus: „… wir können spannende Anwendungen entwickeln, die noch in den Sternen stehen“ (nach Vieweg 2014).
Literatur
Gallant JR, Traege LL et al. (2014) Genomic basis for the convergent evolution of electric organs. Science 344, 1522-1525, doi: 10.1126/science.1254432
Vieweg M (2014) Video der Woche: Buchstäblich schockierende Fische. http://www.wissenschaft.de/home/-/journal_content/56/12054/3923535/Video-der-Woche:-Heikles-Reiseziel-Mars/
Anmerkungen
1 „Beyond this we cannot at present go in the way of explanation; but as we know so little about the uses of these organs, and as we know nothing about the habits and structure of the progenitors of the existing electric fishes, it would be extremely bold to maintain that no serviceable transitions are possible by which these organs might have been gradually developed.” (http://www.birding.in/darwin/species_43.htm)
2 “But when we look at the subject more closely, we find in the several fishes provided with electric organs, that these are situated in different parts of the body,—that they differ in construction, as in the arrangement of the plates, and, according to Pacini, in the process or means by which the electricity is excited—and lastly, in being supplied with nerves proceeding from different sources, and this is perhaps the most important of all the differences. Hence in the several fishes furnished with electric organs, these cannot be considered as homologous, but only as analogous in function. Consequently there is no reason to suppose that they have been inherited from a common progenitor” (http://infidels.org/library/historical/charles_darwin/origin_of_species.html)
Autor dieser News: Michelle Noe
© 2015, http://www.genesisnet.info/schoepfung_evolution/n222.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.07.14 Von Anfang bis heute fast unverändert: Programmierter Zellabbau
Die molekularen Bestandteile, die dem programmierten Zellabbau (oder Zelltod, Apoptose) zugrunde liegen, sind zwischen Mensch und Koralle austauschbar. Aus evolutionsbiologischer Perspektive muss ein erstaunliches Ausmaß an evolutionärem Stillstand (Stasis; Konservierung) postuliert werden. Ausgerechnet unter den basalen und damit als stammesgeschichtlich sehr alt geltenden Vielzellern findet sich das vielseitigste Apoptose-System.
Ein wichtiger formbildender Prozess in der Ontogenese* ist der programmierte Zellabbau, also das physiologische (nicht krankhafte), programmierte Absterben von Zellen (Apoptose). Während es z. B. beim krankhaft bedingten Zelltod in der Regel zu einem unkontrollierten Aufquellen und Zerplatzen der Zelle oder zu einer unkoordinierten Zerstörung von Eiweißen oder Zellorganellen kommt (Nekrose), geschieht die Auflösung der Zellbestandteile und ihr Abbau im Rahmen der Apoptose hochgradig organisiert und physiologisch gesteuert. Apoptose dient damit immer dem Erhalt oder der koordinierten Entwicklung des Organismus. Beispielsweise werden Gewebestrukturen zwischen den ontogenetischen Anlagen von Fingern und Zehen kontrolliert zum Abbau gebracht oder durch die Auflösung der Zellen von Glaskörper und Linse wird die Lichtdurchlässigkeit der Augenlinse ermöglicht. Aber auch im ausgewachsenen Organismus spielt Apoptose eine wichtige Rolle, z. B. in der Kontrolle der Zellzahl und der Größe von Geweben, bei der Entfernung entarteter Zellen oder bei der Rückbildung der Gebärmutter nach der Geburt. Bei der Apoptose werden Nachbarzellen bzw. -gewebe durch austretende Gifte wie im Fall des o.g. krankhaft bedingten Zelltodes nicht geschädigt.
*Ontogenese (= individuelle Entwicklung von der befruchteten Eizelle bis zum ausgewachsenen Organismus)
Es ist schon länger bekannt, dass die molekulare Basis der Apoptose hochkonserviert ist, das heißt, sie ist bei sehr verschiedenen Organismengruppen sehr ähnlich, was evolutionstheoretisch so interpretiert wird, dass dieser Mechanismus stammesgeschichtlich sehr früh entstand und anschließend nicht mehr nennenswert verändert wurde. In einer neuen Untersuchung wiesen amerikanische Wissenschaftler (Quistada et al. 2014) nach, dass Apoptose auch bei riffbildenden Korallen vorkommt. Diese gelten im Rahmen der Evolutionsanschauung als Repräsentanten einer der ältesten Tiergruppen. Es zeigte sich, dass alle wesentlichen Komponenten des Apoptose-Programms bei Korallen vorhanden sind und denen des Menschen gleichen. Dabei spielen die Tumornekrosefaktor (TNF)-Superfamilien (TNFRSF/TNFSF) eine zentrale Rolle. Die Wissenschaftler untersuchten 53 Proteine der TNFRSF- und TNFSF-Familien in der riffbildenden Koralle Acropora digitifera und stellten große Sequenzähnlichkeiten mit den entsprechenden Proteinen des Menschen fest, besonders bezüglich der aktiven Zentren dieser Eiweiße. Ein Vergleich mit der Taufliege Drosophila ergab, dass in dieser Linie im Gegensatz zur Linie zum Menschen wichtige Teile der TNF-Superfamilie fehlen, was evolutionstheoretisch als Verlustentwicklung interpretiert wird. Zuvor war angenommen worden, dass die Zahl der Mitglieder der TNF-Superfamilie nach der Trennung der Linie der Wirbeltiere von der der Wirbellosen zugenommen habe. Nun hat sich herausgestellt, dass im Gegenteil die Korallen das vielseitigste Repertoire der TNF-Superfamilien besitzen.
Die große Ähnlichkeit der molekularen Komponenten des Apoptose-Prozesses bei Korallen und Menschen wurde eindrucksvoll demonstriert durch die Einführung eines menschlichen TNF (HuTNFα) in die Korallen. Es zeigte sich, dass es direkt an die Korallen-Zellen bindet, dadurch die Caspase-Aktivität erhöht, was (über eine Kaskade von Prozessen) zur für die Apoptose typischen Bläschenbildung und zum Zellabbau führt. (Capsasen sind Enzyme, die Proteine durch Hydolyse abbauen und bei der Apoptose eine besondere Rolle spielen.) Umgekehrt führte ein TNF von Korallen (AdTNF1) bei menschlichen Zellen zu einem signifikant häufigeren physiologisch hervorgerufenen Zelltod. TNF von Korallen und Menschen sind also austauschbar.
Beispiele wie der zelluläre Apoptose-Mechanismus zeigen einerseits ein erstaunliches Ausmaß an Konservierung eines zellulären Prozesses und seiner Bestandteile; in evolutionstheoretischer Interpretation muss ein Stillstand bezüglich der Evolution dieses Mechanismus von mindestens 550 Millionen Jahren angenommen werden. Andererseits werfen sie aber auch die Frage auf, wann und wie ein solcher Mechanismus überhaupt evolutiv entstand. Denn wenn er offenkundig nicht nennenswert veränderbar ist, wie sollen dann hypothetische Vorstufen in irgendeiner Weise funktional und damit existenzfähig sein? Die Daten zeigen einmal mehr: Abruptes Auftreten und dann weitgehend Konstanz eines zentralen biologischen Struktur- und Funktionsgefüges.
Literatur
Quistada SD, Stotlanda A et al. (2014) Evolution of TNF-induced apoptosis reveals 550 My of functional conservation. PNAS early ed, doi: 10.1073/pnas.1405912111
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n213.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
05.06.14 Rippenquallen – eine Welt für sich
Die Untersuchung des Erbguts der Seestachelbeere (Pleurobrachia bachei; Ctenophora, Rippenquallen) sowie Daten zur Transkription (= Ablesen des Erbguts) aus zehn weiteren Rippenquallen-Arten bestätigten frühere Deutungen, wonach die Rippenquallen aus evolutionärer Perspektive zu den ursprünglichsten Tieren gehören und ihr komplexes Nervensystem unabhängig vom Nervensystem anderer Tiere entstand. Evolutionsbiologen zeigen sich über die Ergebnisse überrascht. Die Befunde können verschieden interpretiert werden, jede evolutionstheoretische Deutung aber wirft schwerwiegende ungelöste Fragen auf.
In der Diskussion um Schöpfung und Evolution wird häufig behauptet, es gebe keine Befunde, die die Evolutionsanschauung in Frage stellen würden, und die Evolutionstheorie1 werde laufend bestätigt. Das hört sich gut an, aber vermutlich würde man sich kaum darauf verständigen können, was überhaupt widersprechende Befunde wären. Die zweite Behauptung, es gebe immer wieder Bestätigungen, kann man hingegen besser überprüfen, weil Biologen wiederholt Erwartungen formuliert haben, die sie aus der Evolutionstheorie abgeleitet haben. Eine solche Erwartung war, dass ähnliche komplexe Organsysteme nicht zweimal unabhängig entstehen könnten. Nicht umsonst gelten (komplexe) Ähnlichkeiten als Hinweise auf gemeinsame Abstammung und bilden die Basis für Stammbaumrekonstruktionen. So wird Leonid Moroz, Neurowissenschaftler an der Universität von Florida, bezüglich der Entstehung des Nervensystems zitiert: „Jeder ist der Auffassung, dass diese Art von Komplexität nicht zweimal auftreten könne, aber dieser Organismus legt nahe, dass es doch passiert ist“ (Callaway 2014).2
Die Rede dabei ist von der Rippenqualle Pleurobrachia bachei, bekannt als Seestachelbeere, und darüber hinaus vom ganzen Tierstamm der Rippenquallen (Ctenophora). Moroz bezeichnet diese ungewöhnlichen Organismen als „Aliens, die auf die Erde gekommen sind“ (Callaway 2014). Rippenquallen sind Prädatoren (Räuber) des Meeres und bewegen sich mit Hilfe eines Bündels von Cilien fort. Ihre Beute fangen sie mit innervierten Tentakeln, die von klebrigen Zellen (sog. Colloblasten) gesäumt sind, und verschlingen sie mit ihrem Mund, dem sich ein sackartiger Darm anschließt. Die Rippenquallen besitzen ein Nervennetz mit regionalen Spezialisierungen, darunter Sinnesorgane für Lichtrezeption und die Schwerkraftempfindung. Es ist auch bei der Wahrnehmung der Beute und der Bewegung der Muskultur im Einsatz.
Das Nervensystem ist es, das für die Überraschung seiner mutmaßlich zweimaligen unabhängigen Entstehung sorgt. Schon die Untersuchung des Genoms (= komplettes Erbgut) der Rippenqualle Mnemiopsis leidyi hatte nahelegt, dass das Nervensystem der Rippenquallen unabhängig vom Nervensystem anderer Tiere entstanden sei (Ryan et al. 2013). Dafür gibt es zwei Gründe: Die Rippenquallen müssen aufgrund von Gen- und Proteinvergleichen an die Basis des Tierreichs gestellt werden, und nicht – wie zuvor angenommen – die einfacher gebauten Schwämme (Porifera) und Plattentiere (Placozoa), die kein Nervensystem besitzen. Und zweitens, das Nervensystem der Rippenquallen unterscheidet sich in vielerlei Hinsicht von dem aller anderen Tiere mit Nervensystem. Beides wurde nun durch die Entschlüsselung des Genoms der Seestachelbeere Pleurobrachia bachei bestätigt, sowohl die basale Position der Rippenquallen als auch die großen Unterschiede in ihrem Nervensystem gegenüber anderen Organismen. Einige Bespiele dafür: Viele der sonst vorhandenen Komponenten des Nervensystems (Regulationsgene, Strukturgene, Signalmoleküle) fehlen bei der Seestachelbeere. Während in fast allen sonstigen Nervensystemen dieselben zehn primären Neurotransmitter* im Einsatz sind, nutzt Pleurobrachia deren nur einen oder zwei. Außerdem stellte sich heraus, dass einige biologisch aktive molekulare Substanzen, die bei anderen Organismen spezifisch für Nervenzellen sind, bei der Seestachelbeere zwar auch vorkommen, aber überraschenderweise nicht in Nervenzellen, sondern in anderen Geweben genutzt werden (Moroz et al. 2014, Hejnol 2014).
*Neurotransmitter (= Botenstoffe an den Kontaktstellen von Nervenzellen)
Bemerkenswert ist weiter, dass Pleurobrachia weder microRNA besitzt (bei den meisten anderen Tieren ein wichtiger Teil der Genregulation) noch die bei anderen Tieren wichtigen Hox-Gene (die u. a. eine Schlüsselrolle bei der Formung der Körperachsen spielen).
Für die evolutive Einordnung der Rippenquallen ergibt sich aus der Befundsituation ein Trilemma:
- Die zweimalige unabhängige Entstehung des Nervensystems widerspricht bisherigen Erwartungen im Rahmen evolutionstheoretischer Deutungen (s. o.). Es ist ohnehin unbekannt, wie ein Nervensystem evolutiv entstanden ist; nun ist man damit konfrontiert, dass dies zweimal unabhängig geschehen sein muss.
- Es kann alternativ auch angenommen werden, dass ein Nervensystem nur einmal entstanden ist und die Schwämme und Plattentiere es (samt anderen Merkmalen) wieder verloren haben. Diese Sichtweise steht vor dem Problem, weshalb ein derart nützliches komplexes Organsystem komplett aufgegeben worden sein sollte.
- Es ist nicht endgültig gesichert, dass die Rippenquallen tatsächlich an der Basis des evolutionären Tier-Stammbaums stehen. Früher waren die Rippenquallen und die Nesseltiere aufgrund mancher Gemeinsamkeiten zu den Hohltieren zusammengefasst worden. Das Nervensystem könnte auch in diesem Fall nur einmal entstanden sein. Da aber die Nervensysteme von Rippenquallen und anderen Tieren wie dargestellt in vielerlei Hinsicht verschieden sind, stellt sich dann die Frage, wie und warum der enorme Umbau des Rippenquallen-Nervensystems in das Nervensystem der anderen Tiere erfolgte.
Hejnol (2014) versucht in seinem Kommentar aus dem Bock einen Gärtner zu machen und meint, die neue Sicht von der unabhängigen Entstehung zweier unterschiedlicher Nervensysteme sei ein Schlag gegen die anthropozentrische Sicht, dass komplexe Merkmale allmählich entstanden seien, was schließlich mit der Entstehung des Menschen gipfelte, und dass komplexe Merkmale nicht zweimal evolvieren würden. Aber so einlinig funktioniere Evolution nicht, sondern sei ein Prozess, der in allen Linien voranschreite.
Der Kommentar ist sehr erhellend. Aus ihm geht hervor, welche grundsätzlichen Unterschiede bezüglich des Grundverständnisses von Evolution innerhalb der Evolutionsbiologen existieren. Die Behauptung, Evolution führe zu einem Wandel mit ähnlichen komplexen Resultaten in verschiedenen Linien, ist gewagt und beruft sich auf Vorgänge, die bislang niemand kennt und nicht durch Faktenwissen gestützt ist. Und weshalb soll die Erwartung, dass komplexe Organsysteme nicht zweimal entstehen, anthropozentrisch sein? In Wirklichkeit wurde diese Erwartung aus der Kenntnis der bislang ermittelten Evolutionsmechanismen abgeleitet und propagiert (s. o.); diese Erwartung wurde hier enttäuscht. Wenn eine Organismengruppen wie die hier vorgestellten Rippenquallen trotz vieler Gemeinsamkeiten klar von anderen abgrenzbar sind und keine natürlichen Mechanismen bekannt sind, wie diese Grenzen evolutionär überbrückt werden können, muss dies als schwerwiegendes Problem innerhalb des evolutionsbiologischen Forschungsprogramms auch so benannt werden.
Unabhängig davon besteht als Alternative die Deutungsmöglichkeit, dass die Grenzen primär existierten, d. h. nicht die Folge einer wie auch immer gedeuteten Evolution, sondern Folgen einer Schöpfung sind.
Literatur
Callaway E (2014) Jelly genome mystery. Nature 509, 411.
Hejnol A (2014) Excitation over jelly nerves. Nature, doi:10.1038/nature13340.
Morot LL, Kocot KM et al. (2014) The ctenophore genome and the evolutionary origins of neural systems. Nature doi:10.1038/nature13400.
Rokas A (2013) My oldest sister is a walnut? Science 342, 1327-1329.
Ryan F, Pang K et al. (2013) The genome of the ctenophore Mnemiopsis leidyi and its implications for cell type evolution. Science 342, 1242592, doi: 10.1126/science.1242592
Anmerkungen
1 zum Begriff: hier gemeint als Abstammung aller Lebewesen von andersartigen Vorfahren.
2 „Everyone thinks this kind of complexity cannot be done twice,“ Moroz says. „But this organism suggests that it happens“ (Callaway 2014).
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n212.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
17.01.14 Immer verworrener: Verwandtschaftsbeziehungen der ältesten Vielzeller
Erstmals wurde das gesamte Erbgut einer Rippenqualle sequenziert. Die Analyse der Daten ergab mehrere Überraschungen: Rippenquallen stehen anstelle der einfacher gebauten Schwämme an der Basis des Tierreiches. Für den gemeinsamen Vorfahren muss wahrscheinlich die Existenz eines Nervensystems angenommen werden, das die Schwämme wieder verloren hätten. Die Entstehung von Mesoderm und Muskelzellen muss zweimal unabhängig erfolgt sein. Die Befunde sind aus einer Schöpfungsperspektive leichter verstehbar.
In ihrem Buch „The Cambrian Explosion“ beschreiben Erwin & Valentine (2013, 266) die ursprüngliche Erwartung, die man über genetische Ähnlichkeiten von Organismen hatte. Vielzellige Organismen mit verschiedenen Bauplänen sollten kaum Ähnlichkeiten auf genetischer Ebene ausweisen: „Da Vielzeller-Baupläne entlang auseinanderstrebender Pfade evolvierten, um eine reiche morphologische Verschiedenartigkeit hervorzubringen, die wir sowohl bei fossilen als auch heutigen Faunen beobachten, schien es vernünftig zu sein, entsprechend große Unterschiede in ihrem Erbgut zu erwarten. … Es schien wahrscheinlich zu sein, dass beispielsweise Gliederfüßer und Chordaten nur wenige Gene teilen.1 Vor etwa 50 Jahren meinte Ernst Mayr, der „Darwin des 20. Jahrhunderts“, die Suche nach homologen Genen außer bei nah verwandten Formen sei aussichtslos (Mayr 1963, 609; vgl. Carroll 2005, 9).2
Heute wissen wir, dass es sich ganz anders verhält. Viele Regulationsgene, Gen-Netzwerke und Signalwege sind bei Organismen mit den unterschiedlichsten Bauplänen (von Quallen bis Wirbeltieren und Gliederfüßern u. v. a.) sehr ähnlich. Aus evolutionärer Perspektive muss aufgrund dieses eigentlich unerwarteten Befunds angenommen werden, dass der gemeinsame Vorfahr (mit einem einfachen Körperbauplan!) genetisch ausgesprochen komplex gewesen sein muss. Denn alle Gene, die in mehreren Organismen mit verschiedenen Bauplänen vorkommen, müssen in diesem ersten, einfachen und hypothetischen Organismus vorhanden gewesen sein. Die so bereits vorhandenen Gene sollen dann in den verschiedenen Linien für neue Zwecke rekrutiert worden sein. Es hat sich eingebürgert, dabei von genetischen Baukästen („tool-kits“) zu sprechen. Es scheint so, dass „Teile“ aus diesen „Baukästen“ je „nach Bedarf“ verwendet oder weggelassen wurden.3
Eine freie Kombinierbarkeit von Merkmalen („mal so, mal so“) stört bei der Erstellung von Stammbäumen, ist aber dagegen in ein Schöpfungskonzept gut zu integrieren. Ein Stammbaum kann zwar immer konstruiert werden; das erfordert aber in zunehmendem Maße entweder die Annahme von Konvergenzen („ist mehrfach entstanden“) oder von Rückentwicklungen („ist wieder entfallen“). Es gibt mittlerweile zahlreiche Beispiele, bei denen sicher geglaubte Homologien (als abstammungsbedingt interpretierte Ähnlichkeiten) wegen neuer Daten zu Konvergenzen (mehrfach unabhängig entstanden) uminterpretiert wurden mussten.
Dass genetische Daten immer wieder „klassischen“ Verwandtschaftsverhältnissen, die aus morphologischen (= gestaltlichen) Merkmalen erschlossen wurden, widersprechen, zeigte jüngst auch die Veröffentlichung des Erbguts der Rippenqualle Mnemiopsis leidyi. In dieser Arbeit wurde erstmals das komplette Erbgut einer Rippenqualle (Stamm Ctenophora) untersucht (Ryan et al. 2013). Trotz des deutschen Namens Rippenqualle haben diese nichts mit den echten Quallen aus dem Stamm der Nesseltiere zu tun (s. u.). Die Untersuchung der Rippenquallen war interessant, weil diese Organismen zu den einfachsten Tiergruppen gehören und an die Basis der Vielzeller gestellt werden. Dazu zählen außer den Rippenquallen auch die Schwämme (Porifera), die winzigen Placozoa (Plattentiere; durch die einzige Art Trichoplax vertreten) und die Nesseltiere (Cnidaria).
Rippenquallen unterscheiden sich von anderen Quallen, die zu den Nesseltieren gehören, in der zweiseitigen Symmetrie („rechte und linke“ Körperhälften statt „runder“ Radiärsymmetrie); die Körperachse ist durch einen Mund am einen Ende und ein Organ, das die Schwerkraft wahrnimmt, am anderen Ende definiert. Anders als die Placozoa und die Schwämme, aber gemeinsam mit den Nesseltieren besitzen die Rippenquallen Muskeln und Nervengewebe, letzteres ist als diffuses Netz organisiert. Schwämme, und Nesseltiere bilden in der Embryonalentwicklung nur zwei Keimblätter – Ektoderm und Endoderm – aus, jedoch kein Mesoderm (aus dem z. B. das Muskelgewebe gebildet wird), wie das sonst der Fall ist. Verschiedene Untersuchungen führten je nach Gewichtung der Merkmale zu unterschiedlichen Verwandtschaftsbeziehungen dieser vier Gruppen untereinander und zu den (höherentwickelten) Bilateria („Zweiseitentieren“). Ryan et al. (2013) bilden sechs verschiedene Verwandtschaftsbäume ab. Das zeigt deutlich die Schwierigkeiten, eine widerspruchsfreie Phylogenie entwerfen zu können.
Überraschenderweise sprechen nun die Sequenzdaten der 16.548 identifizierten Gene4 von Mnemiopsis leidyi dafür, dass die Rippenquallen der erste Zweig des Tier-Stammbaums sind und nicht die viel einfacher gebauten Schwämme, was man bisher annahm (Ryan et al. 2013). Die zuvor meist angenommene engere Verwandtschaft von Rippenquallen und Nesseltieren hat sich auch nicht bestätigt. Da die Rippenquallen ein Nervensystem besitzen, muss evolutionstheoretisch angenommen werden, dass der gemeinsame Vorfahr ebenfalls ein Nervensystem besaß. M. leidyi teilt zudem viele Gene des Nervensystems mit den Schwämmen, obwohl diese kein Nervensystem besitzen (Ryan et al. 2013). Daraus schließen die Forscher, dass die Schwämme das Nervensystem verloren haben. Wieder einmal soll also, entgegen früherer Erwartungen und Vorhersagen, die Abfolge von komplex nach einfach verlaufen sein (vgl. Von komplex nach einfach?, https://www.genesisnet.info/index.php?News=166). Die Alternative dazu wäre anzunehmen, dass das Nervensystem zweimal unabhängig entstanden ist. Das könnte auch einige Besonderheiten des Rippenquallen-Nervensystems verständlich machen5, wirft jedoch wieder andere Fragen auf (s. u.).
Eine weitere Überraschung ist der Befund, dass Rippenquallen zwar ein Mesoderm besitzen und darüber hinaus auch Muskeln ausbilden, sie dafür aber völlig andere Gene nutzen als die Bilateria. Auch verläuft der Bildungsprozess der Muskelzellen von erwachsenen Tieren ganz anders als bei den Bilateria. Das bedeutet, dass die auch genetische Maschinerie für die Entwicklung des Mesoderms und für die Bildung von Muskelzellen überraschenderweise zweimal unabhängig entstanden sein müsste (Ryan et al. 2013; vgl. Rokas 2013, 1329).
Die neuen Befunde wurden von Rokas (2013) auf eine interessante Weise kommentiert. Seiner Meinung nach ist die evolutionstheoretische Vorstellung einer linearen Entwicklung von „einfach“ zu „komplex“ eine teleologische, also zielorientiert. Das Ziel wäre der komplexe Organismus, der Beginn ein einfacher. Diese „teleologisch getränkte“ Vorstellung könne nun endlich aufgegeben werden – als ob man nur darauf gewartet hätte. Doch gerade neue Erklärungsansätze können ihrerseits in Richtung Teleologie gedeutet werden, denn: Woher kam der anzunehmende komplexere Vorfahr? Rokas spricht von einer „Fabrik des Ursprungs der Tiere“, zu der auch mehrfache unabhängige Entstehung und Verlust von Genen, Stoffwechselwegen und sogar spezialisierten Zelltypen gehören, wie in dieser Studie die Nerven- und Muskelzellen, die Ryan et al. (2013) als „hochentwickelt“6 bezeichnen. Doch jede Annahme einer Rückentwicklung (z. B. Verlust von Nervengewebe bei Schwämmen) verschiebt das Problem der vorherigen Entstehung (es muss vorher etwas gegeben haben, was verloren gehen konnte) nur noch weiter in die Vergangenheit. Die Folge ist, dass die hypothetischen komplexen gemeinsamen Vorfahren „überdesigned“ erscheinen (so Erwin & Valentine 2013, 336), da sie mit mehr genetischen Möglichkeiten ausgestattet zu sein scheinen als sie tatsächlich benötigten. Conway Morris (2000, 1) sprach übrigens in diesem Zusammenhang von „Tieren in Wartestellung“. Doch „warten“ erfordert Wissen über ein zukünftiges Ereignis. Wozu sollten Organismen genetischen „Ballast“ anhäufen? Die unabhängige Entstehung ähnlicher komplexer funktionaler Strukturen war evolutionstheoretisch aus guten Gründen bisher für sehr unwahrscheinlich gehalten worden, eben weil richtungslose Prozesse sich ähnelnde, ausgefeilte Resultate nicht erwarten lassen. Hier wird unter der Hand blinden Prozessen schöpferisches Potential oder eine Art prophetische Weitsicht zugebilligt.
Man kann die Befunde auch aus einer Schöpfungsperspektive deuten: Wenn Merkmale grundsätzlich frei kombinierbar sind, müssen sie nicht in einen widerspruchsfreien Stammbaum eingepasst, oder besser gesagt eingezwängt werden. Konvergenzen sind zu erwarten und hypothetische, unverständlich komplexe Vorfahren erübrigen sich.
Literatur
Carroll SB (2005) Endless Forms Most Beautiful. The New Science of Evo Devo and the Making of the Animal Kingdom. London.
Conway Morris S (2000) Evolution: Bringing molecules into the fold. Cell 100, 1-11.
Erwin DH & Valentine JW (2013) The Cambrian explosion. The construction of animal biodiversity. Greenwood Village, Colorado.
Mayr E (1963) Animal Species and Evolution. Harvard University Press.
Rokas A (2013) My oldest sister is a walnut? Science 342, 1327-1329.
Ryan F, Pang K et al. (2013) The genome of the ctenophore Mnemiopsis leidyi and its implications for cell type evolution. Science 342, 1242592, doi: 10.1126/science.1242592
Anmerkungen
1„As metazoan body plans evolved along many diverging pathways to produce the rich morphological disparity observed both in fossil and living faunas, it seemed reasonable to expect correspondingly large divergences in their genomes. … it seemed likely that arthropods and chordates, for example, shared few genes.“
2 „Much that has been learned about gene physiology makes it evident that the search for homologous genes is quite futile except in very close relatives. If there is only one efficient solution for a certain functional demand, very different gene complexes will come up with the same solution, no matter how different the pathway by which it is achieved. The saying ‘Many roads lead to Rome’ is as true in evolution as in daily affairs.“
3 „Die sehr ähnliche Architektur des Erbguts, die dem Arm, mit dem ich diesen Artikel schreibe, und dem Flügel der vorbeischwebenden Fliege zugrunde liegt, bringt die nüchterne Möglichkeit ins Spiel, dass Vieles in der Architektur der Organismen von einem Satz von Baukästen abhängt, von denen jeder bei Bedarf geöffnet wird“ (Conway Morris 2000, 9).
4 Ca. 6.000 Gene weniger als beim Menschen, aber mehr als bei der Taufliege Drosophila.
5 „It appears that much of the genetic machinery necessary for a nervous system was present in the ancestor of all extant animals. This pattern suggests that a less elaborate nervous system was present in the metazoan ancestor and was secondarily reduced in placozoans and sponges. The alternative is that neural cell types arose independently in both the ctenophore lineage and the lineage that led to cnidarians and bilaterians, which might explain some of the unique aspects of the ctenophore nervous system“ (Ryan et al. 2013, 7).
6 „sophisticated”
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n206.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
23.10.13 Molekulare Konvergenzen in unerwartetem Ausmaß
Dass ähnliche Strukturen bei Lebewesen unabhängig (konvergent) entstehen können, ist lange bekannt und die Beispiele dafür nehmen rasant zu, so dass es immer problematischer wird, anhand von Bauplanähnlichkeiten stammesgeschichtliche Beziehungen zu ermitteln. Die Ergebnisse einer groß angelegten Studie legen nahe, dass die Konvergenzen auch im molekularen Bereich erheblich sein können.
Man hat sich daran gewöhnt: Viele ähnliche morphologische (= gestaltliche) Merkmale oder Fähigkeiten nicht näher verwandter Organismen sind mehrfach unabhängig entstanden. Dieses Phänomen wird als Konvergenz bezeichnet. Eigentlich gelten Bauplanähnlichkeiten als Hinweise auf gemeinsame Abstammung (man spricht dann Homologien), aber es gibt so viele Ausnahmen, dass strukturell homologe Ähnlichkeiten nicht mehr ohne weiteres als Belege für gemeinsame Abstammung gewertet werden können (viele Beispiele z. B. unter http://www.thegreatstory.org/convergence.pdf und www.mapoflife.org; vgl. Braun 2012)
Durch die taxonomische* Forschung in der Biologie sind viele vormals als Homologien interpretierte Ähnlichkeiten durch Gewinnung neuer Daten zu Konvergenzen „mutiert“. Die Daten, die zu Neubewertungen führten, stammen oft (aber nicht nur) aus der Molekularbiologie. Sequenzvergleiche von Proteinen und Genen und neuerdings ganzer Genome** ergaben in zahllosen Fällen Ähnlichkeitsbeziehungen, die sich deutlich von den Merkmalsmustern unterscheiden, die auf morphologischen Merkmalen beruhen. Heute weiß man, dass es verbreitet Konflikte „Morphologie versus Moleküle“ gibt. Da in Konfliktfällen den molekularen Daten in der Regel größeres Gewicht eingeräumt wird, müssen die widersprechenden morphologischen Merkmale konvergent entstanden sein.
*taxonomisch (= die Ordnung der Lebewesen aufstellende)
**Genom (= komplettes Erbgut eines Individuums)
Mittlerweile hat sich herausgestellt, dass Konvergenzen auch im molekularen Bereich in beträchtlichem Ausmaß vorkommen. So sind beispielsweise die Aminosäuresequenzen des Proteins Prestin, das beim Hörvorgang beteiligt ist und bei der Echoortung von Fledermäusen und Walartigen besondere Bedeutung hat1, bei diesen beiden Gruppen nahezu identisch. Wale und Fledermäuse erscheinen in einem auf Prestin-Daten basierenden Dendrogramm (Ähnlichkeitsbaum) daher als eine gemeinsame Gruppe2, obwohl beide Gruppen unter den Säugetieren sonst überhaupt nicht näher verwandt sind (Li et al. 2010; Liu et al. 2010). Nach den DNA-Sequenzdaten des Prestin-Gens gruppieren sich die Wale und Fledermäuse dagegen weitgehend gemäß dem ‚üblichen’ Arten-Stammbaum. Innerhalb der Fledermäuse passen auch die Nukleotid-Sequenzdaten nicht zu den sonst anerkannten Verwandtschaftsbeziehungen.
In einer groß angelegten Untersuchung ist eine Arbeitsgruppe um den Evolutionsbiologen Stephen Rossiter (Queen Mary, University of London) der Frage nachgegangen, ob weitere molekulare Konvergenzen bei Fledermäusen und Walen vorliegen (Parker et al. 2013). Beide Gruppen haben unter den Säugetieren nicht viel gemeinsam, mit einer Ausnahme: Sie haben anders als alle anderen Säuger ein ausgefeiltes Echoortungssystem, das mit ausgestoßenen Ultraschalllauten arbeitet und viele Gemeinsamkeiten aufweist.3 In der Studie der Arbeitsgruppe von Rossiter wurden tatsächlich weitere molekulare Konvergenzen nachgewiesen – zur Überraschung der Wissenschaftler in einem enormen Ausmaß. Knapp 200 Gene weisen konvergente Gemeinsamkeiten bei Walen und Fledermäusen auf, von denen viele mit der Echoortung direkt oder indirekt in Verbindung stehen – die Forscher hatten allenfalls ein Dutzend erwartet. Einige der konvergent veränderten Gene stehen im Zusammenhang mit dem Sehsinn; bei einer Reihe von ihnen ist die Funktion unbekannt oder die Beziehung zur Echoortung unklar.
Die Wissenschaftler nutzten die Genomdaten von sechs nicht näher verwandten Fledermäusen, von denen zwei keine Echoortung betreiben, außerdem vom Großen Tümmler und von weiteren Säugetierarten. Daraus ermittelten sie etwa 2300 Gene, die in allen diesen Arten in einfacher Ausführung (single copy) vorkommen, und verglichen diese untereinander. Auf diese Weise gelangten sie zum genannten Ergebnis von knapp 200 Genen, die unabhängig voneinander in derselben Weise (konvergent) verändert waren.
Dieses enorme Ausmaß konvergenter Veränderungen stößt auf allgemeine Überraschung. Zuvor war aus evolutionstheoretischer Sicht erwartet worden, dass die molekularen Grundlagen der Echoortungssysteme der beiden so verschiedenen Säugergruppen ganz verschieden sein müssten, da Mutationen als „Rohmaterial“ für evolutionäre Neuheiten richtungslos erfolgen. Für eine gleichsinnige Änderung auf breiter Front können keine plausiblen Ursachen angegeben werden. Gewöhnlich wird bei konvergenter Ähnlichkeit auf gleichartige Selektionsdrücke als Ursache verwiesen, doch zum einen handelt es sich dabei generell um eine notwendige, nicht aber hinreichende Voraussetzung für gleichartige Veränderungen. Hier kommt zum anderen das schiere Ausmaß gleichsinniger Abwandlungen hinzu. Joe Parker, Erstautor der Veröffentlichung der erstaunlichen Ergebnisse, kommentiert: „Wir wissen, dass natürliche Auslese eine wirkungsvolle Triebfeder der Evolution von Gensequenzen ist, doch der Nachweis von so vielen Beispielen, wo sie nahezu identische Resultate bei den Gensequenzen bei Tieren ohne jede Verwandtschaft verursacht, ist erstaunlich.“
Die Tatsache, dass molekulare Konvergenzen verbreitet sein können, bringt ein Problem mit sich, das von den morphologischen Konvergenzen längst vertraut ist: Ähnlichkeiten, die nach üblichen Kriterien als homolog interpretiert werden, können nicht sicher als Indikatoren für die Existenz gemeinsamer Vorfahren gewertet werden bzw. sie können irreführend sein, wenn man dies doch tut. Der Genetiker Todd Castoe wird mit den Worten zitiert, dass derzeit keine Handhabe zur Verfügung stünde, wie man mit dieser Situation umgehen kann (http://news.sciencemag.org/biology/2013/09/bats-and-dolphins-evolved-echolocation-same-way#disqus_thread). Er äußert allerdings Bedenken wegen der Untersuchungsmethode, die nur indirekte Hinweise auf das Vorliegen von molekularen Konvergenzen erlaubt. Hier können weitere Untersuchungen aber Aufschluss geben.
Literatur
Braun HB (2012) Warten auf einen neuen Einstein. Stud. Int. J. 19, 12-19.
Li G, Wang J, Rossiter SJ, Jones G, Cotton JA, Zhang S (2008) The hearing gene Prestin reunites echolocating bats. Proc. Natl. Acad. Sci. 105, 13959-13964.
Li Y, Liu Z, Shi P & Zhang J (2010) The hearing gene Prestin unites echolocating bats and whales. Curr. Biol. 20, R55-R56.
Liu Y, Cotton JA, Shen B, Han X, Rossiter SJ & Zhang S (2010) Convergent sequence evolution between echolocating bats and dolphins. Curr. Biol. 20, R53-R54.
Parker J, Tsagkogeorga G, Cotton JA, Liu Y, Provero P, Stupka E & Rossiter SJ (2013) Genome-wide signatures of convergent evolution in echolocating mammals. Nature, doi:10.1038/nature12511.
Teeling EC (2009) Hear, hear: the convergent evolution of echolocation in bats? Trends Ecol. Evol. 24, 351-354.
______________
1 Prestin wird beim Hörvorgang benötigt und codiert für ein Transmembran-Motorprotein, welches das elektrisch vermittelte Bewegungsvermögen der äußeren Haarzellen in der Schnecke steuert und damit für die Sensitivität der Schnecke im Säugerohr verantwortlich ist. Es scheint von besonderer Bedeutung für die Wahrnehmung höherer Frequenzen und für selektives Hören zu sein; beides ist für die Echoortung wichtig (Teeling 2009, 353; Li et al. 2008, 13959).
2 „Bei der aktuellen Untersuchung wurden auch echoortende Zahnwale einbezogen, und basierend auf der Sequenz von Prestin, bestanden mehr Ähnlichkeiten zwischen den echoortenden Fledermäusen und Walen als zwischen den nicht-echoortenden, aber viel näher verwandten Fledermausarten. Dies legt nahe, dass die Veränderungen des Prestinproteins wichtig sind für ein funktionstüchtiges Echoortungssystem.“ (http://www.science-meets-society.com/wissenschaftsnews/die-konvergenz-der-echoortung/)
3 „Doch die Mechanismen der Echoortung ähneln sich stark. Trotz so unterschiedlicher Erfordernisse, wie eine Anpassung an die Dichteunterschiede der Medien wie Luft und Wasser und auch der überbrückten Distanz, von einigen Metern bei Fledermäusen zu mehreren hundert Meter bei Delphinen, finden sich beeindruckende Übereinstimmungen in den anatomischen Bausteinen, die für die Verarbeitung der Echosignale zuständig sind.“ (http://www.science-meets-society.com/wissenschaftsnews/die-konvergenz-der-echoortung/)
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n200.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
20.07.13 Morphologie und Moleküle in scharfem Widerspruch
Unter der Annahme von Evolution war erwartet worden, dass Ähnlichkeitsbäume (= Cladogramme), die auf „traditionellen“ anatomischen und morphologischen (= gestaltlichen) Merkmalen beruhen, mit Ähnlichkeitsbäumen harmonieren, die auf molekularen Daten basieren (also auf Sequenzvergleichen von Proteinen und Nukleinsäuren). In vielen Fällen haben jedoch molekulare Analysen sicher geglaubte traditionell begründete Verwandtschaftsverhältnisse in Frage gestellt. Moleküle und Morphologie passen oft nicht ohne weiteres zusammen. Eine besonders schwerwiegende Diskrepanz wurde unlängst bei den Eidechsenartigen beschrieben.
Als die ersten Cladogramme (baumförmige Darstellung von Ähnlichkeitsbeziehungen) auf der Basis von Proteinvergleichen verschiedener Arten veröffentlicht wurden, schienen sie die Ähnlichkeitsmuster der „klassischen“ Taxonomie zu bestätigen, denen morphologische (gestaltliche) und anatomische Merkmale zugrunde liegen. Moleküle (Proteine, DNA) und Morphologie passten zusammen, so wie es ausevolutionstheoretischer Perspektive auch erwartet worden war. Man sah darin eine Bestätigung, die Cladogramme als evolutionäre Abstammungsabfolgen zu interpretieren.
Bald aber tauchten auch Diskrepanzen auf, die Zusatzhypothesen und kompliziertere Erklärungen erforderten, um die Daten evolutionstheoretisch schlüssig interpretieren zu können. Es stellte sich heraus, dass molekulare Daten den klassischen Verwandtschaftsbeziehungen insbesondere der größeren systematischen Gruppen oft widersprachen (s. z. B. Richter & Sudhaus 2004; Valentine 2004, 115, 148; u. v. a.). Losos et al. (2012) berichten nun von einem Fall einer schwerwiegenden Diskrepanz zwischen morphologischen und molekularen Phylogenien, für deren Auflösung nach ihrer Einschätzung keinen Schüssel in Sicht ist. Es geht dabei um die Ähnlichkeitsbeziehungen der Eidechsenartigen (Lacertilia) mit über 30 Familien und ca. 9000 Arten. Sie werden in die Leguanartigen (Iguania), Geckoartigen (Gekkota), Skinkartigen (Scincomorpha) und die Schleichenartigen (Anguimorpha) unterteilt. Nach traditioneller Sicht gabeln sich die Eidechsenartigen an der Basis in die Leguanartigen und die Scleroglossa, diese wiederum in die Geckoartigen und alle anderen Gruppen (siehe Abb. unter http://www.sciencemag.org/content/338/6113/1428/F1.large.jpg). Die Leguanartigen besitzen demnach viele anzestrale („ursprüngliche“) Merkmale. Doch widersprechen molekulare Daten, die in den letzten zehn Jahren gewonnen wurden, dieser Sicht massiv. Demnach befinden sich die Leguanartigen nämlich weit oben im Cladogramm bei den Schleichartigen und Schlangen. Dem auf morphologischen Merkmalen basierten Cladogramm liegt eine enorm große Datenbasis von 192 Arten und 610 variablen Merkmalen zugrunde, von denen 40% erst durch neue Untersuchungstechniken ermittelt wurden. Es war erwartet worden, dass die Verbesserung des morphologischen Datensatzes zu einer Annäherung an das molekular begründete Cladogramm führen würde, „doch die Daten hätten nicht widersprüchlicher sein können“ (Losos et al. 2012, 1429). Kein einziges relevantes Merkmal unterstützt die Position der Leguanartigen hoch im Cladogramm der Eidechsenartigen.
{kind=link}
Welche Lösungsmöglichkeiten gibt es? Losos et al. (2012) stellen fest, dass bei widersprüchlichen Phylogenien einer der beiden Datensätze irreführend sein müsse. Zum einen könnten viele gemeinsame morphologische Merkmale konvergent oder durch Rückentwicklung entstanden sein. Dann würden die Ähnlichkeiten fälschlicherweise gemeinsame Abstammung vortäuschen. Das müsse man häufig annehmen. Doch in diesem Fall sei dies unwahrscheinlich, weil die anzunehmenden Rückentwicklungen viele funktionell verschiedene Teile der Anatomie betreffen, die zudem embryonal auf verschiedene Weisen entstehen. Außerdem haben die Leguanartigen verschiedene Lebensweisen, von großen Pflanzenfressern über ameisenfressende Krötenechsen bis zu gleitend sich fortbewegenden Formen. Diese Vielfalt spiegle kaum eine Anpassung an einen allgemeinen Lebensstil vonwider, so Losos et al.
Aber die Alternative, dass die molekularen Daten irreführend sind, scheitert an der Tatsache, dass nicht weniger als 44 protein-codierende Gene dem molekularen Cladogramm zugrundliegen, die alle in dieselbe Richtung weisen. Es sind zwar Fälle molekularer Konvergenzen bekannt, aber hier müsste ein unerklärbar hohes Ausmaß an Konvergenzen angenommen werden. Wir sind mit einem Rätsel konfrontiert, konstatieren Losos et al.1 Wenn Konvergenzen derart tiefgreifend sind, welches Vertrauen könne man dann in Phylogenien fossiler Taxa haben, für die keine molekularen Daten verfügbar sind? Und akzeptiert man die morphologisch basierte Phylogenie, fehlt eine Erklärung für die molekularen Befunde.
Sollte dieser durchausgesprochen zahlreiche Daten gestützte Befund ein Hinweis darauf sein, dass das evolutionäre Paradigma sich nicht bewährt? Jedenfalls entsteht das bisher unlösbare Rätsel nur deshalb, weil Evolution als Interpretationsrahmen nicht zur Disposition gestellt wird.
Literatur
Losos JB, Hillis DM & Greene HW (2012) Who speaks with a forked tongue? Science 338, 1428-1429.
Richter S & Sudhaus W (Hg, 2004) Kontroversen in der Phylogenetischen Systematik der Metazoa. Sitzungsbericht der Gesellschaft Naturforschen der Freunde zu Berlin. N.F. 43, 1-221. Buchbesprechung: https://www.wort-und-wissen.org/rezension/kontroversen-in-der-phylogenetischen-systematik/.
Valentine JW (2004) On the origin of phyla. University of Chicago Press.
Anmerkung
1 Eine mögliche (wenn auch unbefriedigende) Erklärung, die die Autoren selbst anführen, liegt in der Struktur des Baumes der molekularen Analyse (Abb. B unter http://www.sciencemag.org/content/338/6113/1428/F1.large.jpg), bei dem es sich um ein Phylogramm handelt, das im Gegensatz zum auf der Morphologie basierenden Cladogramm (Abb. A) nicht nur die Topologie, sondern auch die Astlängen zeigt. Die Linie zu den Leguanen in Abb. 1B ist sehr stark in die Länge gezogen im Vergleich zu allen anderen Ästen des Baumes. Bei solchen Mustern, die man auf unterschiedliche Evolutionsraten der Gene in verschiedenen Organismenlinien zurückführt, werden die Methoden der phylogenetischen Analyse leicht überstrapaziert oder deren Vorannahmen verletzt. Dies könnte bedeuten, dass die unerwartete Gruppierung der Leguane ein Artefakt der Analyse darstellt und dass daher diese molekularen Daten gar nicht in der Lage sind, die Beziehungen zu klären (immer unter der Annahme einer gemeinsamen Abstammung der Reptilien). Plausible Gründe für solche stark beschleunigten Raten sind zumindest in diesem Fall offenbar nicht bekannt. Der molekulare Baum sagt daher im Wesentlichen aus, dass Leguane von allen anderen Reptilien genetisch sehr stark abweichend sind.
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n198.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.01.13 Detaillierte Ähnlichkeiten der Gehörorgane von Heuschrecke und Mensch
Das Gehörorgan einer Laubheuschrecke weist einen ganzen Komplex von Ähnlichkeiten im Bauplan und in der Funktionsweise mit dem Gehörorgan des Menschen auf. Die Wissenschaftler, die das mit Hilfe von Mikro-Tomographen herausfanden, sprechen – evolutionstheoretisch orientiert – von einer „Serie konvergenter Lösungen“. Das heißt: Ohne Zielorientierung (die es im Rahmen evolutionstheoretischer Modellierungen nicht gibt) soll zweimal unabhängig eine bis in die Details sehr ähnliche komplexe Struktur und Funktionsweise bei stammesgeschichtlich weit voneinander entfernten Tiergruppen erreicht worden sein – eine außerordentliche Konvergenz.
Eine höchst erstaunliche Entdeckung hat ein Forscherteam um Fernando Montealegre-Zapata, Insektenforscher an der Universität von Bristol in Großbritannien gemacht. Die Wissenschaftler fanden mit Hilfe von Mikro-Tomographen heraus, dass das Gehörorgan der im Regenwald lebenden Laubheuschrecke Copiphora gorgonensis aus der Gruppe der Katydiden ganz ähnlich funktioniert wie das menschliche, und zwar sowohl in anatomischer als auch in physiologischer Hinsicht. Das an den Beinen befindliche Insektenohr ist wie beim Menschen funktionell dreigeteilt, im Vergleich dazu jedoch extrem miniaturisiert, nämlich etwa 60 mal kleiner (das Insektenohr ist insgesamt nur 0,6 mm groß). Legt man eine evolutionäre Entstehungsgeschichte des Lebens zugrunde, handelt es sich hier um ein Beispiel für eine extrem detailgetreue Konvergenz, dessen Tragweite für die Frage nach dem „Wie“ der Entstehung kaum überschätzt werden kann.
Um die weitreichenden Ähnlichkeiten der beiden Sinnesorgane zu verstehen, soll zunächst der Aufbau des menschlichen Ohres beschrieben werden. Außen-, Mittel- und Innenohr ermöglichen Schallaufnahme, Schallumwandlung und Frequenzanalyse. Die Schallwellen erreichen über den äußeren Gehörgang das Trommelfell, das in Schwingung versetzt wird. Daran schließen sich im Mittelohr die bekannten drei Gehörknöchelchen Hammer, Ambos und Steigbügel an. Diese kleinsten Knochen unseres Skeletts leiten die durch die Schallwellen ausgelösten Schwingungen des Trommelfells zum sehr viel kleineren ovalen Fenster weiter. Von dort werden die Schwingungen durch an die mit Flüssigkeit (Perilymphe) gefüllten Gänge der Schnecke übertragen. Die Gehörknöchelchen haben zusammen mit dem Trommelfell die Funktion eines Impedanzwandlers, d. h. sie ermöglichen die Umwandlung niedriger Schalldrücke und hoher Auslenkungen der Luft in hohe Drücke und geringe Auslenkungen in der Perilymphe des Innenohrs. Diese Verstärkung des Schalldrucks wäre nicht möglich, wenn die Flüssigkeit des Innenohres direkt an das Trommelfell angrenzen würde.1
In der Schnecke stecken winzige Härchen mit Sinneszellen, die durch die erzeugten Wanderwellen in der Flüssigkeit verbogen werden. Dadurch werden in den Sinneszellen des Innenohrs elektrische Signale erzeugt, was zur Erregung des Hörnervs führt. Auf diese Weise wird letztlich der ankommende Schall in Nervenimpulse umgesetzt und über den Hörnerv sowie die zentralen Hörbahnen ans Gehirn weitergeleitet. In der Schnecke sind die Sinneshaarzellen linear der Länge nach auf einer Membran angeordnet. Jede Zelle reagiert entsprechend ihrer Lage auf der Membran auf eine bestimmte Frequenz, wobei die niedrigsten Frequenzen am einen und die höchsten am anderen Ende erfasst werden – wie bei einem Klavier (tonotopische Repräsentanz).
Montealegre-Zapata et al. (2012) berichten nun, dass die anatomischen und funktionellen Aspekte des Hörvorgangs beim Menschen außerordentlich ähnlich den Verhältnissen beim Gehörorgan einer Heuschrecke des Regenwaldes sind. Sie zeigten, dass bei diesen Insekten trommelfellartige Membranen des Außenohres mit einer steifen, hebelartigen Struktur, der Tympanalplatte verbunden sind. Diese ist ihrerseits mit einer verlängerten flüssigkeitsgefüllten Kammer des Innenohrs verbunden und entspricht damit den Gehörknöchelchen der Säugetiere. Die Impedanzverstärkung ist ähnlich stark wie bei den Säugetieren. Und im Innenohr der Heuschrecke zeigt sich wie bei Säugetieren eine lineare Abfolge von Sinnesrezeptoren, die von Flüssigkeit umgeben sind, ähnlich der Situation in der Schnecke des Säugerohrs. Besonders bedeutsam ist die Entdeckung der (oben beschriebenen) Impedanzwandlung und -verstärkung durch das den Gehörknöchelchen analoge Element, welches mit den Trommelfellmembranen zusammenwirkt (vgl. Hoy 2012). „Insgesamt zeigen die Daten, dass die Impedanzwandlung, die Ausbreitung der Schallwellen und die tonotopische Repräsentanz biophysikalisch analog denselben Eigenschaften des Säugetierohrs sind“2 (Montealegre-Zapata et al. 2012, 971).
Ein Komplex von Konvergenzen. Angesichts dieser detaillierten Ähnlichkeiten sprechen Montealegre-Zapata et al. (2012) von einer „Serie konvergenter Lösungen“. (Zur „Konvergenz“-Problematik siehe auch Artikel Ähnlichkeiten in der Morphologie und Anatomie, https://www.genesisnet.info/index.php?Artikel=41301&Sprache=de&l=1.) Es handelt sich nicht um eine einfache Konvergenz eines einzelnen Bauplanelements, sondern um einen Komplex von Konvergenzen. Evolutionstheoretisch werden Konvergenzen gewöhnlich mit gemeinsamen Selektionsdrücken erklärt, doch sind Selektionsdrücke nur notwendige, aber bei weitem keine hinreichenden Voraussetzungen für Evolution. Als alternative Erklärung werden Entwicklungszwänge genannt. Hoy (2012, 895) bemerkt in seinem Kommentar jedoch, dass damit nur im Falle des Säugerohrs argumentiert werden könne, nicht aber in Bezug auf die Konvergenz zum Insektenohr.3 Im Übrigen können aber auch Konstruktionszwänge nicht für die Entstehung von evolutionär Neuem verantwortlich gemacht werden, sondern bilden ebenfalls nur zu berücksichtigende Randbedingungen. Letztlich genügen weder externe (Selektion) noch interne (Konstruktionszwänge) Rahmenbedingungen als Triebfedern für die Entstehung der Gehörorgane. Um die Entstehung der frappierend ähnlichen Baupläne und der analogen Funktionsweise des Insekten- und Säugerohrs zu verstehen, reichen die bisher bekannten natürlichen Mechanismen offenkundig nicht aus. Darauf weist indirekt auch Hoys (2012, 895) Bemerkung am Schluss seines Kommentars hin, dass die miniaturisierten Ohren der Insekten Vorbilder sein könnten für die Entwicklung der nächsten Generation akustischer Biosensoren.4
Weshalb ist Konvergenz in diesem Fall so schwerwiegend? Es ist schon ein evolutionstheoretisches Problem ersten Ranges, wie Sinnesorgane überhaupt in einzelnen Abstammungslinien entstehen. Man muss sich die Details klarmachen, die zusammenstimmen müssen, damit ein Ohr überhaupt eine minimale Funktion aufweisen kann. Und man muss sich vor Augen halten, dass es in einer natürlich verlaufenden Evolution keine Zielorientierung gibt. Wenn nun aber ohne Zielorientierung zweimal unabhängig eine bis in die Details sehr ähnliche komplexe Struktur und Funktionsweise bei stammesgeschichtlich weit voneinander entfernten Tiergruppen erreicht wird, muss das zu denken geben.5 Bisher bekannte natürliche Evolutionsmechanismen sind für die Erklärung dieses Phänomens ungeeignet, jedoch in einem Schöpfungskontext leicht zu verstehen.
Literatur
Hoy RR (2012) Convergent evolution of hearing. Science 338, 894-895.
Montealegre-Z. F, Jonsson T, Robson-Brown KA, Postles M & Robert D (2012) Convergent Evolution Between Insect and Mammalian Audition. Science 338, 968-971.
Anmerkungen und Originalzitate
1 „Nahezu die gesamte Schallleistung, die in den Gehörgang dringt, wird an das Innenohr weitergegeben. Hierbei erhöht sich die ausgeübte Kraft vom Trommelfell bis zum ovalen Fenster etwa um den Faktor 90 und der Druck etwa um den Faktor 22. Das heißt: Wären Trommelfell und ovales Fenster starr verbunden, wäre die Schallübertragung um fast 30 Dezibel schlechter, leise Geräusche wären nicht mehr wahrnehmbar.“ (http://tinyurl.com/ada6dxv)
2 „Altogether, the data show that the impedance conversion, dispersive wave propagation, and tonotopic representation are biophysically analogous to the same qualities of the mammalian cochlea.“
3 „The three compartment hearing organ in terrestrial vertebrates, including mammals, evolved out of the developmental imperatives surrounding the evolution of the vertebrate ear on the head. However, no such constraints apply to the hearing organs of insects“ (Hoy 2012, 895).
4 „The miniature ears of insects may provide valuable insights for developing the next-generation of auditory biosensors“ (Hoy 2012, 895).
5 Man kann das Problem vielleicht andeutungsweise damit vergleichen, dass zwei Wanderer blind von ganz verschiedenen Ausgangspunkten aus ohne Wissen eines Zieles und ohne Lenkung denselben weit von ihnen entfernten Punkt erreichen sollten.
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n191.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.11.12 Rätselhafter Ursprung des Linsenauges der Wirbeltiere
Vergleichende genetische Studien am sehr einfach gebauten Frontalauge des Lanzettfischchens erweisen eine Reihe von Gemeinsamkeiten mit Linsenaugen der Wirbeltiere. Die Augentypen sind aber morphologisch* weit voneinander entfernt. Das Linsenauge der Wirbeltiere erscheint im Fossilbericht in fertiger Form.
*morphologisch (= den Aufbau betreffend)
Die evolutionäre Entstehung des Auges gilt in populären Darstellungen häufig als im Wesentlichen geklärt. Meist wird auf eine Reihe verschiedener Augentypen verwiesen (Flachauge – Grubenauge – Lochkameraauge – verschiedene Typen von Linsenaugen), um die schrittweise Entstehung beispielsweise unseres Linsenauges als plausibel erscheinen zu lassen. Wie schon an anderer Stelle dargestellt, sind diese Reihen aus mindestens zwei Gründen bei weitem nicht beweiskräftig für Evolution: 1. Die Unterschiede zwischen den einzelnen (mutmaßlichen) Stufen sind teilweise so enorm, dass die Übergänge völlig neue Konzepte erfordern würden. 2. Diese Serien sind nur modellhaft und nicht gleichbedeutend mit phylogenetischen Abfolgen (vgl. Ullrich et al. 2006 http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html).
Im Fossilbericht tauchen zudem die Komplexaugentypen bereits sehr früh auf (vgl. „Moderne Optik“ im frühen Kambrium, https://www.genesisnet.info/index.php?News=173), aber auch die Linsenaugen sind – gemessen am Fossilbericht – von Anfang an fertig. Daher beginnen die Autoren einer aktuellen Studie ihren Artikel über das „Frontalauge“ (Pigmentbecherauge) des Lanzettfischchens (Amphioxus) mit der Feststellung, dass der Ursprung der Wirbeltieraugen immer noch rätselhaft sei (Vopalensky et al. 2012). Das klingt überraschend, wenn man sich die eingangs genannten Behauptungen vor Augen hält. Weiter stellen die Autoren fest, dass ein Teil des Problems im Mangel heutiger Tierstämme mit brauchbaren Abstufungen liege, die ein Szenario zur Entstehung des Wirbeltierauges ermöglichen könnten. So sei beispielsweise das Auge ausgewachsener Neunaugen (Petromyzontidae, die als ursprüngliche Vertreter der Wirbeltiere gelten) den Augen von kiefertragenden Wirbeltieren im Gesamtaufbau, den Zelltypen der Netzhaut und in vielen Klassen der Opsine (Proteinanteil eines Sehpigments) bemerkenswert ähnlich. Eine Serie von Augentypen zunehmender Komplexität zum Linsenauge der Wirbeltiere hin ist also bloße Hypothese.

Abb. 1: Lanzettfischchen Branchiostoma lanceolatum. (Wikimedia: © Hans Hillewaert, CC BY-SA 4.0)
Eine Hypothese zur Entstehung der Linsenaugen der Wirbeltiere besagt, dass das Pigmentbecherauge am vorderen Ende von Amphioxus ein Vorläufer dafür gewesen sein könnte. Allerdings ist es vergleichsweise sehr einfach gebaut und besteht aus einfachen bewimperten Zellen, unähnlich den Zapfen und Stäbchen der Wirbeltieraugen, die ausgefeiltere Cilien über der Oberfläche besitzen. Der einzige Hinweis darauf, dass die Becheraugen lichtempfindlich sind, bestand bisher darin, dass die bewimperten mutmaßlichen Sinneszellen in unmittelbarer Nähe zu dunklen Pigmentzellen angeordnet sind (Vopalensky et al. 2012).
In ihrer Studie verglichen Vopalensky et al. (2012) das Pigmentbecherauge von Amphioxus in Bezug auf Gemeinsamkeiten mit Linsenaugen von Wirbeltieren in molekularer Hinsicht und fanden viele Gemeinsamkeiten bei Transkriptionsfaktoren*, Opsinen (s. o.) und Proteinen der Phototransduktionskaskade (Umwandlung eines äußeren Lichtreizes in ein physiologisches Signal). Außerdem fanden sie molekulare Gemeinsamkeiten zwischen den Pigmentzellen von Amphioxus und dem retinalen Pigment-Epithel1 der Wirbeltieraugen (Melanin-Gehalt, Regulationsgene). Und schließlich ähneln Nervenfortsätze des Frontalauges Strukturen im Vorderhirn der Wirbeltiere.
*Transkriptionsfaktoren (= Proteine, die bei der Regulation wichtig sind)
Damit können die Autoren eine (vor allem molekular begründete) Homologie zwischen dem Frontalauge von Amphioxus und dem Wirbeltier-Linsenauge begründen. Ein stammesgeschichtlicher Zusammenhang kann daraus jedoch nicht abgeleitet werden; die verglichenen Strukturen sind, trotz der erstaunlichen Ähnlichkeiten auf biochemischer bzw. molekularbiologischen Ebene, morphologisch weit voneinander entfernt. Zur Klärung der Frage nach den Mechanismen der stufenweisen Entstehung und Abwandlung des Wirbeltierauges sind bloße Vergleiche überfordert (das war allerdings auch nicht das Ziel der Untersuchungen). Die Studie von Vopalensky et al. steuert jedoch ein weiteres interessantes Beispiel dafür bei, dass der ontogenetischen Entwicklung morphologisch verschiedener Organe unterschiedlicher Tierstämme oder -klassen auf der Basis ähnlicher molekularer Grundlagen erfolgt – ein Befund, der bei vielen anderen Organsystemen verschiedener Tierstämme bekannt ist.
Aus den Befunden kann geschlossen werden, dass bereits im hypothetischen Ausgangsindividuum die molekularen Grundlagen des Sehens in Form funktionierender hochkomplexer biochemischer Kaskaden existiert haben müssen. Wie diese entstanden sind, liegt ebenso weiterhin im Dunkeln wie der Ursprung der Sehorgane, die sich ihrer zur Lichtwahrnehmung bedienen.
Anmerkung
1 Zum retinalen Pigment-Epithel vgl. die Ausführungen in Ullrich et al. (2006) (http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html)
Literatur
Vopalensky P, Pergner J, Liegertova M, Benito-Gutierrez E, Arendt D & Kozmik Z (2012) Molecular analysis of the amphioxus frontal eye unravels the evolutionary origin of the retina and pigment cells of the vertebrate eye. Proc. Natl. Acad. Sci. USA, early ed., doi: 10.1073/pnas.1207580109.
Ullrich H, Winkler N & Junker R (2006) Zankapfel Auge. Stud. Int. J. 13, 3-14, http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html.
Autor dieser News: Reinhard Junker
© 2012, http://www.genesisnet.info/schoepfung_evolution/n188.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
11.04.12 Bestätigung einer wichtigen Funktion des Wurmfortsatzes
Der Wurmfortsatz des menschlichen Blinddarms gilt als Paradebeispiel für ein rudimentäres, rückgebildetes, nutzloses Organ. Doch es gibt eine wachsende Anzahl von Hinweisen darauf, dass dies eine Fehlinterpretation ist. Die Hypothese, dass der Wurmfortsatz eine Art „Rettungsstation“ für wichtige Darmbakterien bildet, konnte kürzlich erfolgreich getestet werden.
Der Wurmfortsatz (Appendix) des menschlichen Blinddarms ist vielleicht das bekannteste Beispiel eines rudimentären Organs, das seine frühere Funktion verloren habe und daher oft als nutzloser und sogar gefährlicher Rest einer evolutiven Rückbildung angesehen wird. Solche (vermeintlich) rudimentären Organe werden oft auch als Belege gegen Schöpfung gewertet. Es gibt allerdings schon lange Hinweise auf eine Funktionalität des Wurmfortsatzes und es spricht auch manches dagegen, dass dieser bleistiftgroße Darmanhang überhaupt als rückgebildet gelten kann (zusammenfassend bei Junker 2008). Und vor einigen Jahren fanden Bollinger et al. (2007) heraus, dass der Wurmfortsatz des Menschen eine Art Zufluchtsort und Rettungsstation für symbiotische* Bakterien darstellt, die das Wachstum nützlicher Darmbakterien fördert und bei durchfallbedingten Darmentleerungen die Wiederbesiedlung mit diesen Bakterien ermöglicht bzw. erleichtert. Diesen Bakterien fällt die Aufgabe zu, die Ausbreitung gefährlicher Krankheitserreger im menschlichen Verdauungstrakt zu verhindern, was besonders nach Durchfallerkrankungen sehr wichtig ist. Die Forscher schlussfolgerten, dass der Wurmfortsatz für eine bedeutsame Funktion offensichtlich optimal eingerichtet ist (vgl. Der Wurmfortsatz als Rettungsstation, https://www.genesisnet.info/index.php?News=101).
*symbiotisch (= mit anderen Organismen zusammenlebend)
Die Hypothese vom Wurmfortsatz als Zufluchtsort konnte inzwischen getestet werden. Es wurde untersucht, ob Patienten, die eine schwere Darminfektion hatten, sich besser erholten, wenn sie noch im Besitz des Wurmfortsatzes waren, als solche Patienten, bei denen das nicht der Fall war und die dieselbe Infektion hatten. James Grendell vom Winthrop University-Hospital und Kollegen untersuchten die Folgen von Infektionen mit dem Krankheitserreger Clostridium difficile (Im et al. 2011). C. difficile ist besonders gefährlich, wenn die natürliche Darmflora, die den Erreger normalerweise in Schach hält, zu stark dezimiert ist (z. B. nach einer Antibiotikatherapie). Patienten mit dem Wurmfortsatz sollten daher einen messbar größeren Schutz vor einer Wiederholungsinfektion nach erfolgreicher Ersttherapie einer bakteriellen Darminfektion mit C. difficile besitzen.
Grendells Team konnte 254 geeignete Patienten entsprechend untersuchen. Dabei zeigte sich tatsächlich, dass Personen ohne Wurmfortsatz viermal häufiger von einem Wiederauftreten von C. difficile betroffen waren, genau wie aufgrund der Hypothese vom Wurmfortsatz als Zufluchtsort für schützende Darmbakterien vorhergesagt wurde.
Diese Hypothese ist damit noch nicht endgültig bewiesen, da die festgestellten Unterschiede auch andere Ursachen haben könnten oder gar nicht typisch sind (vgl. Dunn 2012). Zur endgültigen Klärung sind weitere Studien erforderlich.
Literatur
Bollinger RR, Barbas AS, Bush EL, Lin SS & Parker W (2007) Biofilms in the large bowel suggest an apparent function of the human vermiform appendix. J. Theor. Biol. 249, 826-831.
Dunn R (2012) Your Appendix Could Save Your Life. http://blogs.scientificamerican.com/guest-blog/2012/01/02/your-appendix-could-save-your-life.
Im GY, Modayil RJ, Lin CT, Geier SJ Katz DS, Feuerman M & Grendell JH (2011) The appendix may protect against Clostridium difficile recurrence. Clin. Gastroenterol. Hepatol. 9, 1072–1077.
Junker R (2008) Der Wurmfortsatz als Rettungsstation Stud. Int. J. 15, 31-32.
Autor dieser News: Reinhard Junker
© 2012, http://www.genesisnet.info/schoepfung_evolution/n183.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
10.11.11 Superschnelle Muskeln (auch) bei Fledermäusen
Fledermäuse sind in der Lage, ihre Umgebung mit Hilfe von Ultraschall so genau zu scannen, dass man ihre Fähigkeiten am besten mit „Echo-Bildsehen“ beschreibt. Wenn man so will: Fledermäuse „sehen“ mit den Ohren (vgl. Entstehung der Fledermäuse, https://www.genesisnet.info/index.php?Artikel=42846&Sprache=de&l=1). Aber anders als beim Sehen mit den Augen müssen die Fledermäuse auch noch die Daten teilweise selbst generieren, deren Auswertung dann das Echobild liefert.
Um eine geortete Beute schlagen zu können, müssen die Fledermäuse ihre Ultraschall-Rufe bei Annäherung zunehmend häufiger und kürzer ausstoßen, bis sie schließlich kaum vorstellbare 190 Rufe pro Sekunde erreichen (deren Echo sie aufnehmen und auswerten). Bisher erschien den Forschern diese Schlussphase extrem häufiger Rufe, der sogenannte „terminal buzz“, rätselhaft; es war unbekannt, wie die Fledermäuse die Laute in so schneller Abfolge erzeugen können. Biologen von der Universität von Süddänemark in Odense und der Universität von Pennsylvania in Philadelphia kamen den Fledermäusen nun auf die Schliche (Elemans et al. 2011). Sie entdeckten bei Wasserfledermäusen (die knapp über Wasseroberflächen nach Insekten jagen) hochspezialisierte „superschnelle“ Muskeln, deren Tätigkeit die schnellen Laute ermöglicht. Experimente mit isolierten Muskelfaserbündeln des Kehlkopfs von Myotis daubentonii zeigten, dass die Kehlkopfmuskeln der Fledermaus sich bis zu 200-mal pro Sekunde zusammenziehen können. Der Kehlkopfmuskel kann sich damit bis zu zwanzig Mal schneller als die schnellsten Muskeln des Menschen zusammenziehen, die bei der Augenbewegung gebraucht werden.
Außerdem konnten die Forscher zeigen, dass die Leistungsfähigkeit des Kehlkopfmuskels der limitierende Faktor bei der Häufigkeit der Laute ist und nicht die bei Annäherungen an die Beute zunehmende Überlappung von Ruf und Echo.
Die superschnellen Muskeln des Kehlkopfs sind aber auch noch aus einem weiteren Grund interessant. In Spektrumdirekt wird Coren Elemans mit folgenden Aussagen über den superschnellen Muskeltypus zitiert: „Das ist eine ganz andere Muskelart als unsere normalen Skelettmuskeln. … Wir haben etliche Anpassungen auf zellulärer und molekularer Ebene gefunden, zum Beispiel eine stark erhöhte Mitochondrienanzahl und sehr viel mehr sarkoplasmatisches Retikulum, welches Kalzium in und aus der Zelle pumpt.“ Dieser Muskeltypus ist auch bei Klapperschlagen, einigen Fischen und Vögeln bekannt, also bei Arten, die in der Systematik deutlich getrennt stehen. Er müsste also mehrfach unabhängig, konvergent entstanden sein. In allen Fällen spielen die superschnellen Muskeln eine wichtige Rolle bei der akustischen Kommunikation. So zwitschern auch Singvögel mit Hilfe superschneller Stimmmuskeln. Beim Europäischen Star (Sturnus vulgaris) und beim Zebrafinken (Taeniopygia guttata) dauert ein Zyklus von Kontraktion und Entspannung nur 3-4 Millisekunden (Elemans et al. 2008). Mit dem Nachweis von superschnellen Muskeln bei Wasserfledermäusen sind nun diese erstmals auch bei Säugetieren beschrieben worden. Ihre Verteilung bei verschiedenen Tieren erfordert eine mehrfach unabhängige Entstehung, was die Existenz dieser besonderen Muskeln noch erstaunlicher macht.
Literatur
Anonymus (2011) Fledermäuse orten dank superschneller Muskeln. www.wissenschaft-online.de/artikel/1124502.
Elemans CEP, Mead AF, Rome LC & Goller F (2008) Superfast Vocal Muscles Control Song Production in Songbirds. PLoS ONE 3(7): e2581. doi:10.1371/journal.pone.0002581.
Elemans CPH, Mead AF, Jakobsen L & Ratcliffe JM (2011) Superfast Muscles Set Maximum Call Rate in Echolocating Bats. Science 333,1885-1888.
Autor dieser News: Reinhard Junker
© 2011, http://www.genesisnet.info/schoepfung_evolution/n178.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.09.11 Unser Großhirn – „out of the blue“?
Ringelwürmer (Annelida) sind nicht gerade als Intelligenzbestien verschrien. Ihr Gehirn ist relativ einfach aufgebaut, aber gerade deshalb gut geeignet als Modell zur molekularen Erforschung von Gehirnentwicklung und -funktion. Neue Genexpressions*-Analysen am Gehirn von Platynereis dumerilii, einem im Meer lebenden Borstenwurm, haben deutliche Gemeinsamkeiten mit der Großhirnrinde der Wirbeltiere belegt, dem komplexesten Organ in der belebten Welt (Tomer et al. 2010). Die Autoren der Studie schließen daraus: „Die Evolutionsgeschichte unseres Großhirns muss neu geschrieben werden.“
*Genexpression (= Ablesen der Geninformation)
Eine auffällige paarige Struktur im Annelidenhirn, wie auch im Zentralhirn vieler Insekten, ist der sogenannte Pilzkörper. Seine pilzförmige Struktur bildet ein Zentrum mit dicht gepackten Neuronen (Nervenzellen), die tausende bis zu mehreren hunderttausend parallel laufende Nervenfortsätze entsenden. Der Pilzkörper ist vor allem für die Verarbeitung von Geruchsinformationen zuständig, mischt aber auch bei Lern- und Gedächtnisleistungen mit. Aktuell wird z. B. an der Fruchtfliege erforscht, wie Gedächtnis und Lernen molekular im Pilzkörper funktioniert (Davis 2011). Es gibt auch Hinweise, dass der Pilzkörper eine Rolle beim Ausfiltern wichtiger gegenüber weniger wichtigen Umgebungsinformationen spielt (Farris 2011).
Es war schon länger bekannt, dass es strukturelle, anatomische und funktionelle Ähnlichkeiten des Pilzkörpers mit dem Cortex (oder Pallium, Großhirnrinde) der Wirbeltiere gibt, also mit dem Teil unseres Gehirns, das für komplexe Leistungen zuständig ist, wie zum Beispiel dem Ersinnen von Experimenten zur Erforschung des Gehirns. Weil Ringelwürmer und Wirbeltiere stammesgeschichtlich sehr weit entfernt stehen (Protostomia vs. Deuterostomia, Urmünder vs. Neumünder), wurden diese Ähnlichkeiten aber bisher nicht als Homologien (Ähnlichkeit durch Verwandtschaft) gedeutet, sondern als konvergent erworben interpretiert (Ähnlichkeiten, die durch funktionelle Notwendigkeit in nicht näher verwandten Gruppen erzwungen wurden). Nachdem nun aber die oben erwähnten Genexpressions-Experimente neben einigen Unterschieden auch viele Parallelen im Muster aktiver Gene im Pilzkörper zu dem entsprechenden Muster im Wirbeltier-Cortex aufdeckten, schließen die Autoren, dass diese Ähnlichkeiten zu komplex und zahlreich sind, um unabhängig durch funktionelle Notwendigkeit, also konvergent, erworben zu sein. Stattdessen folgern sie, dass der letzte gemeinsame Vorfahr von Ringelwürmern und Wirbeltieren schon die entsprechenden molekularen Gemeinsamkeiten trug, der Vorläufer des Großhirns also schon sehr früh entstanden sei, bevor sich die Linien trennten, die zu den Ringelwürmern bzw. zu den Wirbeltieren führten.
Die Arbeit ist methodisch beeindruckend, die Autoren entwickelten ein Protokoll, das es ermöglicht, Genexpressions-Profile mit zellulärer Auflösung für den gesamten Organismus der Anneliden-Larve durchzuführen. Aufschlussreich ist allerdings auch die Methodik in Bezug auf stammesgeschichtliche Schlussfolgerungen. Im vorliegenden Fall hat sich offensichtlich durch die neuen Genexpressions-Befunde die Anzahl an Ähnlichkeiten von Großhirnrinde zu Pilzkörper gegenüber der Zahl der Unterschiede so erhöht, dass sie einen (keineswegs vordefinierten) „Grenzwert für Plausibilität“ überschritt, so dass Merkmale, die zuvor als vielfache Konvergenzen gedeutet wurden, nun zu Homologien umdefiniert werden. Eigenschaften die bisher als durch funktionelle Notwendigkeit erworben galten, sind nun Folge gemeinsamer Abstammung mit der Konsequenz: die Evolutionsgeschichte des Großhirns muss umgeschrieben werden.
Etwas erstaunlich ist, dass offenbar die gerade erschlossene neue Einstufung dann selbstbewusst von den Autoren als endgültig angesehen wird: Um zu einer möglichen Rekonstruktion des Gehirns des erwarteten letzten gemeinsamen Vorfahren zu gelangen, könne man das Gehirn von Krebstieren oder Mollusken (Protostomia) oder dem Lanzettfischchen (Deuterostomia) untersuchen, so die Autoren im Ausblick ihres Artikels. Bei diesen Organismen sind noch keine deutlichen Pilzkörper-Morphologien entdeckt worden und im angenommenen Stammbaum stehen sie zwischen Ringelwürmern und Wirbeltieren. Sollten Genexpressions-Untersuchungen auf ähnliche Muster wie bei Ringelwürmern stoßen, bestätige dies, dass ihre Gehirne dem angenommen einfacheren Gehirn des letzten gemeinsamen Vorfahren ähneln. Wenn dagegen die Muster nicht gefunden würden, stelle das die neue Gehirn-Systematik allerdings auch nicht infrage: Dann hätten die untersuchten Gruppen vorherige Komplexität eben sekundär wieder verloren …
Wie vor kurzem für Archaeopteryx beschrieben (Fällt eine Ikone vom Sockel?, https://www.genesisnet.info/index.php?News=174) zeigt auch dieses Beispiel, dass Ähnlichkeiten von Merkmalen an sich keine verlässlichen Verwandtschaftsanzeiger sein können. Es kann nicht im Voraus definiert werden, welche Qualität oder Quantität eines ähnlichen Merkmals den Erwerb durch gemeinsame Vorfahren wahrscheinlicher macht als einen unabhängigen, konvergenten Erwerb. Ob Merkmale als Homologien oder Konvergenzen gedeutet werden hängt immer vom vorher erwarteten bzw. bislang konstruierten Stammbaum ab. Dieses Vorgehen führt zu unauflösbaren Zirkelschlüssen. Insbesondere wenn, wie wiederholt berichtet, komplexe Merkmale mosaikartig gleichzeitig bei einfachen und höheren Lebewesen auftauchen, führt dies oft zu neuen, wiederum auch nicht unproblematischen evolutionären Einordnungen. Meist werden sie als Konvergenzen gedeutet, im hier beschriebenen Artikel als Homologien. Das führt dazu, dass Arendt spekuliert: „… we learn that it (der Vorläufer des Palliums) came ‘out of the blue’ – as an adaptation to early marine life in Precambrian oceans“ (http://insciences.org/article.php?article_id=9476).
Im schöpfungstheoretischen Rahmen können entsprechende Mosaik-Befunde schlüssig im Sinne eines der geschaffenen Natur zugrunde liegenden „Baukastensystems“ gedeutet werden, so dass sich spekulative Ausflüge ins Blaue erübrigen.
Literatur
Davis RL (2011) Traces of Drosophila memory. Neuron 70, 8-19.
Farris S (2011) Locusts Provide Clues to Insect Mushroom Body Function. Brain Behav Evol 77, 3-4, doi: 10.1159/000322939.
Jenett et al. (2006) The Virtual Insect Brain protocol: creating and comparing standardized neuroanatomy. BMC Bioinformatics 7, 544, doi: 10.1186/1471-2105-7-544.
Tomer R, Denes A, Tessmar-Raible K & Arendt D (2010) Profiling by Image Registration Reveals Common Origin of Annelid Mushroom Bodies and Vertebrate Pallium. Cell 142, 800-809.
Autor dieser News: Hans-Bertram Braun
© 2011, http://www.genesisnet.info/schoepfung_evolution/n176.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
03.06.11 Gene und soziales Verhalten
Die rasant zunehmenden Daten aus sequenzierten Genomen (#GLOSSAR gesamtes Erbgut einer Art) wurden von Beginn an mit hohen Erwartungen verknüpft. Diese konnten bisher nur in ernüchternd geringem Umfang erfüllt werden. Welche Zusammenhänge gibt es zwischen dem Erbgut und dem Verhalten von Lebewesen? Eine vergleichende Studie an Bienen, die unterschiedlich ausgeprägte Formen sozialen Verhaltens zeigen, dokumentiert Hinweise darauf, dass unterschiedliches Verhalten einhergeht mit markanten Unterschieden im Erbgut. Hier scheint sich eine Möglichkeit zu eröffnen, spannende Fragen zu erforschen.
Mit Eusozialität (gr.: eu = gut; lat.: socialis = kameradschaftlich) bezeichnet man eine Verhaltensweise, einen Lebensstil, bei dem Organismen in Gruppen unterschiedlicher Größe in mehreren Generationen unter Arbeitsteilung kooperativ zusammenleben, beispielsweise durch Staatenbildung.
Verschiedene Insekten sind als soziale, Staaten bildende Arten bekannt, besonders unter Hautflüglern (Hymenoptera): Ameisen, Termiten, Wespen, Bienen. Nacktmulle (Heterocephalus glaber) sind eusoziale Säugetiere.
Das Phänomen des sozialen Verhaltens wird bereits lange und intensiv untersucht (Wilson & Hölldobler 2005), über den Zusammenhang zwischen Molekularbiologie und Sozialverhalten ist bisher aber wenig bekannt. Woodard et al. (2011) haben dazu eine umfangreiche vergleichende Untersuchung an Bienen veröffentlicht. Verschiedene Bienenarten zeigen Sozialverhalten in stark abgestuften Variationen, vom individuellen, so genannten solitären bis zu verschieden ausgeprägten eusozialen Lebensstilen, deshalb sind sie für Forschungen dieser Art vorteilhafte Studienobjekte. Verschiedene Autoren gehen davon aus, dass Bienen mindestens sechsmal unabhängig Eusozialität entwickelt haben (bei Insekten insgesamt soll diese Verhaltensweise mindestens 11-mal unabhängig entstanden sein) – eines der vielen Beispiele für Konvergenz, d. h. Ähnlichkeit, die nicht auf Abstammungsverwandtschaft zurückgeführt wird.
Um die Unterschiede in den Genen im Vergleich mit unterschiedlich ausgeprägten Formen von sozialem Verhalten zu untersuchen, erzeugten Woodard et al. jeweils Kopien derjenigen Bereiche des Bienengenoms, der aktiv genutzt werden (Transkriptom) von 9 verschiedenen Bienenarten. Für die neun Bienenarten wurden DNA-Moleküle im Umfang von 1 Milliarde Basenpaare (~ 1 Gbp) sequenziert. Um die jeweiligen Sequenzen im Bienengenom zuordnen zu können wurden sie mit dem Genom der ausgeprägt eusozialen Honigbiene (Apis mellifera) kombiniert, so dass 10 Gen-Sätze von je 3647 Genen (ca. 33 % der Bienengene) identifiziert werden konnten. Davon wurden 3638 Gene in der vergleichenden Studie genutzt. Nun wurde danach gesucht, in welchen Bereichen der DNA Hinweise auf verstärkte Änderungen für Aminosäureeinbau der codierten Proteine vorliegen. Diese Resultate wurden dann bei Bienenarten mit unterschiedlich ausgeprägter Sozialität miteinander verglichen.
Phylogenetische Analysen der molekularbiologischen Daten ergab für die 10 untersuchten Bienenarten, dass dreimal unabhängig Eusozialität entstanden sein sollte. Für die Untersuchung wurden drei abgestufte Formen für eusoziales Verhalten definiert:
ausgeprägt eusozial (highly eusocial): große Kolonien (1 000 bis 10 000 Individuen), mehrjähriger Kolonie-Lebenszyklus, hoch spezialisierte Königin und Arbeiterinnen, hohe Lebenserwartung für die Königin (10-fache Lebenserwartung einer Arbeiterin), Einfluss der Nahrung auf die Entwicklung, ganzjährige thermoregulierte Bedingungen im Nest, verschiedene exokrine1 Drüsen.
wenig eusozial (primitively eusocial): kleine Kolonien (10 bis 100 Individuen), Jahreszyklus in der Kolonie, wenig spezialisierte Königin und Arbeiterinnen, Einfluss der Nahrung auf die Entwicklung, dominante Hierarchien, verschiedene exokrine Drüsen.
nicht eusozial (non eusocial): Einzelgänger oder Individuen teilen sich ein Nest, 1-jähriger Lebenszyklus, alle Individuen sind reproduktiv aktiv, weibliche Bienen kümmern sich nur um den eigenen Nachwuchs.
Die Untersuchungen für eine Korrelation zwischen veränderten Genen bzw. modifizierte Proteine und der Sozialität ergaben, dass sich 212 von 3638 Genen (~ 6 %) in allen eusozialen Arten deutlich von denjenigen in den nicht eusozialen unterscheiden. 173 (~ 5 %) Gene sind in ausgeprägt eusozialen Bienen und 218 (~ 6 %) in wenig eusozialen im Vergleich zu den jeweils anderen Bienen deutlich verändert.
Am häufigsten waren von den Veränderungen Gene betroffen, die an der Signalübertragung beteiligt sind oder auch an der Drüsenentwicklung und dem Kohlenhydratstoffwechsel, diese zeigen in eusozialen Bienenarten die deutlichsten Unterschiede.
Die Ergebnisse zeigen, dass konvergente2 Entwicklungen zu mosaikartigen Mustern an molekularen Veränderungen führen können, und zwar sowohl bei Genen, die allen untersuchten Bienenarten gemeinsam sind, als auch bei spezifischen. So zeigten beispielsweise Gene, die im Zusammenhang mit der Reproduktion stehen, in wenig sozialen Bienen (Exoneura robusta und Bombus impatiens und B. terrestris [Hummeln]) starke Veränderungen im Vergleich zu den entsprechenden Genen in Vertretern von ausgeprägt oder nicht eusozialen Bienen, wobei sich die Gene der Hummeln und der Biene stark voneinander unterscheiden. In der untersuchten Bienengruppe tritt Eusozialität also insofern konvergent auf, als die Verteilung der eusozialen Verhaltens nicht der phylogenetischen Verwandtschaft der Bienen entspricht. Bei den eusozialen Bienen sind auffällig ähnliche Gruppen von Genen – wie mosaikartige Muster – modifiziert.
Die Untersuchungen zeigen also eine Korrelation von (veränderten) Verhaltensweisen und der Nutzung von Genen bzw. deren Veränderung. Die Autoren interpretieren das als Einfluss des Verhaltens auf die genetische Evolution.
Woodard et al. stellen zum Abschluss ihrer Arbeit eine Reihe von Spekulationen im Zusammenhang mit den von ihnen gefunden Daten an. So halten sie es nicht für überraschend, dass Gene, die im Zusammenhang mit der Entwicklung von Drüsen stehen, starkem Selektionsdruck ausgesetzt sind, da die Drüsenfunktion für die chemische Kommunikation in einer Kolonie von großer Bedeutung ist. Ebenso ist die Signalübertragung ein bedeutsames Merkmal bei der Entwicklung von sozialem Verhalten (Wahrnehmung und Interpretation von Verhaltenweisen anderer Individuen die zur Kolonie gehören). Die kooperative Lebensweise in Kolonien geht auch einher mit einer größeren Abhängigkeit von hochwertigem Honig. Daher ist nachvollziehbar, dass Gene im Zusammenhang mit dem Zuckerstoffwechsel sich in diesem Zusammenhang stark ändern.
Bei wenig sozialen Bienen werden Gene, die mit dem Gehirn im Zusammenhang stehen, auffällig verstärkt exprimiert. Vielleicht, weil bei ihnen die Verhaltensmuster noch nicht so festgelegt und damit variabel sind und das Zusammenspiel zwischen Kooperation und Konkurrenz sehr flexibel ist. Damit wird nachvollziehbar, dass für die Wahrnehmung und Verarbeitung der entsprechenden Signale vergleichsweise viel investiert werden muss.
Das Untersuchungssystem, das Woodard et al. in der hier vorgestellten Arbeit etabliert haben im Zusammenhang mit der Fragestellung des Zusammenhangs zwischen Verhalten und genetischen Veränderungen, erscheint spannend und weckt Hoffnungen und Erwartungen für ein besseres Verständnis der dabei ablaufenden Prozesse. Toth & Robinson (2009) hatten vorgeschlagen, dass für die Entwicklung von Eusozialität ein Satz Werkzeuge (tool-kit) an hoch konservierten Genen verwendet wird, die je nach Bedarf benutzt werden (Evo-Devo3). Woodard et al. sehen in ihren Resultaten einen Hinweis darauf, dass die Gene für den Kohlenhydratstoffwechsel Schlüsselbausteine im vermuteten Werkzeugsatz darstellen.
Mit den von Woodard et al. genutzten Studienobjekten und dem vorgestellten Konzept lassen sich Fragestellungen über den Zusammenhang zwischen Entwicklung von Verhalten und den damit im Zusammenhang stehenden molekularbiologischen Änderungen grundlegend studieren. Die Frage, ob ein Werkzeugsatz vorliegt, auf den bei Bedarf zugegriffen werden kann, oder ob sich soziale Verhaltensweisen sich auf andere Weise entwickeln, bleibt vorerst offen.
Der Gedanke von einem Satz von Werkzeugen provoziert Fragen nach dessen Ursprung und Herkunft. Evolutionäre Modelle konnten bisher keine plausiblen Antworten dazu beisteuern. Das ist u. a. auch in den Ansätzen von „Evo-Devo“-Vertretern dokumentiert. Molekularbiologische Untersuchungen wie die hier vorgestellte Arbeit bieten noch keine Antworten, sondern machen überhaupt erst die Problemstellung deutlicher erkennbar. Unter naturwissenschaftlichen Gesichtspunkten sind Mechanismen zu fordern, die die Zusammenhänge zwischen molekularbiologischen Modifikationen und Änderungen im Phänotyp bzw. Verhalten (allgemein: empirisch feststellbare Äußerungen von Lebewesen) plausibel erklärt.
Literatur
Junker R (2008) Evo-Devo Schlüssel für Makroevolution? Teil 1: Ausgangspunkt und Anerkennung eines ungelösten Evolutionsproblems.
Todt AL & Robinson GE (2009) Evo-devo and the evolution of social behavior: Brain gene expression analysis in social insects. Cold Spring Harb. Symp. Quant. Biol. 74, 419-426.
Wilson EO & Hölldobler B (2005) Eusociality: Origin and consequences. Proc. Nat. Acad. Sci. USA 102, 13367-13371.
Woodard SH, Fischman BJ, Venkat A, Hudson ME, Cameron SA, Clark AG & Robinson GE (2011) Genes involved in convergent evolution of eusociality in bees. Proc. Nat. Acad. Sci. USA doi/10.1073/pnas.1103457108
Anmerkungen
1 Bei exokrinen Drüsen wird das Sekret nicht in den Blutkreislauf (endokrin) sondern in einen Körperhohlraum oder an die Körperoberfläche abgegeben.
2 Konvergenz: ähnliche Anlagen werden bei Organismen entwickelt, die nicht näher mit einander verwandt sind.
3 Zu Evo-Devo s. Junker (2008).
Autor dieser News: Harald Binder
© 2011, http://www.genesisnet.info/schoepfung_evolution/n169.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
20.05.11 Buckelzirpen – wie entstehen spektakuläre Körperanhänge?
Zikaden sind Insekten, deren Mundwerkzeuge zum Anstechen und Aufsaugen von Pflanzensäften geeignet ist. Eine Gruppe von Zikaden bildet aus dem Halsschild z. T. skurrile Strukturen, die über den Rücken verlängert sind. Wie werden diese Strukturen gebildet? Wozu dienen sie? Warum gibt es das nur bei Buckelzirpen? Viele dieser Fragen sind bisher ohne befriedigende Antwort. Zur ersten Frage haben Wissenschaftler jetzt erste Teilantworten gefunden. Ihre Evolution bleibt hypothetisch, auch wenn in der Presse ein anderer Eindruck erweckt wird.
Für die taxonomische Einteilung von Insekten in Einheiten höherer Ordnung orientieren sich die Entomologen häufig am Verlust oder an der Reduktion von Körperanhängen (wie z. B. Beinen oder Flügeln). Dabei liegen den Bezeichnungen Verlust oder Reduktion vergleichende Betrachtungen zugrunde, die evolutionstheoretisch geprägt sein können. Neue, zusätzliche Körpermerkmale werden sehr selten angenommen, und die Bedingungen, unter denen sie entstehen, sind noch weitgehend unverstanden.
Ein Team von Wissenschaftlern unter der Leitung von Nicolas Gompel hat nun eine Untersuchung veröffentlicht (Prud’homme et al. 2011), in der sie die Entstehung der teilweise bizarren Halsschilde1 (Pronotum, engl. helmet; manche haben die Form eines Helms) von Buckelzirpen studieren. Buckelzirpen (Membracidae) sind eine Insektenfamilie aus der Ordnung der Schnabelkerfe (Hemiptera) und der Unterordnung der Rundkopfzikaden (Cicadomorpha); sie stechen Pflanzen an und ernähren sich von den zuckerhaltigen Pflanzensäften (Phloem-Sauger).
Der Halsschild wird am ersten Brustsegment gebildet und von Prud’homme et al. als homolog2 zu den dort prinzipiell angelegten Flügeln interpretiert. Diese Strukturen sind einzigartig in der Geschichte der Insekten und charakteristisch für alle Buckelzirpen. Bisherige Bearbeiter haben die Körperanhänge ganz unterschiedlich interpretiert und es gab darüber kontroverse Diskussionen. Von einfachen Auswüchsen unterscheiden sich die Halsschilde als Körperanhänge dadurch, dass sie beweglich mit dem Körper verknüpft sind, so wie die Flügel am zweiten und dritten Brustsegment. Darin sind die Halsschilde von Auswüchsen bei anderen Insekten, z. B. den „Horn“-artigen Strukturen bei Käfern unterschieden.
Gompel und sein Team gingen nun der Frage nach, wie solche Körperanhänge auf dem Rücken am ersten Brustsegment im Laufe der Evolution entstanden sein könnten: entweder komplett neu (de novo) oder unter Verwendung und Modifikation eines bestehenden Entwicklungsprogramms?
Dazu untersuchten sie verschiedene Larvenstadien von Buckelzirpen. Auf den ersten Blick erscheinen die Halsschilde nicht als paarig angelegte Struktur (wie z. B. Flügel), sondern als eine einheitliche Struktur, die über dem Rücken verlängert ist. Durch Untersuchungen am ersten Larvenstadium von Buckelzirpen konnten die Autoren jedoch zeigen, dass die Körperanhänge zunächst doch paarig angelegt und später auf dem Rücken fusioniert werden. Als Körperanhänge der Brustsegmente (T2 und T3), die zum Rücken hin ausgebildet werden, sind bei Insekten nur Flügel bekannt. Vergleichende Untersuchungen von Halsschild und Flügel in verschiedenen Larvenstadien zeigen, dass das Drehgelenk und die Aufhängung des Halsschilds dem der Flügel sehr ähnlich ist, und beide Strukturen sind durch ein Netzwerk von Adern charakterisiert. Aufgrund dieser Beobachtungen interpretieren Prud’homme et al. beide Strukturen als homolog. Wenn Halsschild und Flügel homolog sind, dann müssten nach der Erwartung der Autoren beide Strukturen während Entwicklung durch dieselben genetischen Programme gesteuert werden. Die Aktivität eines Transkriptionsfaktors (Nubbin) und zweier weiterer Gene (Distal-less [Dll] und homothorax [hth]) lassen sich tatsächlich sowohl in den Flügelanlagen als auch im sich entwickelnden Halsschild nachweisen. Die Entwicklung beider Anlagen unterliegt also mindestens teilweise derselben genetischen Steuerung.
Alle heute bekannten geflügelten Insekten bilden ausschließlich an den Brustsegmenten 2 und 3 (T2 und T3) entsprechende Anhänge, aber nicht an T1. Es ist bekannt, dass das genetische Programm für die Anlage von Flügeln unterdrückt werden kann; es wird schon länger vermutet, dass das beim 1. Brustsegment der Fall ist. Im Fossilbefund gibt es Hinweise darauf, dass es auch Insekten gegeben hat, die in allen drei Brustsegmenten (T1, T2 und T3) Flügel oder flügelähnliche Strukturen ausgebildet hatten (Grimaldi & Engel 2005) und vermutlich erst im Lauf der Zeit flügelartige Anhänge nur an T2 und T3 beschränkt blieben. Prud’homme et al. vermuten daher, dass auch im Fall der Buckelzirpen diejenigen Regulationsgene, die die Ausbildung von Flügeln unterdrücken können, an der veränderten Ausbildung von Flügeln bei Insekten und eben auch bei der Bildung der Halsschilde beteiligt sind.
Die Entstehung der Formenvielfalt der Halsschilde bei den Buckelzirpen ist allerdings nach wie vor unverstanden; dazu müsste noch eine Vielzahl weiterer genetischer Faktoren ins Spiel kommen.
Für die interessanten Befunde, die Prud’homme et al. (2011) dokumentieren, scheint allerdings beim derzeitigen Stand der Erkenntnisse die Bemerkung von der „Entstehung von Neuheit“, mit der Moczek (2011) seinen Kommentar betitelt („The origins of novelty“) voreilig und nicht durch die Befunde gedeckt.3 Die bisher bekannten Zusammenhänge sprechen eher dafür, dass ein Arsenal an genetischen Möglichkeiten erstaunlich variabel eingesetzt wird. Hier stellt sich die Frage, wo und wie die Entscheidungen fallen, welche Kombination eingesetzt wird. Bei der Ausbildung der Halsschilder von Buckelzirpen erstaunt zwar die phantastische Formenvielfalt, aber für die genetische Steuerung von deren Entwicklung scheint eigentlich alles vorhanden zu sein und es ist eher erstaunlich, dass solch exotische Anhänge unter den Insekten ausschließlich bei den Buckelzirpen auftreten. Wenn in zukünftigen Untersuchungen die genetischen Steuerelemente, die während der Ontogenese wirksam sind, beschrieben und plausibel erklärt werden, dann ist damit immer noch nicht die Art und Weise von deren (Neu-)Verwendung im Laufe der hypothetischen Stammesgeschichte verstanden. Es stellen sich Fragen nach den Umständen und den Instanzen, die die Faktoren in unterschiedlichen Zusammenhängen zur Anwendung bringen.
Literatur
Grimaldi D & Engel MS (2005) Evolution of Insects. Cambridge University Press.
Prud’homme B, Minervino C, Hocine M, Cande JD, Aouane A, Dufour HD, Kassner VA & Gompel N (2011) Body plan innovation in treehoppers through the evolution of an extra wing-like appendage. Nature 473, 83-86.
Moczek AP (2011) The origins of novelty. Nature 473, 34-35.
Anmerkungen
1 Beispiele von exotischen Buckelzirpen hat das Magazin GEO veröffentlicht und im Internet zugänglich gemacht: http://www.geo.de/GEO/natur/tierwelt/5218.html; weitere sind unter folgender Adresse zu finden: http://www.spiegel.de/fotostrecke/fotostrecke-67707.html (Stand: 18. 5. 2011), beide Fotostrecken sind sehr sehenswert!
2 Homologie: Gemeinsamkeit aufgrund (vermuteten) gemeinsamen evolutionären Ursprungs aus Vorfahren.
3 In der Tagespresse wurde die hier referierte Arbeit ebenfalls besprochen, allerdings suggerieren die Titelformulierungen viel mehr als inhaltlich geboten wird und auch die angeführten evolutionären Erklärungen erscheinen oberflächlich und sehr optimistisch. Unter der Adresse http://www.spiegel.de/wissenschaft/natur/0,1518,761084,00.html (Stand: 18. 5. 2011) wurde über „Darth Vader der Insektenwelt“ berichtet und die Befunde als „eindrucksvoller Beleg für die Macht der Evolution“ interpretiert. In der FAZ Ausgabe vom 11. 5. 2011 war zu lesen: „Wie die Evolution zum Großbildhauer wurde“.
Autor dieser News: Harald Binder
© 2011, http://www.genesisnet.info/schoepfung_evolution/n168.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
16.03.11 Pünktchen und Streifen
Warum sind die Raubkatzen so schön und doch so unterschiedlich?
Raubkatzen haben es uns schon immer angetan. Ihre Eleganz, Geschmeidigkeit und Kraft sind faszinierend. Der majestätische Löwe Aslan in C. S. Lewis’ „Chroniken von Narnia“ beweist es aktuell genauso wie auch die weniger schöne Tatsache, dass Menschen den Raubkatzen das Fell über die Ohren ziehen, in der Hoffnung, etwas von deren Attributen auf sich zu übertragen, sei es bei Schamanenkulten oder im Pelzgeschäft der Reichen und Schönen. Heute ist diese Praxis zum Glück mehr und mehr verpönt, so dass die Raubkatzen außer für Werbekampagnen auch als interessante Studienobjekte dienen.
Wissenschaftler der School of Experimental Psychology und der School of Biological Sciences in Bristol machten sich einmal mehr an die Untersuchung, warum die heute bekannten etwa drei Dutzend Katzenarten ihre speziellen Fellmuster tragen. Ihre Herangehensweise unterschied sich von früheren Untersuchungen vor allem darin, dass sie mathematische Modelle benutzten, um Fellmusterung zu beschreiben und damit in numerische Werte zu überführen. Diese Transformation erlaubte ihnen, Korrelationen zwischen verschiedenen Teilaspekten der Fellmusterung und Parametern wie Körper- oder Beutegröße, Sozialverhalten, Bewegungsart, Tag/Nachtaktivität und vor allem verschiedenen Habitaten (Lebensräume) statistisch zu analysieren. Basierend auf jeweils sechs Fotografien unterschiedlicher Individuen von 35 Arten aus der Familie der Katzen wurden die Fellmuster durch Probanden unabhängig kategorisiert und in ein mehrdimensionales mathematisches Modell überführt. In der Originalarbeit (Allen et al. 2010) kann man in einer Gegenüberstellung von mathematisch generierten Mustern zu entsprechenden Ausschnitten aus Fotos anschaulich feststellen, dass die Transformation relativ gut gelungen ist.
Korrelationsanalysen und der Vergleich der Verteilung der Felldesigns in einem molekularen Stammbaum der Katzen bestätigten statistisch, was schon früher vermutet worden war: Die Fellmusterung korreliert am ehesten mit dem Habitat der Katzen und nicht mit Abstammungszusammenhängen. Das weist darauf hin, dass Tarnung der Hauptzweck der verschiedenartigen Schönheit sein könnte. Raubkatzen, die sich in geschlossenen Umgebungen nachts vor allem in Bäumen bewegen, haben eher komplexe Musterungen, während Arten, die sich in offenen Habitaten hauptsächlich am Boden aufhalten, eher einfarbig daherkommen.
Im Vergleich zur angenommenen Stammesentwicklung, die in der vorliegenden Arbeit auf molekularen Daten beruhte, sind die Designs sehr unregelmäßig verteilt. Nah verwandte Arten tragen oft sehr unterschiedliche Musterungen, während sehr ähnliche Muster über die ganze Familie verteilt vorkommen. Dies weist klar darauf hin, dass sich die Tarnfärbung der Raubkatzen, wenn nicht gar ihre Aufspaltung selbst, in evolutiv sehr kurzen Zeiträumen herausgebildet bzw. abgespielt hat.
Statistisch wurde untermauert, dass die Musterung am ehesten der Tarnung dient, doch gibt es einige offenen Fragen. Zum Beispiel finden sich die senkrechten Streifen des Tigers in keiner anderen Katzenart auch nur entfernt wieder, und sie sind auch nicht streng mit Grasland als Habitat korreliert, was naheliegend erschiene.
Bedeutender aber sind statistische „Ausreißer“. Der Tarnungshypothese zum Trotz gehorchen als bekannteste Arten der Gepard und der Serval nicht den Statistikern: Sie halten sich mit gepünkteltem Fell in der offenen Steppe oder in buschigem Gelände auf, während weitere, weniger bekannte Katzenarten einfarbig in geschlossenen Habitaten leben. Insgesamt sind mindestens 5 der 35 untersuchten Arten deutliche „Ausreißer“. Die Autoren schlagen hypothetische Erklärungen für diese Nonkonformisten vor, geben aber zu, dass es sich um ad hoc-Argumente handelt, die nicht durch Daten untermauert sind.
So erweist sich wieder einmal eine auf den ersten Blick sehr anschauliche Tarnungshypothese beim zweiten Hinsehen als wesentlich komplizierter (wie beim berühmten Birkenspanner). Statistisch lassen sich Zusammenhänge berechnen, Voraussagen im Einzelfall halten aber der Faktenlage nicht immer stand und erfordern Zusatzhypothesen. Starke Selektionsdrücke, die für sehr schnelle Aufspaltungen nötig wären, bleiben hypothetisch. Die oben beschriebene Beobachtung, dass eine signifikante Anzahl an „Ausreißer“-Arten bis heute erfolgreich mit vermeintlich schlechterer Tarnung lebt, macht starken Auslesedruck auf die Fellzeichnung auch nicht wahrscheinlicher.
Viel einfacher lassen sich die Befunde mit der Annahme eines polyvalenten Grundtyps bei den Katzen vereinbaren (vgl. Genetisch polyvalente Stammformen von Grundtypen, https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1 und Die Katzenartigen – ein klar abgegrenzter Grundtyp, http://www.si-journal.de/index2.php?artikel=jg13/heft2/sij132-2.html. Die beobachtete rasche Aufspaltung lässt sich gut durch Rekombination und Rückgriff auf ein „Baukastensystem“ bei der Besiedelung neuer Habitate erklären. Und erneut kann man feststellen, dass Lebewesen in weit größerem Formenreichtum, Farbe und Vielfalt existieren, als man durch unbarmherziges und auf höchste Ökonomie angewiesenes „Survival of the fittest“ erwarten würde. Vielleicht ist doch ein Grund für die Vielfalt der Musterung der Raubkatzen einfach ihre Schönheit?
Quelle
Allen WL et al. (2010) Why the leopard got its spots: relating pattern development to ecology in felids. In: Proceedings of the Royals Society B. doi: 10.1098/rspb.2010.1734, 2010.
Autor dieser News: Hans-Bertram Braun
© 2011, http://www.genesisnet.info/schoepfung_evolution/n162.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
05.02.11 Hummeln lösen das Problem des Handlungsreisenden
Tiere, die Nahrung aus Reservoirs sammeln welche sich im Laufe der Zeit wieder auffüllen, besuchen die Nahrungsquellen in vorhersagbaren Sequenzen, die „traplines“ genannt werden, analog zu menschlichen Fallenstellern, die ihre Fallen in einer festen Reihenfolge inspizieren. Dieses Verhalten ist im Tierreich weit verbreitet. Dennoch ist wenig darüber bekannt, wie die Tiere diese spezifischen Reihenfolgen entwickeln. Frühere Experimente hatten ergeben, dass sich z. B. Prachtbienen im Wesentlichen an der Reihenfolge der erstmaligen Entdeckung der Nahrungsquellen orientierten, auch wenn diese zu suboptimalen Sammelstrecken führte.
Sollen die Wegstrecken verkürzt werden, müssen sich die Sammler mit einer kombinatorischen Optimierungsaufgabe herumschlagen, die als „Problem des Handlungsreisenden“ in der Mathematik bekannt ist. Die optimale Route zu finden, wenn mehrere Orte jeweils nur einmal besucht werden sollen, stellt bei zunehmender Zahl zu besuchender Orte eine Aufgabe dar, die exponentiell an Komplexität zunimmt, so dass die Berechnung der beweisbar optimalen Lösung auch Supercomputer an ihre Grenzen bringt.
- Lihoreau, L. Chittka and N. E. Raine von der Queen Mary und der Royal Holloway University in London untersuchten, in welcher Reihenfolge Hummeln (Bombus terrestris, Dunkle Erdhummel) künstliche Blüten besuchen, die nach und nach so verteilt wurden, dass die Hummeln die Sammelstrecke dynamisch entwickeln mussten. Die jeweils neuen Nahrungsquellen wurden schließlich so platziert, dass sie maximal voneinander entfernt waren, und bei Abfliegen in der Reihenfolge des Erscheinens zu einer deutlich suboptimalen, also unnötig langen Flugstrecke führen mussten. Die Experimentatoren stellten den Hummeln nur soviel Nahrung pro Blüte zur Verfügung, dass diese gezwungen waren, während eines Ausflugs alle Blüten zu besuchen, wenn sie ihren Kropf vollständig füllen wollten.
Es stellte sich heraus, dass die Hummeln bei zunehmender Komplexität der Sammelaufgabe schnell davon abkamen, die Blüten in der suboptimalen Reihenfolge der ursprünglichen Reihenfolge der Entdeckung anzufliegen, stattdessen die Routen schrittweise verbesserten und häufig sogar die optimale Route wählten. Die Insekten optimierten die mittlere Flugdistanz innerhalb der ersten 6-10 Ausflüge sehr deutlich und näherten sich am Ende der 20 protokollierten Ausflüge eng dem kürzesten Flugweg an. Die suboptimale Route der ursprünglich ersten Entdeckung spielte dann nur noch eine sehr untergeordnete Rolle. Offensichtlich lernten die Hummeln aus Versuch und Irrtum und optimierten die Routen sehr schnell. Es konnte außerdem gezeigt werden, dass sie die am ersten Tag gefundene optimale Route auch noch am zweiten Tag bevorzugt, wenn auch etwas weniger häufig, benutzten. Außerdem wurden zu Beginn des zweiten Tages immer auch ganz neue, alternative Routen gewählt. Das könnte entweder von Gedächtnislücken des kleinen Hummelhirns herrühren oder aber darauf hinweisen, dass die Hummeln die Umgebung am nächsten Morgen erst einmal nach potentiellen neuen Nahrungsquellen abscannen.
Die Studie wurde in den Medien u. a. mit „Insekten schlagen Supercomputer“ überschrieben, sie hat schon in den Wikipedia-Artikel „Problem des Handlungsreisenden“ Eingang gefunden! Tatsächlich mussten die untersuchten 11 Hummeln im Experiment die optimale Route zwischen 4 künstlichen Blumen (d. h. aus 24 möglichen Routen) wiederholt finden, was sie nach etwa 10 von 20 beobachteten Versuchen mit statistischer Signifikanz in der Mehrzahl schafften. Die optimale Route wurde am ersten Tag von 8 Hummeln im Mittel zu ca. 40% benutzt, am zweiten Tag von 4 Hummeln zu 30%. Statistisch sind die Ergebnisse signifikant, sie zeigen klar, dass nicht einfach die Route der ersten Entdeckung beibehalten wird und dass das kleine Hummelhirn sehr lernfähig ist. Die Behauptung, Hummeln hätten auf unerklärliche Weise das Problem des Handlungsreisenden effizienter als Computer gelöst, erscheint allerdings leicht extrapoliert und ist durch die Publikation (noch) nicht gedeckt.
Doch auf jeden Fall stellen die Leistungen der kleinen Hummelhirne beim Optimieren der Nahrungssuche ein weiteres erstaunliches und noch unerklärtes Beispiel für Mustererkennung, Orientierung im Raum und für Lern- und Gedächtnisleistung sogenannter „einfacher“ Lebewesen dar. Das Beispiel der lernfähigen Hummeln ist besonders eindrucksvoll, weil hier eine mathematische Aufgabe gelöst wird. Die Analogie zu menschlicher geistiger Tätigkeit ist offenkundig. Wie ein geistloser, natürlicher Evolutionsprozess den Hummeln diese strategische Fähigkeit zu vermitteln vermag, erscheint dagegen als eine schier unlösbare Aufgabe.
Literatur
Lihoreau M, Chittka L & Raine NE (2010). Travel optimization by foraging bumblebees through readjustments of traplines after discovery of new feeding locations. Am. Nat. 176, 744-757.
Autor dieser News: Hans-Bertram Braun
© 2011, http://www.genesisnet.info/schoepfung_evolution/n161.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.12.10 Noch besser als schon bekannt: das Linsenauge
Eines der beliebtesten Beispiele von vermeintlichen Konstruktionsfehlern der Lebewesen ist das Linsenauge der Wirbeltiere. Trotz atemberaubender Fortschritte im Verständnis des Aufbaus der Funktionsweise der Retina (Netzhaut) hält sich die Meinung der Fehlerhaftigkeit hartnäckig – mittlerweile gegen besseres Wissen! Hintergrund ist die inverse Lage der Netzhaut, die dazu führt, dass das Licht vor dem Auftreffen auf die lichtempfindliche Seite der Netzhaut einige Nervenfasern durchqueren muss, die von den Netzhautzellen in Richtung Glaskörper abgehen. Die abgehenden Nervenendigungen werden am blinden Fleck zum Sehnerv gebündelt, was dazu führt, dass ein kleiner Teil des Augenhintergrunds kein Licht aufnehmen kann (und daher ein „blinder Fleck“ ist). Hätte man das nicht besser konstruieren können? So wird oft gefragt und es werden entsprechende Behauptungen aufgestellt.
Es ist aber schon sehr lange bekannt, dass es gute funktionelle Gründe für die inverse Lage der Retina gibt, vor allem kann die Netzhaut dadurch sehr effektiv versorgt und Stoffwechselabfälle können entsorgt werden (vgl. Ullrich et al. 2006). Seit einigen Jahren ist aber auch bekannt, warum die der Netzhaut vorgelagerten Schichten keine Beeinträchtigung des optischen Apparats bewirken. Im Jahr 2007 war es einem Leipziger Forscherteam um K. Franze gelungen zu zeigen, dass sogenannte Müller-Zellen über Zellfortsätze verfügen, die senkrecht die Netzhaut von der Oberfläche bis zu den Sinneszellen durchlaufen und als Lichtleitkabel verlustfrei das ankommende Licht zu den Stäbchen und Zapfen führen (Franze et al. 2007; vgl. Ullrich 2008). Das Rätsel der scheinbar falsch herum liegenden Netzhaut (inverse Retina) und ihrer unerwarteten perfekten Funktionalität erhielt eine unerwartete Antwort, die weltweit Begeisterung hervorrief.
Diese Befunde wurden Anfang dieses Jahres durch Studien von A. M. Labin und E. N. Ribak, Physiker vom Israel Institute of Technology in Haifa unterstützt. Die Forscher berichteten von experimentellen Untersuchungen mit Müller-Zellen aus der Netzhaut des Menschen. Und zwar untersuchten die Forscher, wie das Licht in den Nervenfortsätzen der Müller-Zellen weitergeleitet wird. Geschieht dies unspezifisch, verlustbehaftet oder trägt der Aufbau der dünnen Kanälchen bzw. Zellfortsätze zur Optimierung der Sehfähigkeit des Auges bei? Ihr Resümee ist im wahrsten Sinne des Wortes erhellend: „Die grundlegenden Eigenschaften in der Anordnung der Glia-Zellen müssen als eine optimale Struktur betrachtet werden, die designt wurde, um die Schärfe der Bilder in der menschlichen Retina zu garantieren. Diese Tatsache besitzt eine Schlüsselrolle für die Qualität des Sehens bei Menschen und anderen Arten“ (Labin & Ribak 2010, 1).
Die Messergebnisse von Labin und Ribak dokumentieren also: Durchmesser, Länge oder die Verjüngung der Lichtleiter sind so gestaltet, dass auf der Netzhaut ankommende Lichtstrahlen optimal gesammelt (durch becherförmige Erweiterung des oberen Kabelschachtes), gefiltert (z. B. durch Ausblenden von Streulicht und von Wellenlängen außerhalb des sichtbaren Spektrums) und refokussiert (Ausgleich der Streuung des Lichtes durch Aberration der Linse) zu den Stäbchen und Zapfen gelangen. Im Ergebnis tragen alle diese Konstruktionsdetails dazu bei, die Bildinformationen verlustfrei, die resultierenden Bilder klarer und die Farben schärfer zu präsentieren. Damit liefern Labin und Ribak eine wunderbare Bestätigung und feine Ergänzung der Entdeckungen von Franzes Team. In einem kürzlich auf der Seite der International Society for Optics and Photonics („SPIE“, http://spie.org/x42206.xml?ArticleID=x42206) veröffentlichten Beitrags schreiben die beiden Autoren: „Die Retina hat ihre inverse Form ausgebildet, um die Bündelung aufgefangener Lichtstrahlen und die Sehschärfe zu verbessern, die Empfindlichkeit gegen Streulicht und Störungen herabzusetzen, um mehr Licht auf die Zapfen zu konzentrieren und um die chromatische Aberration auszuschalten.“ (Chromatische Aberration ist die unterschiedliche Brechung von verschiedenfarbigem Licht.)
Dennoch lebt das Argument von der Fehlerhaftigkeit des Linsenauges munter weiter. Um es zu „retten“ wird einfach behauptet, diese genialen Fähigkeiten seien als Ausgleich der vorhandenen Mängel entstanden (so z. B. McAlpine 2010). Zunächst seien in der Evolution fehlerbehaftete Lösungen entstanden, die anschließend erstaunlich kreativ und perfekt ausgeglichen worden seien. Es gibt angesichts des heutigen Wissens eine sehr viel einfachere und überzeugendere Erklärung, sie hat nur den „Nachteil“, dass sie die Möglichkeit einer Schöpfung ernsthaft in Betracht zieht.
Quellen
Franze K et al. (2007) Müller cells are living optical fibers in the vertebrate retina. Proc. Natl. Acad. Sci. 104, 8287-8292. (online: http://www.pnas.org/cgi/reprint/104/20/8287).
Labin AM & Ribak EN (2010) Retinal Glial Cells Enhance Human Vision Acuity. Phys. Rev. Lett. 104, 158102.
McAlpine K (2010) Evolution gave flaved eye better vision. New Scientist (2759: 6 May 2010) (http://www.newscientist.com/article/mg20627594.000-evolution-gave-flawed-eye-better-vision.html).
Ullrich H, Winkler N & Junker R (2006) Zankapfel Auge. Ein Paradebeispiel für „Intelligent Design“ in der Kritik. Stud. Int. J. 13, 3-14. (online: http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html).
Ullrich H (2008) Augenblicke – raffiniertes Design der Linsenaugen. Stud. Int. J. 15, 32-35. (online: http://www.si-journal.de/index2.php?artikel=jg15/heft1/sij151-4.html).
Autor dieser News: Henrik Ullrich
© 2010, http://www.genesisnet.info/schoepfung_evolution/n158.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
12.11.10 Das Design-Argument in der Diskussion
In der Öffentlichkeit ist es um „Schöpfung“ und „Intelligent Design“ merklich ruhiger geworden. Nach wie vor aber erscheinen national und international Beiträge und Artikel zur Frage nach der Erkennbarkeit Gottes in der Natur und zum Design-Ansatz in der Biologie.
Hansjörg Hemminger, Weltanschauungsbeauftragter der Württembergischen Landeskirche, hat in einer kürzlich erschienenen Veröffentlichung seine Ablehnung von „Intelligent Design“ begründet (Hemminger 2010a). Zuvor hatte er im Frühjahr eine Rezension des Buches „Spuren Gottes in der Schöpfung?“ (https://www.wort-und-wissen.org/produkt/spuren-gottes-in-der-schoepfung/) auf der Webseite der AG Evolutionsbiologie veröffentlicht (Hemminger 2010b). Der Autor dieses Buches, Reinhard Junker, hat Hemmingers Argumentation in zwei Artikeln kritisch analysiert.
Im ersten Artikel („Sind Gottes Spuren in der Schöpfung verwischt? Eine Antwort auf irreführende Argumente gegen den Design-Ansatz in der Biologie“, online unter https://www.wort-und-wissen.org/artikel/sind-gottes-spuren-in-der-schoepfung-verwischt-eine-antwort-auf-irrefuehrende-argumente-gegen-den-design-ansatz-in-der-biologie/) schreibt Junker in der Zusammenfassung:
„In seinem Beitrag über „Schöpfungsglaube und Intelligentes Design“ erläutert Hemminger seine Sicht, dass und weshalb schöpferisches Wirken in der Natur nicht erkennbar sei. Designer-Spuren würden durch natürliche Prozesse nachträglich verwischt und es gebe Beispiele von Design-Indizien, die natürlich erklärbar seien. Damit könne an solchen Design-Kennzeichen nichts abgelesen werden. Hemmingers Kritik am Design-Argument stützt sich jedoch auf eine verkürzte Darstellung des Design-Arguments und auf selbst formulierte Design-Kriterien, die von den Befürwortern des Design-Arguments gar nicht vertreten werden. So betrachtet er fälschlicherweise bloße Muster wie Hexenringe und Baumringe als Design-Indizien, die nach den Kriterien des in Junker (2009) dargestellten Design-Arguments gar nicht als Design-Indizien gelten können. Dazu kommt, dass er diesen Mustern fälschlicherweise Information und Bedeutung zuschreibt und in eine Reihe von Strukturen stellt, die tatsächlich Information tragen (wie das Erbmolekül DNS). Die richtige Feststellung, dass Designer-Spuren im Laufe der Zeit verwischt werden können, beweist in keiner Weise, dass dies immer der Fall ist. Vielmehr muss jeder Einzelfall geprüft werden. Hemmingers Frage nach einer experimentell kontrollierbaren Ursache „Intelligenz“ geht ebenfalls am Design-Ansatz vorbei. Es geht beim Design-Ansatz um eine historische Fragestellung, die nicht experimentell beantwortbar ist, sondern anhand von Indizien und eventuell durch Simulationen beantwortet werden muss.“
Beim zweiten Artikel („Eine Karikatur des Design-Arguments“, online unter https://www.wort-und-wissen.org/disk/d10-1/) handelt es sich um eine Entgegnung auf Hemmingers Buchrezension. Junker zeigt, dass Hemmingers Ausführungen an der Argumentation seines Buches vorbeigehen. Der Rezensent geht eingangs zwar auf den Ansatz des „spezifischen Design“ ein, der grundlegend für den Design-Ansatz ist, wie er in „Spuren Gottes in der Schöpfung?“ erläutert und vertreten wird, gibt aber die Argumentation verkürzt und sinnentstellend wieder. Zudem konstruiert er Design-Argumente, die der Autor gar nicht vertritt, um sie anschließend ebenso zu widerlegen wie das verkürzte Design-Argument. Seine Behauptung, die Evolutionsforschung habe dem Design-Ansatz voraus, dass ihre Aussagen aus Gesetzen ableitbar und überprüfbar seien, erweist sich als Zirkelschluss.
Zitierte Quellen
Hemminger H (2010a) Schöpfungsglaube und „intelligentes Design“. Eine Einführung in die Naturwissenschaft beim Spazierengehen. In: Hempelmann, R (Hg) Religionsdifferenzen und Religionsdialoge. EZW-Texte 210, Berlin 2010, S. 59-70.
Hemminger H (2010b) Über „Intelligent Design“ hinaus? Reinhard Junkers neues Buch: „Spuren Gottes in der Schöpfung?“ http://ag-evolutionsbiologie.de/app/download/3294042502/Spuren-Gottes-HH.html.
Junker R (2009) Spuren Gottes in der Schöpfung? Eine kritische Analyse von Design-Argumenten in der Biologie. Studium Integrale. Holzgerlingen. (http://www.wort-und-wissen.de/si/bio/spurengottes.html)
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2010, http://www.genesisnet.info/schoepfung_evolution/n157.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
06.10.10 Füchse ähnlich wie Hunde zähmbar
Von interessanten Domestizierungsversuchen mit Füchsen berichtet eine Forschergruppe um Lyudmilla Trut vom Institut für Zellbiologie und Genetik der Universität Novosibirsk (Trut et al. 2009). In einem über etliche Generationen andauernden Zuchtexperiment sollte die frühe Domestikation von Säugetieren reproduziert werden. Dabei wurde versucht, Rotfüchse (Vulpes vulpes) zu zähmen. Bei einem Teil der Tiere gelang die Zähmung und es stellte sich heraus, dass Hand in Hand mit der Zähmung (gleichsam als Nebenprodukt) viele Merkmale auftreten, wie sie aus der Hundezucht bekannt sind: weiße Flecken, Hängeohren, Ringelschwanz, aber auch Verhaltensmerkmale wie Kontaktsuche zum Menschen, Schwanzwedeln, Wimmern, Abschlecken. Diese Merkmale traten schon nach wenigen Generationen auf. Die Autoren schildern, dass in der sechsten Generation einige Welpen eifrig Kontakt mit Menschen suchten und die eben genannten Verhaltensweisen zeigten, und beschreiben das Verhalten als „sehr ähnlich“ dem Haushund (S. 352).
Die Befunde von Trut et al. sind auch für den Ansatz der polyvalenten Stammformen von Grundtypen interessant (vgl. Genetisch polyvalente Stammformen von Grundtypen, https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1). Füchse und Hunde gehören zum selben Grundtyp (Crompton 1993) und man kann daher annehmen, dass sie ein ähnliches Variationspotential besitzen. Dieser Ansatz wird dadurch bestätigt, dass nach Darstellung der Autoren die Variabilität sich „in immensen Raten“ angesammelt habe, die nicht durch Zufallsmutationen erklärbar seien (S. 349). Das Variationspotential hat offenbar andere Quellen. So können viele Merkmale durch Neotenie (Stehenbleiben auf Jugendstadium) erklärt werden (breitere Schädel, verkürzte Schnauzen, Hängeohren, Ringelschwänze; S. 353f.). Daher wurde schon lange vor dem Aufkommen von „Evo-Devo“ (evolutionäre Entwicklungsbiologie) vermutet, dass Änderungen bei Regulationsgenen eine große Rolle bei der Erzeugung der Variabilität der Zuchtformen spielen (S. 350, 353). Die Untersuchungen zeigten, dass die Selektion Gene betraf, die den neurohormonalen Status kontrollieren. Einige dieser Gene, die für die Verknüpfung von Zähmbarkeit und dem Level von Hormonen und Neurotransmittern verantwortlich sind, dürften zusammengebracht und bereits nach 8-10 Generationen fixiert worden sein. Dies wiederum dürfte die Aktivität vieler nachgeschalteter Gene beeinflussen, was zur Veränderung im Timing der Entwicklung geführt haben könnte. Außerdem könnten auf diese Weise phänotypisch versteckte Potentiale freigesetzt worden sein (S. 358).
Die in kurzer Zeit erreichbare morphologische Variation ist enorm, so dass die Unterschiede nach dem äußeren Erscheinungsbild für sich alleine betrachtet durchaus Unterschieden zwischen verschiedenen Säugerordnungen entsprechen, wie Trut et al. (2009, 349) vermerken.
Nicht alle Züchtungsmerkmale können jedoch als grundtypspezifisch betrachtet werden, so sind z. B. weiße Flecken an der Stirn oder Schlappohren auch von gezüchteten Arten vieler anderer Grundtypen bekannt. Solche Merkmale werden auch als „morphologische Marker der Domestikation“ bezeichnet. Die oben genannten Verhaltensmerkmale scheinen jedoch spezifisch für die untersuchten Hundeartigen zu sein.
Literatur
Crompton N (1993) A review of selected features of the family Canidae with reference to its fundamental taxonomic status. In: Scherer S (Hg) Typen des Lebens. Neuhausen-Stuttgart, S. 217-224.
Trut L, Oskina I & Kharlamova A (2009) Animal evolution during domestication: the domesticated fox as a model. BioEssays 31, 349-360.
Autor dieser News: Reinhard Junker
© 2010, http://www.genesisnet.info/schoepfung_evolution/n154.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.06.10 Ist „Intelligent Design“ schlechte Theologie?
Dieser Frage widmete sich Anfang Mai der Philosoph Michael Ruse in der britischen Zeitung „The Guardian“.1 Ruse ist in den Kontroversen um Schöpfung, Intelligent Design (ID) und Evolution kein Unbekannter; er ist unter anderem Mitherausgeber eines kontroversen Sammelbandes über ID.2 Er muss die Diskussion und die beiderseits vorgetragenen Argumente also kennen, und von einem Philosophen darf man eine differenzierte Argumentation erwarten. Umso enttäuschender sind seine im Guardian veröffentlichten Ausführungen. Sie eignen sich aber gut, um einige grundsätzliche Dinge zum Design-Ansatz in der Biologie klarzustellen, wie nachfolgend geschehen soll.
Ruse schreibt: „Wissenschaft erlaubt Gott einfach nicht als ursächlichen Faktor.“ Auch Christen unter den Wissenschaftlern wie der berühmte Evolutionsbiologe Theodosius Dobzhansky hätten niemals Gott in ihre wissenschaftliche Arbeit eingeführt; wie alle anderen Wissenschaftler waren sie „methodologische Atheisten“. – Beim Design-Ansatz geht es jedoch um eine historische Frage, nämlich um die erstmalige Entstehung z. B. einer biologischen Struktur oder des Lebens schlechthin. Das Ausblenden des Wirkens Gottes ist im experimentellen Rahmen methodisch sinnvoll; kein Befürworter von ID ist je auf den Gedanken gekommen, Gottes Wirken als Faktor bei der Erklärung eines Experiments einzuführen. Experimentelle Forschung ist im Rahmen von ID unstrittig. Wenn aber in Ursprungsfragen die Möglichkeit einer absichtsvollen Verursachung von vornherein ausgeschlossen wird, wird eine möglicherweise richtige Antwort ausgeschlossen. Wissenschaft hat dann aber nichts mehr mit Wahrheitssuche zu tun. Der atheistische Philosoph Bradley Monton schreibt in diesem Zusammenhang: „Wenn Wissenschaft wirklich dem methodologischen Naturalismus verpflichtet ist, dann folgt daraus, dass das Ziel der Wissenschaft nicht darin besteht, wahre Theorien zu bilden.“ Wenn Wissenschaft nicht die Suche nach Wahrheit ist, drohe sie zu einer Randerscheinung und zu einer „irrelevanten sozialen Praxis“ zu werden.3 (Wie man in Ursprungsfragen methodologisch vorgehen kann, wird hier beschrieben: https://www.wort-und-wissen.org/wp-content/uploads/fadenkreuz-wissenschaftstheorie.pdf. Die naturwissenschaftliche Methode der Erkenntnisgewinnung ist dabei unverzichtbar.)
Ruse weiter: „Angesichts von Enttäuschungen gibt man nicht auf, sondern versucht es erneut. Man stelle sich vor, Watson und Crick hätten die Flinte ins Korn geworfen, nachdem sich ihr erstes Modell des DNS-Moleküls als fehlerhaft erwiesen hatte.“4 – Dieses Zitat offenbart ein grundlegendes Missverständnis des Design-Ansatzes. In seinem Rahmen wird experimentelle Forschung nicht anders betrieben als im Rahmen eines naturalistischen Weltbildes – und schon gar nicht wird die die Forschung eingestellt. Im Gegenteil: Der Design-Ansatz braucht Forschung. Denn zum einen wird im Design-Ansatz mit Grenzen natürlicher Prozesse bei der Veränderlichkeit der Lebewesen gerechnet, zum anderen wird nach definierten Design-Kennzeichen gesucht. Für beides ist Forschung unabdingbar. Um mögliche Grenzen natürlicher Prozesse nachweisen oder wenigstens wahrscheinlich machen zu können, müssen biologische Systeme detailliert erforscht werden, ebenso die Variationsmechanismen der Lebewesen. Nur durch den Fortschritt der Forschung können sich Grenzen natürlicher Vorgänge abzeichnen – oder auch nicht! Und nur auf dem Wege der Forschung kann man Design-Kennzeichen entdecken, was schon oft geschehen ist, selbst wenn man gar nicht damit gerechnet hatte (vgl. https://www.wort-und-wissen.org/produkt/spuren-gottes-in-der-schoepfung/).
Ruse fährt fort: „Das bakterielle Flagellum ist komplex. Flüchte zu Gott!“ – Als ob jemals so argumentiert worden wäre. Die Anweisung müsste hier korrekterweise lauten: Untersuche das bakterielle Flagellum möglichst genau! Erforsche die Variationsmechanismen der Lebewesen detailliert! Findet man Designer-typische Kennzeichen? Zeichnen sich Grenzen für die Variationsmechanismen ab? Kann das Flagellum über Zwischenschritte mittels der bekannten Variationsmechanismen entstehen, ohne dabei eine Zielvorgabe zu machen? Je nachdem, wie diese Fragen beantwortet werden, wird sich zeigen, ob es wahrscheinlicher ist, dass das Flagellum schöpferischen oder natürlichen Ursprungs ist.
Auf diese verkehrten Ausführungen zum Design-Ansatz baut Ruse schließlich seine theologische Kritik auf: „ID ist Theologie – sehr schlechte Theologie.“ Sobald man Gott in das Tagesgeschäft einführe, erhebe das Theodizee-Problem – das Problem des Bösen – sein hässliches Haupt. Wenn Gott wunderhaft handelt, um sehr Komplexes zu erschaffen, warum verhindert er dann nicht mit vergleichsweise einfachen Mitteln Krankheiten?5
Das Problem des Leids in der Welt ist ohne Frage schwerwiegend und allemal wert, ernsthaft bedacht zu werden (vgl. dazu: Das Theodizee-Problem, https://www.genesisnet.info/index.php?Artikel=2044&Sprache=de&l=1). Aber mit dem Design-Ansatz hat das nichts zu tun. Wie Gott im täglichen Leben (im „Tagesgeschäft“) wirkt, ist kein Gegenstand des Design-Ansatzes, ebenso wenig wie die Frage, wie Gott in den regelmäßigen Abläufen der Welt wirkt. Dieses Wirken Gottes ist verborgen und wird nur durch sein Wort dem Glaubenden ein Stück weit erschlossen. Und dass Gott beständig auf verborgene Weise in der Welt wirkt, auch in den für uns unverständlichen Dingen, ist Aussage der Heiligen Schrift. Doch das liegt nicht auf der Ebene des Design-Ansatzes. In Wirklichkeit ist ID bzw. der Design-Ansatz selbst überhaupt keine Theologie, wenn er auch mit Vorstellungen über Gottes Wirken verbunden werden kann und davon inspiriert ist. Die Bibel bezeugt an vielen Stellen, dass Gott ein souveräner Schöpfer ist, der durch sein Wort augenblicklich Dinge ins Dasein bringen kann – sie bezeugt beides: Gottes beständiges und Gottes besondere Wirken. Nur auf Gottes besonderes Wirken in der Schöpfung bezieht sich der Design-Ansatz, indem er danach fragt, wie Spuren der Schöpfung erkannt werden können.
Anmerkungen
1 http://www.guardian.co.uk/commentisfree/belief/2010/may/03/religion-atheism
2 Dembski WA & Ruse M (eds) Debating Design. From Darwin to DNA. Cambridge University Press, 2004. Zu seiner Person siehe http://de.wikipedia.org/wiki/Michael_Ruse.
3 „If science really is permanently committed to methodological naturalism, it follows that the aim of science is not generation true theories. … But if science is not a pursuit of truth, science has the potential to be marginalized, as an irrelevant social practice“ (Monton B (2009) Seeking God in Science. An Atheist Defends Intelligent Design. Toronto: Broadview Press, S. 58).
4 „You don’t give up in the face of disappointments. You try again. Imagine if Watson and Crick had thrown in the towel when their first model of the DNA molecule proved fallacious.“
5 „As soon as you bring God into the world on a daily creative basis, then the theodicy problem – the problem of evil – rears its ugly head. If God works away miraculously to do the very complex, presumably in the name of goodness, then why on earth does God not occasionally get involved miraculously to prevent the very simple with horrendous consequences? Some very, very minor genetic changes have truly dreadful effects, causing people life-long pain and despair. If God thought it worth His time to make the blood clot, then why was it not worth His time to prevent Huntingdon’s Chorea?“
Autor dieser News: Reinhard Junker
© 2010, http://www.genesisnet.info/schoepfung_evolution/n151.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.01.10 Texte zu „Intelligent Design“ neu bearbeitet
Das Thema „Intelligent Design“ oder allgemeiner der „Design-Ansatz“ in der Biologie war in den letzten Jahren Gegenstand vieler kontroverser Diskussionen; zu einem großen Teil oberflächlich und oft unsachlich, aber es gab – meist weniger auffällig – auch ernsthafte und tiefergehende Diskussionen. Reinhard Junker hat die Diskussion im Buch „Spuren Gottes in der Schöpfung? Eine kritische Analyse von Design-Argumenten in der Biologie“ aufgearbeitet (https://www.wort-und-wissen.org/produkt/spuren-gottes-in-der-schoepfung/; dort ist auch ein Link zum Inhalt und Vorwort). Verschiedene Kurzfassungen dieses Buches sind seit kurzem auf Genesisnet verfügbar, sie ersetzen die bisherigen Texte und stellen gegenüber den bisheirgen Versionen auf Genesisnet eine weitgehende Neubearbeitung dar. Die Texte gibt es in der kurzen „Interessierten“- und etwas längeren „Experten“-Version, außerdem wird auch eine neue PDF-Version zum Herunterladen angeboten. Im Einzelnen:
Überblick über den Design-Ansatz: Einführung in „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1621&Sprache=de&l=1 mit folgenden Themen:
– Der Grundgedanke des Design-Ansatzes
– Eine wichtige Unterscheidung
– Warum der Design-Ansatz nach wie vor im Rennen ist
– Der Design-Ansatz in der Biologie – eine neue Art von Wissenschaft?
– Wie wird Design begründet?
– Der Analogieschluss
– Design-Indizien
(Zum „Experten“-Text und zur PDF-Version wird dort verlinkt)
Auseinandersetzung mit Kritik am Design-Ansatz: Kontroverse um „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1622&Sprache=de&l=1 mit den Themen:
– Bedeutet der Design-Ansatz Erkenntnisverzicht?
– Ist Design falsifizierbar?
– Ist das Fehlen eines Mechanismus ein Argument gegen Design?
– Erklärt Design überhaupt etwas?
– Beruht der Schluss auf Design nur auf Nichtwissen?
– Ist spezifisches Design anthropomorph?
– Der Designer ist nicht beobachtbar und nicht fassbar
– Designer-Befürworter machen keine Forschung und tragen nichts zum Wissenszuwachs bei
– Der Bezug auf einen Designer hat in der Wissenschaft keinen Platz
Neu ist ein Text, in dem auf theologische Aspekte zum Design-Argument eingegangen wird: Design und Theologie, https://www.genesisnet.info/index.php?Artikel=1627&Sprache=de&l=2
Die Detaildiskussion mit vielen zusätzlichen Quellenbelegen und Zitaten bleibt dem o. g. Buch vorbehalten.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2010, http://www.genesisnet.info/schoepfung_evolution/n140.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.09.09 Der hat Nerven!
Woher kommen die Neuronen?
In einem Artikel über die evolutive Entstehung der Nervenzellen wirft der Wissenschaftsjournalist Greg Miller einige interessante Schlaglichter. Er beginnt mit einigen staunenswerten Details über Nervenzellen. So seien Fadenwürmer mit einigen hundert Nervenzellen ausgestattet, die ausreichten, um Futter und einen Partner zu finden. Fruchtfliegen haben etwa 100.000 Nervenzellen; mit ihrer Hilfe absolvieren sie akrobatische Flugkunststücke, führen Brauttänze auf und treten Rivalen weg. Das Gehirn des Pottwals ist mit 8 kg das größte Gehirn, es ermöglicht dem Wal Reisen über Ozeane und Tauchgänge bis 1000 m. Und schließlich sei das menschliche Gehirn die Quelle von Kunst, Literatur und Wissenschaft. Doch, so Miller, ist der evolutive Beginn unklar.
Nervenzellen unterscheiden sich von anderen Zellen durch Richtungsabhängigkeit und sie können elektrische Impulse erzeugen. Nahezu alle Nervenzellen formen Synapsen, an denen das elektrische Signal über eine kurze Strecke in ein chemisches umgewandelt wird und so von einer nachfolgenden Nervenzelle wahrgenommen werden kann. Bereits Schwämme begegnen Signalen von außen mit einem elektrischen Impuls. Der Glasschwamm Rhabdocalyptus dawsoni reagiert auf Sediment in seinen Öffnungen mit einem elektrischen Impuls, der die Futter heranstrudelnden Flagellen für einige Zeit ausschaltet. Mackie schlug in 70er Jahren des vergangenen Jahrhunderts als Vorläufer Zellen vor, wie sie bei Nesseltieren zu finden sind. Bestimmte Zellen im Schirm von Quallen reagieren auf Berührung sowohl mit einem elektrischen Reiz als auch einer Kontraktion. Später habe sich dann eine Arbeitsteilung in reine Wahrnehmung (respektive Nerven) und reine Bewegung (respektive Muskelzellen) ergeben.
Doch wie immer bei solchen Szenarien sind die Probleme nicht fern. Bei Quallen entdeckte man ein ähnlich komplexes System zur Erzeugung bzw. dem Abbau von Nervenzell-Botenstoffen wie bei anderen Tieren auch. Diese Befunde lassen schließen, dass alle Gene, die zur Herstellung von Nervenzellen nötig sind, bereits in den primitivsten Organismen vorhanden sind bzw. waren. Wieder einmal muss der vermutete Vorfahr komplex gewesen sein. Wäre er das nicht, müsste man die mehrfache evolutive Entstehung annehmen, was aufgrund geringer Wahrscheinlichkeiten zur Entstehung komplexer Ähnlichkeiten aber abgelehnt wird (s. die kurze Diskussion zu dieser Problematik weiter unten). Aber woher kommt der komplexe Vorfahre? Je komplexer dieser im evolutionstheoretischen Rahmen angenommen werden muss, desto unwahrscheinlicher wird dessen Entstehung.
Ähnliche Probleme eröffnen sich, wenn man die Evolution von einem „primitiven“ Nervennetz hin zu einem Gehirn verstehen möchte. Einer Hypothese zufolge soll der letzte gemeinsame Vorfahre der Zweiseitentiere (Bilateria) bereits ein zentrales Nervengeflecht („Gehirn“) besessen haben. Doch wurde vor nicht langer Zeit bei einem Tier nahe an der Wurzel des Stammbaumes (Saccoglossus) ein diffuses Nervennetz gefunden. Nachdem aber viele moderne Bilateria ein Nervenzentrum haben, sind die Schlussfolgerungen – in welche Richtung auch immer – heikel: Wenn der Vorläufer der Bilateria ein diffuses Nervensystem hatte, dann muss ein zentrales Nervengeflecht mehrfach unabhängig voneinander entstanden sein. Unter evolutiven Annahmen eine nicht besonders sparsame Erklärung. Auch hier haben wir es mit komplexen Ähnlichkeiten zu tun, die normalerweise nach anerkannten Homologiekriterien als Belege für gemeinsame Vorfahren gewertet werden. Wird also eine mehrfache unabhängige Entstehung eines zentralen Nervengeflechts angenommen, läge ein weiteres Beispiel dafür vor, dass eine (homologe) Bauplanähnlichkeit doch nicht als Hinweis auf einen gemeinsamen Vorfahren gelten würde. Deswegen gibt es den Ähnlichkeitsbeweis für Evolution nicht (vgl. Ähnlichkeiten in der Morphologie und Anatomie, https://www.genesisnet.info/index.php?Artikel=41301&Sprache=de&l=1 und Junker 2002, Kap. 2 und 3).
Nimmt man aber an, dass der Vorfahre der Bilateria bereits ein zentrales Nervengeflecht besaß, so muss andererseits in vielen Linien (so auch in Saccoglossus) eine Rückentwicklung (Reversion) zu einem diffusen Nervensystem stattgefunden haben. Eine offensichtliche Verschlechterung, die schwierig zu erklären ist. Denn warum sollte eine nützliche Einrichtung wie ein zentrales Nervengeflecht wieder aufgegeben werden und das auch noch mehrfach?
Jedoch, so Miller, hören die Fragen hier nicht auf. Wie entstand das Myelin, welches die Nerven isolierend einhüllt? Woher kommen die Glia-Zellen, die im Gehirn vielfältige Aufgaben, u.a. in der Informationsverarbeitung, übernehmen? Zum Schluss ist auch die Frage offen, wie Nervenzellen verknüpft werden müssen, um ein Gehirn zu bauen. So sollen – heutiger Meinung zufolge – Gehirne vier- bis fünfmal unabhängig voneinander entstanden sein. Die Beziehung zwischen der Anatomie eines Gehirns und dessen Funktionsweise scheint also keineswegs festgelegt zu sein.
Literatur
Junker R (2002) Ähnlichkeiten, Rudimente, Atavismen. Holzgerlingen.
Miller G (2009) On the Origin of The Nervous System. Science 325, 24-26.
Autor dieser News: Niko Winkler
© 2009, http://www.genesisnet.info/schoepfung_evolution/n135.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.03.09 Nur Sinn im Licht der Evolution?
Einer der meistzitierten Sätze im Zusammenhang mit der Evolutionsbiologie ist der berühmte Satz des Populationsgenetikers Theodosius Dobzhansky: „Nichts in der Biologie macht Sinn außer im Licht der Evolution“ (Dobzhansky 1973). Fast beschwörend wird dieser Satz bei allen möglichen Anlässen zitiert. Man könne „die Biologie nicht verstehen, wenn man die Evolution nicht verstehe“, beklagt der Biologe und Fachdidaktiker Dittmar Graf von der TU Dortmund im Zusammenhang mit Umfragen unter Biologie-Studienanfängern.1 Von diesen würden ca. 7 % die Evolution der Lebewesen nicht als Tatsache akzeptieren.2 Das sei nicht hinnehmbar; daher müsste in der Schule Evolution früher als bisher unterrichtet werden, damit sich falsches Denken nicht verfestige. „Wenn sich aber einmal eine Meinung verfestigt hat, können auch er und seine Fachkollegen mit Argumenten nichts mehr ausrichten. ‚Da hat man keine Chance.’“3 Die Inhalte im Fach Biologie seien aber nur erklärbar, wenn man die Evolution zu Grunde lege.4
Doch was ist an Dobzhanskys Behauptung überhaupt dran? Dazu gibt es unter Evolutionsbiologen durchaus verschiedene Auffassungen. Olivier Rieppel vom Field Museum der Universität von Chicago bezeichnet diesen Satz in seinem Buch „Unterwegs zum Anfang“ bemerkenswerterweise als „Glaubensbekenntnis“ und liest aus ihm heraus, „daß die Welt aus verschiedenen Blickwinkeln betrachtet werden kann und daß sie – je nach Sicht der Dinge – dem Betrachter auch unterschiedlich erscheint.“ Und weiter: „Stellt der Forscher verschiedene Fragen an die Natur, so wird er unterschiedliche Antworten erhalten“ (Rieppel 1989, 13). Rieppel soll keineswegs für eine evolutionskritische Position vereinnahmt werden, und er will sicher nicht eine Schöpfungsperspektive empfehlen. Aber seine Sätze sprechen für sich. Er schreibt weiter: „Wird die Idee der Evolution, oder einer speziellen Evolutionstheorie wie jener Darwins, der Beobachtung vorausgestellt, so wird die Welt im Licht jener Theorie erscheinen. Die Theorie wird sich nie als falsch erweisen können, sondern stets mit der Beobachtung in Einklang stehen“ (Rieppel 1989a, 18). Dobzhanskys Satz ist Ausdruck einer Konvention, die Welt unter einem bestimmten Blickwinkel sehen zu wollen, und beschreibt keine Notwendigkeit.
Ähnlich zurückhaltend ist Mathias Gutmann, Professor für Technikphilosophie in Karlsruhe: Evolution der Lebewesen ist nur als ein Prozess „im hypothetischen Modus“ rekonstruierbar (Gutmann 2005, 259ff.). Das heißt: Evolution ist eine Rekonstruktion und bleibt wie jede Rekonstruktion eine Hypothese von der Vergangenheit und ist nicht die Vergangenheit selber. (Deshalb kann nebenbei die Evolution der Lebewesen nicht eine beobachtbare Tatsache sein wie eine Mondfinsternis.) Evolutionstheorien greifen zwar auf empirisches Wissen zurück (z. B. vergleichende Anatomie, Biochemie, Molekularbiologie, Paläontologie etc.). Für diese Bereiche der morphologischen, physiologischen oder molekularbiologischen Analyse biologischer Phänomene (z. B. des Sehvorganges oder der Nierenfunktion) gilt aber, dass eine Beschreibung gelingt, „ohne dass auf nur eine evolutionsbiologische Aussage zurückgegriffen werden müsste“ (Gutmann 2005, 259). Evolutionsbiologie ist nur unter Rückgriff auf bereits „nicht-evolutionär“ erworbenes Wissen möglich. Die Evolutionstheorie ist daher ein nachgeordneter Typ wissenschaftlicher Begründung. „Insofern wäre sie für die (in der Regel funktional orientierte) laborwissenschaftliche Praxis letztlich irrelevant“ (Gutmann 2005, 263) – also: es macht sehr vieles Sinn in der Biologie ohne das „Licht“ der Evolution! Das meiste ist der Biologie ist erforschbar und verstehbar unabhängig davon, ob die Geschichte der Lebewesen mit einer Evolutionstheorie korrekt beschrieben werden kann.
Ähnlich äußert sich auch Bock (2000, 33): „Die Grundthese, die in diesem Artikel vertreten werden soll, ist, dass alle evolutionären Erklärungen von zuvor gemachten funktionalen Erklärungen abhängen.“5 Und dieser Autor stellt daher trotz seiner evolutionstheoretischen Perspektive ausdrücklich fest, dass Dobzhansky falsch liegt: „Funktionale Erklärungen sind unabhängig von evolutionären Erklärungen, aber das Umgekehrte ist nicht wahr. Darüber hinaus ist es einfach nicht zulässig zu schließen, dass ‚Nichts in der Biologie Sinn macht außer im Licht der Evolution’ (Dobzhansky, 1973). Funktionale Erklärungen machen nicht nur bedeutenden Sinn bei Abwesenheit irgendeiner und aller evolutionären Erklärungen, sondern bilden die große Mehrheit der Erklärungen in der Biologie, der reinen und der angewandten“ (Bock 2000, 34).6
Forschung unter der Design-Perspektive. Tatsächlich erfolgt ein großer Teil der biologischen Forschung faktisch gar nicht unter der Vorgabe einer ungerichteten Evolution, sondern unter der Design-Perspektive. Angus Menuge, Philosophieprofessor an der Concordia University Wisconsin, stellt fest: „Die wissenschaftliche Analyse hängt in keiner Weise von der Annahme ab, dass das System auf Darwinsche Weise evolvierte. Sie hängt jedoch häufig von der Annahme ab, dass das System untersucht werden kann, als ob es designed wäre“ (Menuge 2007, 27f.). Dass die Design-Perspektive heuristisch (=zur Gewinnung neuer Erkenntnisse) unverzichtbar ist, ist unter Biologen selbstverständlich (vgl. Rammerstorfer 2006). Nicht umsonst müht man sich manchmal, teleologische (= zielgerichtete) Begriffe wie „Design“ aus der Terminologie nachträglich auszuschalten (Raff 2005, Bock 2009). Daher muss Dittmar Graf widersprochen werden, wenn er sagt: „Kreationistisches Denken ist gleichzeitig auch antiwissenschaftliches Denken und die Aufgabe der Universität ist natürlich, Wissenschaft zu vermitteln, insofern ist Kreationismus schädlich und muss auch intensiv bedacht und angegangen werden. Es ist ja so, dass Kreationisten vielfach von sich selber behaupten, sie würden Wissenschaft betreiben. Wenn man aber genauer hinschaut, ist es alles andere als Wissenschaft. Es ist Pseudowissenschaft.“7 Hier wird nur mit Schlagworten operiert. Graf bedauert: „Vielfach thematisiert die Schule Wissenschaftstheorie überhaupt nicht.“8 Eine differenzierte Behandlung wissenschaftstheoretischer Fragen würde aber auch deutlich machen müssen, dass Evolution als Geschichte der Lebewesen nicht als wissenschaftlich begründete Tatsache gelten kann (s. o.). Graf beklagt im Spiegel-Interview außerdem, die Studenten wüssten nicht, „dass jedes wissenschaftliche Wissen immer nur vorläufig ist und jede Aussage überprüfbar sein muss.“9 Das muss dann aber auch für Evolutionstheorien gelten – es sei denn, sie sind etwas anderes als „wissenschaftliches Wissen“.
Design und Variationsmechanismen. In einem bestimmten Sinne gehört „Evolution“ („Entwicklung“, „Auswicklung“ im Wortsinne) allerdings doch zum Leben. Denn es ist eine empirisch (= durch Erfahrung begründet) bestens begründete Eigenschaft des Lebens, wandlungsfähig zu sein, aber nur auf der Basis des bereits Vorhandenen. Man kann diese Fähigkeit als Variationsfähigkeit bezeichnen. Dieser Aspekt gehört zum Leben ebenso wie seine Geschichtlichkeit. Daher sollten Biologiestudenten natürlich Evolutionsbiologie gründlich kennenlernen. Nur wer darüber gut Bescheid weiß, kann auch begründet die Grenzen der Wandelbarkeit der Lebewesen abschätzen.
Literatur
Bock WJ (2000) Explanations in a historical science. In: Peters DS & Weingarten M (Hg) Organisms, Genes and Evolution. Stuttgart, S. 33-42.
Bock WJ (2009) Design – an inappropriate concept in evolutionary theory. J. Zool. Syst. Evol. Res. 47, 7-9.
Dobzhansky T (1973): Nothing in biology makes sense except in the light of evolution. American Biology Teacher 35, 125-129.
Gutmann M (2005) Begründungsstrukturen von Evolutionstheorien. In: Krohs U & Toepfer G (Hg) Philosophie der Biologie. Frankfurt/M, S. 249-266.
Menuge AJL (2007) Releasing Captive Teachers: How to Refute the Case for Methodological Materialism. Paper presented at the EPS Meeting, Friday 16. Nov., 2007.
Rammerstorfer M (2006) Nur eine Illusion? Biologie und Design. Marburg.
Rieppel O (1989) Unterwegs zum Anfang. Zürich, München.
Raff RA (2005) Editorial: Stand up for evolution. Evolution & Development 7, 273-275.
Anmerkungen
1 http://www.welt.de/wissenschaft/article3186530/Der-Mensch-ein-Affe-Das-kann-nicht-sein.html
2 http://www.dradio.de/dlf/sendungen/campus/923063; Sendung vom 20. 2. 09, 14,35 Uhr
3 http://www.welt.de/wissenschaft/article3186530/Der-Mensch-ein-Affe-Das-kann-nicht-sein.html
4 http://www.dradio.de/dlf/sendungen/campus/923063; Sendung vom 20. 2. 09, 14,35 Uhr
5 Original: „The basis thesis to be advocated in this paper is that all evolutionary explanations are dependent on prior functional explanations.“
6 Original: „Functional explanations are independent of evolutionary explanations, but the reverse is not true. Moreover, it is simply not valid to conclude that ‘Nothing makes sense in biology except in the light of evolution.’ (Dobzhansky, 1973). Functional explanations not only make eminent sense in the absence of any and all evolutionary explanations, but they constitute the large majority of explanations within biology, both pure and applied.“
7 http://www.dradio.de/dlf/sendungen/campus/923063; Sendung vom 20. 2. 09, 24,35 Uhr
8 http://www.spiegel.de/unispiegel/wunderbar/0,1518,608271,00.html
9 http://www.spiegel.de/unispiegel/wunderbar/0,1518,608271,00.html
Autor dieser News: Reinhard Junker
© 2009, http://www.genesisnet.info/schoepfung_evolution/n124.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.08.08 Das Genom der Hundeartigen
Hunde sind nicht nur seit vielen Generationen treue Begleiter des Menschen, sie wurden auch in gut dokumentierten Züchtungen hinsichtlich unterschiedlichster Erscheinungsbilder, Verhaltensweisen und Charaktere selektiert.
Das Genom (= Erbgut) von Hunden ist auf 78 Chromosomen verteilt (2 Geschlechtschromosomen – X und Y – und 38 autosomale Chromosomenpaare) und enthält nach derzeitigem Kenntnisstand ca. 19 000 proteincodierende Gene und insgesamt ca. 2,4 Milliarden Basenpaare (bp). E. Kirkness und Kollegen (2003) hatten anhand von Proben eines Pudelrüden eine Übersichtssequenz des Hundegenoms vorgestellt. Eine besser abgesicherte Version wurde zwei Jahre später präsentiert; dazu hatten K. Lindblad-Toh et al. (2005) das Genom einer Boxerhündin ca. 7,5-mal gelesen1; für den ersten Entwurf war die DNA-Sequenz ca. 1,5-mal1 analysiert worden.
Aufgrund der vielfältigen Erscheinungsformen der verschiedenen Rassen von Haushunden vermutete Charles Darwin (und mit ihm viele andere bekannte Biologen wie z. B. der Verhaltensbiologe Konrad Lorenz), dass die Haushunde auf eine Vielzahl unterschiedlichster Wildtiere zurückgehen, wie z. B. verschiedene Wölfe, Kojoten und Schakale. Aber schon erste genetische Studien anhand von mitochondrialer DNA (mtDNA) seit Ende der 1980er Jahre bestätigten, dass die verschiedenen Rassen der Haushunde (Canis lupus familiaris) vom Wolf (Canis lupus) abstammen.
Vor dem Hintergrund von Studien der mtDNA von Hunden und Wölfen setzen Wissenschaftler die Entstehung der Haushunde vor 40 000–135 000 Jahren an. Archäologische Hinweise auf Hunde als Begleiter des Menschen reichen dagegen nur ca. 15 000 Jahre zurück (Ostrander & Wayne 2005)2. Erweiterte genetische Untersuchungen deuten darauf hin, dass verschiedene Populationen und mehrere hundert Individuen am Ursprung der Hunde anzunehmen sind. Dieser ist geographisch wohl in Ostasien anzusiedeln.
Basierend auf den bekannten Genomdaten, begünstigt durch weit zurückreichende gute Dokumentationen durch verschiedenste Zuchtinstitutionen und motiviert durch das Interesse vieler Hundezüchter weltweit wurde in jüngster Zeit eine Vielzahl von Studien über Zusammenhänge zwischen Genotyp (= Erbgut) und Phänotyp (= Erscheinungsbild) bei Hunden veröffentlicht. Hinzu kommt, dass bei manchen Hunderassen charakteristische Krankheitsbilder beschrieben wurden, die ähnlich auch in der Humanmedizin bekannt sind. Für die Grundlagenforschung erhofft man sich durch die Kenntnisse bei Hunden eine effektivere Suche nach den (mit-)ursächlichen genetischen Grundlagen für die Krankheiten.
Vergleicht man die molekularbiologischen Daten zwischen Hund und Mensch, so ergibt sich für beide Genome eine Variabilität von 8 x 10-4 Nukleotid-Austauschen pro bp. Der genetische Unterschied zwischen Hunderassen ist mit 27,5 % deutlich größer als derjenige zwischen verschiedenen Menschengruppen, der bei 5,4 % liegt.3 Allerdings ist die genetische Homogenität innerhalb einer Zuchtlinie bei Hunden deutlich größer (94,6 %) als bei bestimmten Gruppen von Menschen (72,5%).
Die Unterschiede in Größe des Skeletts und dem Verhältnis dessen Teile zueinander ist bei Hunden größer als bei allen anderen Säugetieren und übertrifft sogar diejenigen der gesamten Familie der Hundeartigen, zu der auch Cojoten, Schakale und Füchse gehören. Auf der Basis von Studien über Portugiesische Wasserhunde – einer sehr alten Hunderasse – untersuchten Sutter et al. (2007) Zusammenhänge zwischen genetischen Merkmalen und dem Skelettbau der Hunde. Die Autoren konzentrierten sich dabei auf einen Bereich auf Chromosom 15, der eine starke Korrelation mit der Körpergröße (und -masse) aufwies. Sie identifizierten einen Bereich in der Nähe des IGF1-Gens (IGF1: insulin-like growth factor 1, Insulin-ähnlicher Wachstumsfaktor 1). Von diesem ist bekannt, dass er auch bei Mäusen und Menschen die Körpergröße beeinflusst. IGF1 tritt bei 96 % der Portugiesischen Wasserhunde in zwei Varianten (Haplotypen) auf: Haplotyp B und I. Hunde, die homozygot B (2 B-Varianten im diploiden Chromosomensatz) sind, gehören zu den kleinsten; diejenigen, die homozygot I aufweisen, die größten; heterozygote (je einmal B und I) liegen dazwischen. Sutter et al. untersuchten mit diesen Kenntnissen 353 Tiere aus 14 kleinen und 9 sehr großen Hunderassen. Die Ergebnisse bestätigen, dass IGF1 für den Kleinwuchs einer Rasse (mit-) verantwortlich ist. Darüber hinaus zeigt diese Studie einmal mehr den starken Einfluss der Züchter: in kleinen Rassen tritt der entsprechende Haplotyp homozygot auf. Auch für den Geschlechtsdimorphismus – das Phänomen, dass bei Säugertieren männliche Organismen typischerweise größer sind als die weiblichen – konnten genetische Faktoren identifiziert werden.
An Whippets (eine kleine englische Windhundrasse) konnte gezeigt werden, dass das Myostatin-Gen für das gelegentliche Auftreten von bulligen, sehr muskulösen Tieren bei dieser eigentlich zierlichen und langgliedrigen Rasse verantwortlich ist. Myostatin ist ein Protein, das als Wachstumsfaktor den Aufbau von Muskelgewebe begrenzt. In den auffallend bulligen Whippets lagen beide Myostatingene mit einer bestimmten Mutation vor, was zur Folge hat, dass die entsprechenden Proteine nur eine stark eingeschränkte Funktion aufweisen. In Hunden, die erfolgreich bei Rennen eingesetzt werden, lag typischerweise eine normale und eine mutierte Variante des Myostatingens vor.
Parker et al. (2004; Wayne & Ostrander 2007) untersuchten anhand von Mikrosatellitenmarkern4 414 reinrassige Hunde aus 85 Zuchtlinien. Unter Anwendung statistischer Verfahren (Bayesian) konnten bis auf eine Ausnahme Hunde in Cluster (Gruppen) zusammengefasst werden, die auch ihren Züchtungen entsprach. Umgekehrt konnten die einzelnen Hunde allein aufgrund ihres Genotyps mit 99 %-iger Sicherheit (die Zuordnung von 4 aus 414 war nicht eindeutig) der entsprechenden Züchtung zugeordnet werden. Dies bedeutet, dass die modernen Hunderassen unterscheidbare genetische Einheiten darstellen.
Klassische phylogenetische Analysen der Daten separieren zwar einige der gezüchteten Hunderassen (9), aber für die große Mehrheit konnte keine aussagekräftige Auflösung gefunden werden. Die Autoren führen das vor allem auf zwei Ursachen zurück: zum einem ist die Geschichte der Züchtungen komplex (dramatische Reduktion der Individuenzahl z. B. in Not- und Kriegszeiten und der Aufbau neuer Züchtungen aus Restbeständen von alten Züchtungen), zum andern gehen beim Erstellen von genetischen Distanz-Matrizen Informationen verloren, da alle genetischen Daten in eine Zahl gepresst werden, mit der der Unterschied zweier zu vergleichender Züchtungen dargestellt wird. Beide Argumente sind prinzipiell bei phylogenetischen Untersuchungen zu berücksichtigen und sind hier nur aufgrund der Datenlage so offensichtlich.
Unter weiterer Anwendung der Bayesischen Verfahren konnten vier große Gruppen unter den Züchtungen etabliert werden, wobei Wölfe eindeutig einer dieser vier Gruppen zugeordnet werden können. Diese Resultate stützen damit ein Modell der Geschichte von Haushunden, nach dem die alten Rassen zuerst in Asien und Afrika aufgetreten sind. Des Weiteren zeigen die Ergebnisse dieser Studie, dass in erstaunlich kurzer Zeit genetisch klar unterscheidbare Rassen entstehen können, wenn hoher Selektionsdruck herrscht. Für manche Zuchtlinien, deren Vertreter unter Züchtern als sehr alte Hunde gelten, wie z. B. der Pharaonenhund, zeigen die genetischen Daten kein hohes Alter, so dass man davon ausgehen muss, dass diese Hunde zwar den alten ägyptischen Hunden im Erscheinungsbild sehr ähnlich sind, aber die Zucht erst in jüngerer Vergangenheit erfolgt ist.
Crompton (1993) hatte aufgrund ausgewählter Merkmale grundsätzliche taxonomische Zusammenhänge unter den Hundeartigen (Canidae) vorgestellt und diskutiert. Die Befunde wurden von ihm auch im Rahmen des Grundtypmodells (vgl. „Heutige Grundtypen“, https://www.genesisnet.info/index.php?Artikel=1241&Sprache=de&l=1) interpretiert. Demnach kann die Familie der Canidae als Grundtyp gegen andere Säugergruppen abgegrenzt werden. Die bisher bekannten Erkenntnisse über das Genom von Hunden und dessen Auswirkungen auf deren Erscheinungsbild bestätigen die dort aufgezeigten Linien und verfeinern das Bild. Von großem Interesse ist in diesem Zusammenhang, dass am Beispiel der Hunderassen aufgrund der umfangreichen Erfahrungen und Daten sehr gut dargelegt werden kann, wie aufgrund vergleichsweise geringer genetischer Unterschiede das Erscheinungsbild der verschiedenen Hunderassen in erstaunlichem Umfang variieren kann. Am Beispiel der Hunde können auch die gravierenden und umfangreichen Konsequenzen starker Selektionsdrücke innerhalb kurzer Zeit eingehend studiert werden. Daher sollte dem Grundtyp der Canidae für die weiteren Untersuchungen von Grundtypen besondere Bedeutung zugemessen werden.
Literatur
Crompton NEA (1993) A review of selected features of the family Canidae with reference to its fundamental taxonomic status. In: Scherer S (Hg, 1993) Typen des Lebens. Pascal-Verlag, Berlin.
Kirkness EF, Bafna V, Halpern AL, Levy S, Remington K, Rusch DB, Delcher AL, Pop M, Wang W, Fraser CM & Venter JC (2003) The dog genome: survey sequencing and comparative analysis. Science 301, 1898-1903.
Mosher DS, Quignon P, Bustamante CD, Sutter NB, Mellersh CS et al. (2007) A mutation in the myostatin gene increases muscle mass and enhances racing performance in heterozygote dogs. PLoS Genet 3(5): e79. doi:10.1371/journal.pgen.0030079
Lindblad-Toh K et al. (2005) Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 438, 803-819.
Ostrander EA & Wayne RK (2005) The canine genome. Genome Res. 15, 1706-1716.
Ostrander EA (2008) Warum Hunde so verschieden sind. Spektrum der Wissenschaft (7) 48-55.
Parker HG, Kim LV, Sutter NB, Carlson S, Lorentzen TD, Malek TB, Johnson GS; De France HB, Ostrander EA & Kruglyak L (2004) Genetic structure of the purebred domestic dog. Science 304, 1160-1164.
Wayne RK & Ostrander EA (2007) Lessons learned from the dog genome. Trends Genetics 23, 557-567.
Sutter NB et al. (2007) A single IGF1 allele is a major determinant of small size in dogs. Science 316, 112-115.
Anmerkungen
1 Diese Angaben sind statistische Größen: bei der Sequenzierung wird die gesamte DNA (in vielen Kopien) in kleine Stücke zerteilt, die dann jeweils sequenziert werden. Um anschließend das ursprüngliche Genom rekonstruieren zu können, muss man aus den unzähligen Schnipseln die sich jeweils überlappenden suchen. Dies ist ungeheuer zeitaufwändig und wird mit Einsatz großer Rechnerkapazitäten erreicht. Man geht davon aus, dass man statistisch jede Base 7-8 mal analysieren muss, um einigermaßen sicher zu sein, dass die ermittelte Sequenz vertrauenswürdig ist.
2 Die Altersangaben der DNA-Analyse beruhen auf phylogenetischen Untersuchungen in Kombination mit molekularen Uhren. Dabei werden bestimmte Mutationsraten zugrundegelegt, die dann entsprechend hohe Alter ergeben. Für die archäologischen Befunde wurden übliche Methoden zur Altersbestimmung herangezogen (Radiocarbon, 14C-Methode und andere indirekte Methoden).
3 Diese Daten gewinnt man dadurch, dass von Individuen verschiedener Gruppen (z. B. Rassen) ausgewählte Abschnitte von deren Genom analysiert und miteinander verglichen werden.
4 Mikrosatellitenmarker: kurze DNA-Sequenzen wiederholen sich im Markerbereich unterschiedlich oft, wodurch sie verhältnismäßig einfach und spezifisch aufzufinden sind. Diese und andere Marker (wie z. B. SNPs) können zur Orientierung in Genomen genutzt werden. Gene in der Nähe von Markern werden anhand derselben beispielsweise in einem Erbgang verfolgt.
Autor dieser News: Harald Binder
© 2008, http://www.genesisnet.info/schoepfung_evolution/n118.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
05.05.08 Nichtreduzierbare Komplexität update
In der Diskussion um „Intelligent Design“ (ID) spielt nichtreduzierbare Komplexität („irreducible complexity“, IC) molekularer Maschinen in den Zellen eine besondere Rolle. Die Auffassung, viele Strukturen in der Organismenwelt seien nichtreduzierbar komplex, ist zwar in ihrem Grundgedanken keineswegs neu, wurde aber durch Michael Behes vielbeachtetes Buch „Darwin’s Black Box“ einer breiteren Öffentlichkeit bekannt und ist aus der Diskussion um „ID“ nicht mehr wegzudenken. Ein System ist nichtreduzierbar komplex, wenn es notwendigerweise aus mehreren fein aufeinander abgestimmten, interagierenden Teilen besteht, die für eine bestimmte Funktion benötigt werden, so dass die Entfernung eines beliebigen Teils die Funktion restlos zerstört. Ein solches System wird als „IC-System“ bezeichnet. Daran anknüpfend wird als IC-Argument formuliert: Es ist nicht möglich, ein IC-System kleinschrittig durch ungerichtete graduelle evolutive Prozesse aufzubauen. Allgemeiner lautet das IC-Argument: Es ist nicht möglich, IC-Strukturen durch irgendwelche ungerichtete evolutive Prozesse aufzubauen. Darüber hinaus gilt das IC-Argument für viele ID-Befürworter auch als Hinweis auf das Wirken eines Schöpfers. Man muss also drei Ebenen der Argumentation mit IC unterscheiden:
- Die Behauptung, es gebe IC-Strukturen,
- die Behauptung, die Evolution (bzw. allgemeiner: eine natürliche Entstehung) von IC-Strukturen sei ungeklärt (d. h. das IC-Argument ist nicht widerlegt), und
- der Schluss von IC auf das Wirken eines Designers.
Häufig wird behauptet, ID sei unwissenschaftlich oder pseudowissenschaftlich, weil es keine Testmöglichkeit gebe. Diese Kritik ist in ihrer pauschalen Formulierung nicht berechtigt. Das wird schon daran ersichtlich, dass eine große Zahl von Argumenten und Befunden vorgebracht wurden, um entweder die Existenz von IC-Strukturen zu bestreiten (1. Ebene) oder das IC-Argument (2. Ebene) zu widerlegen.
Im stark erweiterten und gründlich überarbeiteten Artikel „Nichtreduzierbare Komplexität“, https://www.genesisnet.info/pdfs/Irreduzible_Komplexitaet.pdf (PDF-Datei, 1,1 MB) von Reinhard Junker wird zunächst das IC-Argument ausführlich erläutert und auf fehlerhafte Rezeptionen des Arguments eingegangen. Im Hauptteil wird anschließend gezeigt, dass die vorgebrachten zahlreichen Kritikpunkte das IC-Argument nicht entkräften. Gegenüber der früheren Version werden weitere Kritikpunkte diskutiert. Unter anderem werden folgende Themen behandelt:
- Von redundanter zu nichtreduzierbarer Komplexität
- Von einem nützlichen zu einem unentbehrlichem Teil
- Mit Hilfe eines Gerüsts zu IC?
- Multifunktionalität
- Computermodell von S. Sadedin
- Genduplikation
- Kooption und Funktionswechsel
- Der TTSS-Apparat als Vorstufe des Bakterienmotors
- Experimenteller Nachweis der Entstehung einer IC-Struktur?
- Modularität und „exploratives Verhalten“
- Sprunghafte Entstehung einer IC-Struktur?
- Belousov-Zhabotinsky-Reaktion
- Vergleichende Biologie
Folgendes Fazit wird gezogen: „Es kann festgehalten werden, dass die Versuche, das IC-Argument zu widerlegen, bislang gescheitert sind. Das Argument kann gegen viele Einwände behauptet werden. Vorgeschlagene Entstehungswege sind unrealistisch, spekulativ oder führen nicht zu IC.“
Abschließend wird auf wissenschaftstheoretische Aspekte im Zusammenhang von nichtreduzierbarer Komplexität und „Intelligent Design“ eingegangen. Auch dieser Teil wurde gegenüber der früheren Version stark überarbeitet; manche Argumente wurden modifiziert oder sind ganz entfallen. Der Schluss auf das Wirken eines Designers kann nicht stringent gezogen werden. Doch das IC-Argument ist widerlegbar: sowohl die Reduzierbarkeit eines (vermeintlichen) IC-Systems als auch seine Evolvierbarkeit würden dieses Argument zunichte machen.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2008, http://www.genesisnet.info/schoepfung_evolution/n110.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.03.08 Evolution durch Vorprogrammierung?
Zum Buch „The Design Matrix. A Consilience of Clues“ von Mike Gene
Der unter einem Pseudonym schreibende Genetiker Mike Gene hat kürzlich das Buch „The Design Matrix. A Consilience of Clues“ veröffentlicht (Arbor Vitae Press, 2007). Er hält darin ein starkes Plädoyer für den ID-Ansatz, allerdings unterscheidet er sich in mancher Hinsicht von der „Mainstream-ID-Bewegung“, vor allem dadurch, dass er Intelligent Design („ID“) nicht als Wissenschaft betrachtet – genauer gesagt: nicht als „science“, was eher Wissenschaft im engeren Sinne von Naturwissenschaft meint. ID ist für ihn bestenfalls eine „aufkeimende Proto-Wissenschaft“ (S. xi). Gene hält Evolution nicht für falsch; sie sollte in öffentlichen Schulen auch nicht kritisiert werden.
Der Grundgedanke seines Buches ist der einer vorprogrammierten Evolution (front-loading). Damit meint er jedoch nicht eine deterministische Evolution; vielmehr sollen bestimmte Ergebnisse des Evolutionsprozesses durch vorprogrammierte Voraussetzungen und vorprogrammierte Mechanismen wahrscheinlicher eintreten als andere. Am Anfang des Lebens standen demnach Einzeller, in denen das Potential zu einer biologischen Evolution angelegt war: Evolution auf der Basis einer ausgeklügelten Planung. Gene diskutiert vier Verdachtsmomente auf intelligentes Design: 1. Analogien mit Strukturen, die erwiesenermaßen designt sind, 2. Diskontinuität zu nicht-teleologischen Prozessen (d. h. natürliche Prozesse sind weit davon entfernt, die beobachteten Phänomene hervorzubringen), 3. Rationalität (eine Analyse der untersuchten Struktur weist einen durchdachten Aufbau nach) und 4. Voraussicht (die untersuchte Struktur ist auch auf Zukunft hin angelegt). Diese vier Kriterien können weitgehend unabhängig geprüft werden und stützen je nach Tendenz der Ergebnisse einen Verdacht auf Planung. Dass dabei subjektive Bewertungen unvermeidlich sind, ist Gene bewusst (S. 272). Er stellt schon im Vorwort klar, dass es um einen Verständnisschlüssel gehe, nicht um Beweise (S. xiii).
Das Buch ist in vier Teile aufgebaut. Teil I („The Way“) steigt mit der Frage ein, wie im Rahmen des Design-Paradigmas geforscht werden kann; in Teil II („The Clues“) geht es um Befunde, die deutlich auf Design hinweisen; Thema von Teil III ist das bereits genannte Konzept des „front-loading“ und präsentiert Überlegungen, wie eine Koexistenz der Selektionstheorie und des ID-Ansatzes aussehen könnte. Diese drei Teile liefern die Basis für eine Methode, wie Strukturen auf Design hin überprüft werden können, was in Teil IV ausgeführt wird.
Der Autor beginnt seinen Argumentationsweg in Teil I mit der Lektion, die aus einer geologischen Struktur auf dem Mars gelernt werden kann, die früher bei geringer Auflösung einem Gesicht glich. Wann erkennen wir eine Struktur als designt? Zum einen braucht es eine Vertrautheit, d. h. eine Ähnlichkeit mit einem bekannten, designten Muster. Nur solches Design ist für uns erkennbar, das eine Analogie mit bereits bekanntem Design aufweist. Zum anderen muss es eine Diskontinuität geben, das heißt: die betrachtete Struktur ist weit von solchen Strukturen entfernt, die ohne Design entstehen können. Dagegen müssen weder die Identität des Designers noch seine Methoden noch der Zeitpunkt seines Wirkens bekannt sein (S. 10). Die teleologische Deutung des Marsgesichtes erwies sich schließlich durch zusätzliche Befunde als falsch: Eine höhere Auflösung zeigte Strukturen, die durch natürliche Prozesse erklärbar waren und die beiden genannten Kennzeichen nicht mehr aufwiesen. Ganz anders aber verlief die Entwicklung bei der Erforschung des Inneren der lebenden Zellen. Die „höhere Auflösung“ lässt dort zunehmend diese beiden Kennzeichen von Design hervortreten (S. 17).
Nach diesen ersten Schritten ins Thema hinein folgt mit Teil II das erste Kernstück, in dem Hinweise auf Design in der Molekularbiologie zusammengestellt werden. Die Forschung hat gezeigt, dass die Abläufe und „Maschinen“ im Zellgeschehen viel stärker technologischen Produkten ähneln, als von irgendjemanden je vorhergesagt worden war (S. 39). Dass es sich bei den Begriffen wie „Maschine“, „Programmierung“, „Code“ etc. nicht um irreführende Metaphern handeln kann, belegt Gene durch eindrucksvolle Vergleiche der Häufigkeit der Verwendung der betreffenden teleologischen Begriffe in verschiedenen Disziplinen (S. 48, 58, 60). Es hat sich gezeigt, dass der genetische Code in verschiedener Hinsicht optimal ist. Erstaunlich sind auch die Korrekturmechanismen bei der Replikation (= Kopie der DNA) und der Übersetzung der DNA in Proteine (Transkription und Translation). Zudem sind die verwendeten Nukleotide der DNA optimal, um Fehler bei der Replikation zu minimieren. All das sieht nicht nach einem „eingefrorenen Zufall“, sondern viel mehr nach vorausschauender Planung aus.
Teil III ist mit „Challenge“ überschrieben. Hier entfaltet Gene das Konzept des front-loadings in der Evolution. Sein Grundgedanke: Die Gene und die Variationsmechanismen, die in den ersten Zellen vorhanden waren, besitzen das Potential zur Evolution von vielzelligen Lebewesen und ihrer ganzen Vielfalt. Das ist allerdings nicht so zu verstehen, als seien die vielfältigen Organe bereits deterministisch angelegt. Vielmehr seien die Gene so beschaffen, dass sie auf unterschiedlichste Weise kombiniert werden und vielfach auf verschiedenste Weise genutzt werden können. Mechanismen wie Genduplikation (= Verdopplung von Genen) und Gen-Tinkering („Basteln“ mit Genen; vgl. „Gene tinkering“, https://www.genesisnet.info/index.php?Artikel=41264&Sprache=de&l=1) versteht er als Prozesse, die auf Design zurückgeführt werden können. So werde verstehbar, dass Einzeller bereits viele Gene besitzen, die ihr volles Spektrum an Aufgaben erst in vielzelligen Organismen erfüllen. Er bezieht sich hier auf einen Befund, der die Biologen in den letzten Jahren in wachsendem Maße überrascht hat: Viele Gene, die für die Bildung von Organen vielzelliger Organismen benötigt werden, gibt es schon in Einzellern, die diese Organe natürlich gar nicht besitzen. Und diese Gene erfüllen bei den Vielzellern gleichzeitig ganz unterschiedliche Aufgaben (beispielsweise spielt das Linsencrystallin auch eine Rolle im Zellstoffwechsel). „The multi-functional nature of many proteins can be unlocked across time“ (S. 163), die Proteine sind „designed to redesign“ (S. 164). Hier knüpft Gene teilweise an moderne Evolutionsvorstellungen an, wie sie vor allem von Evo-Devo-Forschern entwickelt wurden (vgl. „Evo-Devo“, https://www.genesisnet.info/index.php?Artikel=41266&Sprache=de&l=2). Deren Begrifflichkeit suggeriert in der Tat häufig ein dahinterstehendes Design.
Im abschließenden Teil IV geht es um die „Matrix“. Die vier eingangs bereits genannten Kriterien können genutzt werden, um den Verdacht auf Design zu begründen bzw. zu erhärten: Analogie mit bekanntermaßen designten Dingen, Diskontinuität zu nicht-teleologischen Prozessen, Rationalität der Strukturen bzw. Abläufe sowie Anzeichen einer Vorausschau. Diese vier Kriterien bilden als „Design-Matrix“ zum einen eine Methode, um Design zu entdecken (oder wahrscheinlich zu machen), zum anderen eine Anleitung zur Forschung. Denn die Design-Matrix leitet zu gezieltem Fragen an die Natur und zur Entwicklung testbarer Hypothesen an (Tab. 10-1, S. 270). Jedem Kriterium ordnet Gene eine Bewertungsskala von –5 bis +5 zu; aus den Werten aller vier Kriterien ergibt sich ein Durchschnittswert, der die Wahrscheinlichkeit für Design angibt. Beispielsweise haben Pseudogene den Wert –4, der genetische Code +2,5 und ein Auto den Wert +4. Die Anwendung der Design-Matrix ist zwar unvermeidlich zu einem gewissen Grad subjektiv (so halte ich den Wert 0 für das Linsenauge für deutlich zu wenig), aber ein Test ist dennoch möglich. Dies zeigt sich auch daran, dass ID-Kritiker zu zeigen versuchen, dass die vier Kriterien keine Hinweise auf Design geben, z. B. indem sie versuchen, behauptete Analogien zurückzuweisen, eine Kontinuität von verschiedenen Strukturen nachzuweisen oder die Rationalität zu bestreiten.
Kommentar: Mike Gene präsentiert mit dem Konzept eines front-loading zweifellos eine originelle Deutung der Evolution, deren Realität er nicht in Frage stellt. Dafür, dass es intelligentes Design in der Biologie gibt, vor allem was die Ausstattung der Zellen betrifft (genetischer Code, Fehlerkontrolle, Maschinencharakter usw.), legt der Autor starke Indizien vor. Ob das front-loading-Konzept schlüssig ist, wird die weitere Forschung zeigen müssen. Gene kann manche Einwände gegen seine Sicht mindestens mildern. Es könnte durchaus auch sein, dass sich durch weitere Forschung enge Grenzen für evolutive Änderungen zeigen werden. Die Design-Argumente blieben davon unberührt, denn sie finden auch im Rahmen der Grundtypenbiologie eine sinnvolle Deutung und sie sind genauso prüfbar, wenn man vom Konzept polyvalenter und flexibler Stammformen ausgeht (vgl. „Genetisch polyvalente Stammformen von Grundtypen“, https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1. Das hieße nämlich: Front-loading von Grundtypen, die als polyvalente Stammformen mit einer potentiellen Komplexität ausgestattet sind. Die Anwendbarkeit der Design-Matrix auf das Konzept polyvalenter Grundtypen ist für die drei Kennzeichen Analogie, Diskontinuität und Rationalität sofort einzusehen, gilt aber auch für das Kennzeichen der Vorausschau. Im Rahmen der Grundtypenbiologie wäre die Vorausschau allerdings wesentlich kürzer als im Evolutionskonzept. Vorausschau würde hier nur Mechanismen der Anpassung und Flexibilität in sich wandelnden Umwelten beinhalten, nicht aber ein Blick in eine ferne Zukunft.
Wie immer man über Evolution, Schöpfung und ID denkt: Die Lektüre des Buches lohnt sich unbedingt für jeden, der ernsthaft am ID-Ansatz interessiert ist.
Hinweis: Gene präsentiert in seinem Buch noch viele weitere interessante Ideen, auf die in dieser kurz gehaltenen Besprechung nicht eingegangen werden kann. Eine ausführliche Vorstellung seines Buches finden Sie unter http://evolution-schoepfung.blogspot.com/2008/03/evolution-front-loaded.html.
Autor dieser News: Reinhard Junker
© 2008, http://www.genesisnet.info/schoepfung_evolution/n108.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.02.08 Design ist Evolution?
Den Begriff „Evolution“ gab es in der Biologie bereits vor Darwin. Bis zur Mitte des 18. Jahrhunderts verband man mit ihm eine besondere Vorstellung zur Ontogenese (= individuelle Entwicklung) der Organismen. Deren frühe Entwicklung wurde als eine Art Heranwachsen oder „Auswickeln“ (lat. evolvere) eines bereits im Samen komplett ausgebildeten Individuums betrachtet (Einschachtelungs- bzw. Präformationslehre; vgl. „Biogenetisches Grundgesetz – Geschichte“, https://www.genesisnet.info/index.php?Artikel=41305&Sprache=de&l=1). In diesem Sinne sprechen wir auch heute z. B. von „Persönlichkeitsentwicklung. Da der Begriff „Evolution“ – der Wortbedeutung entsprechend – also für die Ausprägung von bereits Vorhandenem reserviert war, passte er nicht zu Darwins Abstammungs- oder Deszendenzlehre. Denn Darwin meinte eine Stammesgeschichte, in deren Verlauf immer wieder gänzlich Neues entstanden sein musste. Schließlich war „Goethe“ noch nicht in der „Amöbe“ stammesgeschichtlich präformiert (= vorgebildet). Darwin und seine Zeitgenossen nannten die „Stammesentwicklung“ daher nicht Evolution, sondern Deszendenz, also Abstammung.
Später aber wurde der Begriff „Evolution“ für die Deszendenztheorie verwendet – man könnte auch sagen: zweckentfremdet. Und heute verbindet jeder mit „Evolution“ die Stammesgeschichte, obwohl dieser Begriff dafür einen falschen Inhalt suggeriert. Und wer mit „Mikroevolution“ die tatsächliche Bedeutung des Begriffs „Evolution“ meint, nämlich das Ausprägen von bereits Vorhandenem, muss sich viel Mühe geben, nicht missverstanden zu werden (zu „Mikroevolution“ siehe „Mikro- und Makroevolution“, https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=1).
Nun zeichnet sich ab, dass den Begriff „Design“ dasselbe Schicksal ereilen könnte. Design zeigt sich in der Kunst, in der Technik, in der Architektur und anderen kreativen Tätigkeiten und schließt Planung und Zielsetzung ein (vgl. http://en.wikipedia.org/wiki/Design). In der Biologie wird der Begriff „Design“ jedoch schon lange verwendet, ohne dass damit eine kreative, planvolle Schöpfung gemeint wäre. Design in der Biologie sei vielmehr das Ergebnis vernunftloser Prozesse, ein Ergebnis von ungerichteten Mutationen, Selektion und anderen zukunftsblinden Mechanismen. Und manche Biologen betonen eigens, dass der Begriff „Design“ nur als Metapher verwendet werde, die nicht missverstanden werden dürfe, als impliziere man damit einen Designer. Nicht umsonst müssen heutzutage diejenigen, die mit dem Begriff „Design“ das meinen, was er tatsächlich bedeutet, noch ein „intelligent“ oder „intentional“ (absichtsvoll, willensgesteuert) dazu sagen: „intelligentes Design“.
In diesem Sinne will Kenneth R. Miller, Biologe an der Brown University und Schulbuchautor, nun die Besetzung des Begriffes „Design“ mit dem neuen Inhalt und die damit verbundene Zweckentfremdung gleichsam sanktionieren. Das wird jedenfalls so für sein im Mai erscheinendes Buch „Only a Theory: Evolution and the Battle for America’s Soul“ angekündigt. Miller fordert, das Design-Vokabular der Kreationisten für die Wissenschaft in Anspruch zu nehmen. „Seiner Ansicht nach könne man auch Errungenschaften der Evolution als ‚designed‘ bezeichnen, da sie klare Funktionen erfüllen“ (www.heise.de/tp/blogs/3/103522). Wissenschaft selbst basiere tatsächlich auf der Idee des Designs; für Miller ist das aber gleichbedeutend mit „Gleichmäßigkeit der Ordnung, der Funktion und des Naturgesetzes, das unsere Welt errichtet hat“. Design als Folge bloßer Naturgesetze, ohne Plan und Zielsetzung. Telepolis (www.heise.de/tp/blogs/3/103522) schreibt über Miller weiter: „Er sieht in den Vertretern der ‘Intelligent-Design’-Theorie die Sieger in der emotional geführten Schlacht um Anhänger. ‘Die Leute wollen glauben können, dass das Leben nicht zwecklos und zufällig ist.’ Dieses Gefühl könne aber auch die wissenschaftlich argumentierbare Evolutionstheorie vermitteln – wenn die Forscher ihr Vokabular überdenken.“ Das heißt aber nichts anderes, als dass Miller letztlich einen Zweck durch die Hintertür einführen will, indem er Begriffe verwendet, mit denen man Zweck und Sinn assoziiert, um auf der anderen Seite dann zu sagen, dass man es so doch nicht meine. Begriffsverwirrung nennt man das!
Gilt in Zukunft also: Design = Evolution? Nachdem der Begriff „Evolution“ Ende des 19. Jahrhunderts für die Stammesgeschichte zweckentfremdet wurde, geht es jetzt also dem Begriff „Design“ an den Kragen!
Anmerkung: Eine Gleichsetzung beider Begriffe würde bedeuten, dass ein gegenwärtig wahrzunehmendes, ständig prüfbares und beschreibbares Phänomen wie Design mit einem hypothetischen Prozess wie „Evolution“, welcher uns nur als abstraktes Modell verfügbar ist, vereint wird. Der Philosoph Alfred North Whitehead bezeichnete schon 1974 diesen Versuch der Gleichsetzung als einen „Trugschluß der unzutreffenden Konkretheit“, weil er auf der Verwechslung bzw. unzulässigen Charakterisierung von Objektklassen beruht. Denn zu dem speziellen Gegenstandsbereich der Biologie, den Organismen mit ihrem Design, kann Evolution nichts aussagen, sondern nur zu deren Veränderung. (Whitehead, Alfred N. (1974) Die Funktion der Vernunft. Stuttgart; nach: Michael Jungert (2006) Welchen Zweck soll das haben? Untersuchungen zu teleologischen und funktionalen Erklärungsansätzen in Philosophie und Biologie. Magisterarbeit im Magisterstudiengang der Otto-Friedrich-Universität Bamberg)
Autor dieser News: Reinhard Junker
© 2008, http://www.genesisnet.info/schoepfung_evolution/n106.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
11.12.07 Diskussion über „Intelligent Design“
Im Newsbeitrag „Götter und Designer bleiben draußen“ (https://www.genesisnet.info/index.php?News=95) wurde bereits auf eine besondere Ausgabe der Zeitschrift „Religion • Staat • Gesellschaft – Zeitschrift für Glaubensformen und Weltanschauungen“ (erschienen im renommierten Verlag Duncker & Humblot, Berlin) hingewiesen – sie ist besonders hervorzuheben, weil in dieser Ausgabe auf knapp 300 Seiten Pro und Contra „Intelligent Design“ (ID) diskutiert werden. Nun legte Christoph Heilig auf der Homepage der SG Wort und Wissen eine ausführliche und informative Rezension des ganzen Bandes vor (https://www.wort-und-wissen.org/rezension/diskussion-ueber-intelligent-design-religion-staat-gesellschaft-7-jahrgang-2006-2-heft/). Er widmet allen Beiträgen eine Inhaltsangabe und fügt kritische Bewertungen an. Damit bietet er einen guten Überblick über den gesamten Band. Trotz grundsätzlicher Sympathie steht er manchen Ansprüchen der ID-Befürworter skeptisch gegenüber. Das macht die Lektüre besonders reizvoll.
Hier sollen einige Streiflichter aus der Rezension wiedergegeben werden. Einer der Herausgeber der Zeitschrift, Gerhard Besier, gibt als Motivation für die Herausgabe dieses Bandes an, dass die Interessenpolitik der Etablierten nicht verhindern dürfe, dass Außenseiter zu Wort kommen. Da diesen gewöhnlich wenig Raum gewährt werde, hätten sie hier einmal das Übergewicht erhalten.
Über den Artikel von Robert Schmidt: „Götter und Designer bleiben draußen – eine kritische Diskursanalyse der Medienberichterstattung zu Intelligent Design im deutschsprachigen Raum“ wurde im eingangs genannten Newsbeitrag bereits berichtet. Der Folgebeitrag des Botanikers Hans Peter Comes behandelt „The Synthetic Neo-Darwinian Theory of Evolution and its Legacies on Research in Modern Evolutionary Biology“. Darin werden Aspekte des Artbildungsproblems diskutiert, die mit der ID-Thematik nur am Rande in Verbindung stehen.
Der nächste Artikel von Stephen C. Meyer, „A Scientific History – and Philosophical Defense – of the Theory of Intelligent Design“, bietet einen Überblick über eine Reihe von ID-typischen Themen. Heilig wertet den von Meyer propagierten „Schluss auf die beste Erklärung“ (S. 218) als einen auf negative Argumentation gestützten Analogieschluss (das heißt, man schließt auf Design, weil alle anderen Möglichkeiten der Erklärung versagen) und widerspricht Meyer, der den Schluss auf Design als positiv begründet ansieht (S. 237). Trotz mancher Kritikpunkte hält der Rezensent Meyers Zusammenstellung für einen exzellent gelungenen Einleitungsartikel.
Den Beitrag „Intelligent Design – jenseits des Schlagwortes“ von Markus Rammerstorfer hält Heilig vor allem deswegen für interessant, „weil der Autor als Europäer nicht dem amerikanischen ID-Movement und dessen politischen Zielen zuzuordnen ist. Gerade in diesem Punkt wird der Kontrast zu Meyer besonders deutlich.“ Rammerstorfer zeige auf, dass die Frage nach dem Ursprung der teleologischen Synorganisation in der Natur eine der Grundfragen der Naturwissenschaften sei, von der man zuunrecht glaubte, Darwin hätte sie beantwortet. ID als Fragestellung sei daher vor allem aufgrund ihres naturhistorischen Charakters auch heute berechtigt.
Auf knapp 80 Seiten folgen drei Diskussionsrunden, in denen Frieder Meis und Dr. Wolf-Ekkehard Lönnig die ID-Seite vertreten, während JProf. Dr. Dr. Mathias Gutmann und Willem Warnecke die Gegenseite übernehmen. Das Format, in dem die Kontrahenten hier auftreten konnten, sei zwar sehr zu loben, doch hätten die beiden Parteien die damit verbundenen Möglichkeiten eines Disputs nicht gut genutzt, da sie weitgehend aneinander vorbei diskutieren. Heilig zeigt dies anhand mehrerer Themen, die die Diskutanten ansprechen. Während Gutmann & Warnecke wissenschaftstheoretisch diskutieren, bewegen sich Lönnig & Meis großenteils auf der fachwissenschaftlichen Ebene.
Es folgt der Beitrag „Muster und Entwurfsmuster in Softwareentwicklung und Biologie – Kriterien zur Erkennbarkeit von intelligenten Ursprüngen“ von dem Mathematiker Klaus Wittlich. Der Begriff des „Musters“ sei hochinteressant, wenn es um die Merkmalsverteilung oberhalb der Grundtypebene geht. Diesem Thema widmet sich Reinhard Junker in dem darauf folgenden Artikel „Macht das Ähnlichkeitsmuster der Lebewesen nur Sinn im Licht der Evolution?“ Darin wird das Konzept der Homologie kritisch durchleuchtet und gezeigt, dass Evolution nicht aus den Ähnlichkeitsmustern erschlossen werde, sondern als Voraussetzung für deren Deutung diene, die auch anders gewählt werden könne.
Den Abschluss des Bandes macht ein Beitrag von Dr. Harald Binder, Prof. Dr. Peter Imming und Prof. Dr. Siegfried Scherer: „Was ist über die Entstehung des Lebens bekannt?“ Eine ebenso zutreffende Überschrift wäre nach Heilig die Formulierung „Was ist über die Entstehung des Lebens allgemein nicht bekannt?“ gewesen, da die aus der Primärliteratur zusammengetragenen Befunde in der Öffentlichkeit gerade nicht allgemein bekannt seinen. Heilig veranschaulicht die Diskrepanz zwischen den Einschätzungen der Fachliteratur und den populären Darstellungen mit einem besonders krassen Beispiel; gerade in dieser Hinsicht werde die Bedeutung dieses Artikels als Aufklärung deutlich und lasse ihn zu einer aktuellen Standardquelle werden.
Heilig schließt seine Rezension mit folgendem Fazit: „Alles in Allem ist dieser Band von Religion • Staat • Gesellschaft sehr erfreulich und sollte im Bücherregal eines jeden zu finden sein, der sich mit der Ursprungsfrage eingehender beschäftigt. Trotz der unterschiedlichen Qualität der einzelnen Beiträge ist die Ausgabe ein gelungener Schritt in die richtige Richtung, hin zu einer fachlichen Diskussion und weg von polemischen Diffamierungen. All das macht Mut zu ähnlich gestalteten Folgeprojekten, für welche wir hier ein solides Vorbild vorliegen haben, das aber qualitativ auch noch ‘Platz nach oben’ lässt.“
Die Rezension kann auch aus PDF von der Wort-und-Wissen-Homepage heruntergeladen werden: https://www.wort-und-wissen.org/wp-content/uploads/b30.pdf
Autor dieser News: Reinhard Junker
© 2007, http://www.genesisnet.info/schoepfung_evolution/n100.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
15.10.07 Entstehung von nichtreduzierbarer Komplexität
Erklärt ein elementar-geometrisches Computermodell die Entstehung nichtreduzierbarer Komplexität in der Biologie?
In der Diskussion um „Intelligent Design“ (Einführung in „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1621&Sprache=de&l=1) spielt nichtreduzierbare Komplexität (irreducible complexity) als „Design-Signal“ eine besondere Rolle. Bekannt wurde dieses Konzept durch Michael Behes Buch „Darwins Black Box“, das im Sommer dieses Jahres auch in Deutsch herausgegeben wurde (https://www.wort-und-wissen.org/rezension/darwins-black-box-biochemische-einwaende-gegen-die-evolutionstheorie/).
Definition und Argument. Ein System ist nichtreduzierbar komplex, wenn es notwendigerweise aus mehreren fein aufeinander abgestimmten, interagierenden Teilen besteht, die für eine bestimmte Funktion benötigt werden, so dass die Entfernung eines beliebigen Teils die Funktion des Systems restlos zerstört (nach M. Behe, Darwin’s Black Box. The Free Press, New York, 1996, S. 36). Das sich darauf beziehende evolutionskritische Argument lautet: Es ist nicht möglich, ein nichtreduzierbar komplexes System kleinschrittig durch ungerichtete, graduelle, evolutive Prozesse aufzubauen. Behe diskutiert nichtreduzierbare Komplexität bei biochemischen Systemen, und sein darauf aufgebautes evolutionskritisches Argument wird auch „nur“ dafür behauptet.
Es wurde auf vielerlei Weise versucht, die Existenz nichtreduzierbarer Komplexität in Frage zu stellen oder das darauf aufgebaute evolutionskritische Argument zu widerlegen. In einem ausführlichen Beitrag (Nichtreduzierbare Komplexität, https://www.genesisnet.info/pdfs/Irreduzible_Komplexitaet.pdf, Stand 30. 4. 2008) bin ich auf zahlreiche Kritikpunkte eingegangen.
Computermodell. In einem 2006 veröffentlichten Artikel will die Informatikerin Suzanne Sadedin (Monash University, Australien) mit einem einfachen Computermodell die schrittweise evolutive Entstehung einer nichtreduzierbar komplexen Struktur demonstrieren, die Behes Definition erfüllen soll (http://www.csse.monash.edu.au/~suzannes/files/Sadedin2006TR.pdf).
Es handelt sich um ein einfaches geometrisches Modell eines zweidimensionalen, durch Aufteilung einer Fläche in Dreiecke entstandenen Gitters. Die Knoten (Eckpunkte der Dreiecke) können im EIN- oder AUS-Zustand sein. Stehen zwei benachbarte Knoten auf EIN, bildet sich eine Verbindung zwischen ihnen. Von einer kreisförmig geschlossenen Verbindung, die mehrere Knoten durchläuft, wird nun angenommen, sie habe eine Funktion, von einer offenen hingegen, sie sei funktionslos (Abb. 1). Wird aus der kreisförmig geschlossenen Verbindung ein Element entfernt, geht folglich die „Funktion“ verloren.

Abb. 1: Simulierte „Evolution“ im Gittermodell nach Suzanne Sadedin
Ein solch einfaches System kann problemlos sukzessive durch Mutation und Selektion aufgebaut werden. Mutiert ein AUS-Knoten in den EIN-Zustand, so wird das Ergebnis der Mutation selektiert, wenn der Knoten einer kreisförmig geschlossenen Figur benachbart ist (dann bleibt die „Funktion“ „Geschlossen sein“ nämlich erhalten, s. Abb. 1 A-E). Größere Systeme sollen selektiv im Vorteil gegenüber kleineren sein. Nun können ab einer bestimmten Größe solche Knoten, die sich in der Mitte befinden, auf AUS mutieren, ohne dass die Funktion verloren geht (Redundanz!), und es kann so ein nichtreduzierbarer Zustand erreicht werden. Denn jetzt darf kein Knoten mehr auf AUS mutieren, ohne dass die „Funktion“ verlorengeht. Auch die Entfernung redundant gewordener EIN-Knoten soll selektiv begünstigt sein (Abb. 1 F; gleiche Größe mit weniger Knoten).
Kritik. Ist dieses Modell geeignet, die Entstehung nichtreduzierbarer Komplexität in der Biochemie zu simulieren? Den biochemischen Systemen, anhand derer Behe seine Behauptungen illustriert und belegt, wird der Ansatz von Sadedin nicht annähernd gerecht, und zwar aus folgenden Gründen:
- Ein zentraler Aspekt von Behes Definition fehlt: Das System, das Sadedin präsentiert, hat keine Funktion. Es wird ihm zwar eine Funktion unterstellt, aber es ist nicht ersichtlich, worin diese bestehen soll.
- Es fehlt der Aspekt der Interaktion der Teile (siehe Behes Definition oben), es sei denn, bloße Verbindungen zwischen Knoten werden als Interaktionen betrachtet. Eine solche „Interaktion“ trifft aber sicher nicht Behes Definition. Die Autorin steckt auch hier in das System etwas hinein, das es nicht hat.
- Es fehlt folglich der Nachweis, dass das Computermodell mit einem funktionalen System eines Lebewesens vergleichbar ist. Eine Modellierung, die die Vergleichbarkeit dessen, was modelliert werden soll, nicht einmal diskutiert, ist wertlos.
- Größere Systeme sollen selektiv im Vorteil gegenüber kleineren sein (s. o.). Auch hier stellt sich die Frage nach der Entsprechung zu biochemischen Systemen.
- Die EIN-Knoten im Gitter werden mit funktionalen Proteinen gleichgesetzt. Dies geht aus folgendem Zitat hervor: „Even within this short time, the evolved systems acquired levels of complexity similar to those of systems commonly cited as examples of irreducible complexity. Irreducibly complex systems produced by the model contained between 6 and 30 parts; the bacterial flagellum requires around 33 parts …“. Das ist ausgesprochen unsachgemäß, da die Entstehung neuer funktionaler Proteine oder deren Einbau in ein System nicht mit einem einzigen Schritt (wie im Modell von Sadedin) möglich ist. Der im obigen Zitat getroffene Vergleich der Verbindungen von EIN-Knoten mit der Bakteriengeißel demonstriert die Irrelevanz der Computersimulation für die Entstehung biochemischer Apparate.
- Lässt man sich dennoch auf das Modell von Sadedin ein, so ist zu beanstanden, dass die betrachtete Fläche im Modell im Vergleich zur Komplexität biochemischer Prozesse (z. B. immenser Sequenzraum von Proteinen) viel zu klein gewählt ist. Wählt man die Gitterfläche in angemessener Größe, dann wird der Ausgangspunkt (Abb. 1 A) nur sehr schwer erreicht. Außerdem würden sich dann auch verwertbare Mutationen zu selten ereignen, so dass das Modell didaktisch nicht mehr brauchbar wäre.
Fazit. Die Argumentation in dieser Veröffentlichung ist ausgesprochen oberflächlich. Der Begriff der „Komplexität“ wird zunächst so stark verallgemeinert, dass er sogar bei ganz elementaren, nach üblichem Verständnis gar nicht komplexen (also unkomplizierten) Sachverhalten verwendbar ist. Dass einfache Gebilde durch Zufall (Würfeln) erzeugt werden können, weiß aber jedes Kind. Die Arbeit geht deshalb in ihrem Ansatz und ihren Kernaussagen völlig an dem vorbei, was in der Diskussion über nichtreduzierbare Komplexität in der Biochemie erörtert werden müsste, nämlich die Entstehung von zielgerichteten Mechanismen, deren Einzelbestandteile keine abstrakten Punkte und Striche, sondern ihrerseits aufwändig herzustellende Gebilde sind.
Im Sadedin-Modell sind die Zwischenphasen zur Erreichung eines „komplexen“ Zielzustandes konzeptionell ganz offensichtlich. Dass sie später zumeist nicht mehr erkennbar sind, ist demgegenüber nachrangig. Im Falle der Biologie ist gerade die konzeptionelle Möglichkeit von Zwischenphasen eines der Hauptprobleme.
Dank: Wertvolle Hinweise für diesen Text erhielt ich von Prof. Dr. Eberhard Bertsch, Wetter, und Dr. Harald Binder, Konstanz.
Autor dieser News: Reinhard Junker
© 2007, http://www.genesisnet.info/schoepfung_evolution/n96.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
03.09.07 Mikroevolution, Makroevolution und „ID“
Eine Unterscheidung zwischen Mikroevolution und Makroevolution sollte sich an der Qualität der Veränderungen der Lebewesen und an entsprechenden unterschiedlichen Fragestellungen (kurz: Optimierung oder Konstruktion) orientieren. Nur dann kann die Frage nach der Entstehung des Neuen in der Biologie adäquat formuliert und darauf Forschung aufgebaut werden. „Makroevolution“ steht für die Entstehung neuer Konstruktionen. Dieser Vorgang kann nicht als Extrapolation von Variationsvorgängen wie Anpassungen, Spezialisierungen oder Optimierungen betrachtet werden“ (Junker 2006; vgl. Mikro- und Makroevolution, https://www.genesisnet.info/index.php?Artikel=41223&Sprache=de&l=2). Auch manche Evolutionsbiologen teilen diese Einschätzung im Wesentlichen und halten eine qualitative Unterscheidung zwischen Mikroevolution und Makroevolution für angebracht.
Gibt es einen Unterschied zwischen Mikroevolution und Makroevolution? In der Kontroverse um Schöpfung und Evolution wird die Unterscheidung zwischen Mikroevolution und Makroevolution häufig als unsachgemäße Argumentationsstrategie der Kreationisten zurückgewiesen. Tatsächlich aber wächst die Zahl der Evolutionsforscher, die einen deutlichen Unterschied konstatieren. So stellt der Evo-Devo-Forscher Gerd B. Müller (2003, 51) fest (alle nachfolgenden Zitate sind aus dem Englischen übersetzt):
„Nur wenige Prozesse, die oben zusammengestellt sind, werden durch die kanonische neodarwinistische Theorie abgedeckt. Diese betifft hauptsächlich die Häufigkeit von Genen in Populationen und die Faktoren, die für ihre Variation und Fixierung verantwortlich sind. Obwohl sie sich auf phänotypischer Ebene mit der Modifikation existierender Teile befasst, zielt die Theorie weder auf die Erklärung des Ursprungs der Teile noch auf die Erklärung ihrer morphologischen Organisation noch der Innovation ab. In der Welt des Neodarwinismus war der motivierende Faktor für morphologische Veränderung natürliche Selektion, die für die Modifikation und den Verlust von Teilen verantwortlich gemacht werden kann. Selektion besitzt aber keine innovative Fähigkeit: sie eliminiert oder erhält, was existiert. Die generativen und Ordnungsaspekte der morphologischen Evolution fehlen daher der Evolutionstheorie.“ Hier wird klar zwischen „Variation“, „Modifikation“, „Verlust“ und „Erhaltung des bereits Existierenden“ einerseits und „Ursprung der (Bau-)Teile“, „morphologischer Organisation“ und „Innovation“ andererseits unterschieden. Außerdem: Nur Ersteres werde durch den Neodarwinismus erklärt; Letzteres sei darin abwesend.
Ähnlich schreiben Müller & Newman (2003, 3; Hervorhebung im Original): „Die Frage, warum und wie bestimmte Formen in der organismischen Evolution auftauchen, betrifft nicht die Erhaltung (und die quantiative Variation) des Bestehenden, sondern vielmehr die Neuentstehung in einem qualitativen Sinne. Diese kausale Frage nach den spezifischen Bildungsmechanismen, welche dem Ursprung und der Neuentstehung phänotypischer Merkmale zugrunde liegen, kann wahrscheinlich am besten durch den Begriff Entstehung („origination“) ausgedrückt werden. … Das Auftauchen spezifischer phänotypischer Konstruktionselemente darf nicht als durch natürliche Selektion verursacht betrachtet werden; Selektion kann nur am bereits Existierenden wirken.“
Diese beiden Autoren formulieren als Ziel des von ihnen herausgegebenen Sammelbandes Origination of Organismal Form, „die Unterscheidung zwischen Entstehung (Innovation) und Diversifikation (Variation) von Formen herauszuarbeiten, indem auf die Pluralität kausaler Faktoren abgehoben wird, die für den vernachlässigten ersteren Aspekt verantwortlich sind, nämlich den Ursprung der organismischen Form.“ Evolutionäre Innovation halten sie für nicht erklärt: „Eine der größeren Lücken in der kanonischen Evolutionstheorie besteht darin, dass man damit scheiterte, diesen Aspekt einzubeziehen.“ Damit aber fehlt der kausalen Evolutionsforschung Entscheidendes. Denn: Ohne „arrival of the fittest“ gibt es auch kein „survival of the fittest“.
Im gleichen Artikel stellen diese beiden Autoren 24 offene Fragen aus vier Teilgebieten der Evolutionsforschung zu den Mechanismen der Makroevolution zusammen. Die Fragen könnten kaum grundsätzlicher sein, und es kann wohl keinen Zweifel geben, dass angesichts eines solchen Bergs offener Fragen und angesichts des von den Autoren konstatierten qualitativen Unterschieds von Mikro- und Makroevolution die Mechanismen der Makroevolution als nicht geklärt gelten müssen. Nachfolgend sind neun dieser 24 Fragen und drei weitere Gebiete mit offenen Fragen in Übersetzung zusammengestellt:
Offene Fragen zur morphologischen Evolution nach Müller & Newman (2003)
- Burgess shale-Effekt: Weshalb entstanden die Baupläne der Vielzeller explosionsartig?
- Homoplasie: Weshalb entstehen ähnliche Gestalten unabhängig und wiederholt?
- Konvergenz: Weshalb produzieren entfernt verwandte Linien ähnliche Designs?
- Homologie: Weshalb organisieren sich Bauelemente als fixierte Baupläne und Organformen?
- Neuheit: Wie werden neue Elemente in bestehende Baupläne eingeführt?
- Modularität: Weshalb werden Design-Einheiten wiederholt verwendet?
- Constraint: Weshalb sind nicht alle Design-Optionen eines phänotypischen Raums verwirklicht?
- Atavismen: Weshalb erscheinen Merkmale, die lange Zeit in einer Linie verschwunden waren, erneut?
- Geschwindigkeit: Weshalb sind die Raten morphologischer Veränderungen ungleich?
Müller & Newman listen weitere 15 offene Fragen aus diesen Gebieten auf:
- Beziehung zwischen Genotyp und Phänotyp in Ontogenese und Phylogenese
- Epigenese und ihre Rolle in der morphologischen Evolution
- Theorie der morphologischen Evolution
Müller & Newman (2003, 7) folgern: „Mit anderen Worten: der Neodarwinismus hat keine Theorie für die Entstehung. Daraus folgt, dass die gegenwärtige Evolutionstheorie vorhersagen kann, was erhalten bleibt, aber nicht, was neu erscheinen wird.“ Und weiter: „Es fehlt immer noch eine Evolutionstheorie, die speziell die morphologischen Aspekte der Evolution betrifft und die Wechselwirkungen zwischen epigenetischen und genetischen Aspekten integriert.“
Ähnlich fasst Theissen (2006) den Begriff „Makroevolution“: „Ich werde den Begriff Makroevolution in einem engeren Sinne nur für diejenigen Evolutionsprozesse verwenden, die Innovationen (oder Neuheiten) hervorbringen oder Veränderungen in den Körperbauplänen“; dann stellt er fest: „Indem daran festgehalten wurde, dass Evolution graduell sein müsse und dass makroevolutionäre Muster vollständig und alleine durch das Wirken der natürlichen Selektion und durch Anpassung an die Umwelt erklärt werden könnten, machte die Synthetische Theorie überzogene Behauptungen und verließ daher den Bereich der Wissenschaft und entwickelte sich zu einer Ideologie (Wagner und Laubichler, 2004). … Während jedoch diese Prinzipien leicht erklären mögen, wie irgendeine Art von Organismus Ausgangspunkt für einen optimierten Organismus wird (welches Kriterium man auch zugrunde legt), ist kaum erkennbar, wie sie den Ursprung beispielsweise der Eukaryoten, der Pflanzen und Tiere von Prokaryoten, erklären kann. Studien an digitalen Organismen legen nahe, dass komplexe Funktionen durch Zufallsmutationen und natürliche Selektion entstehen können (Lenski et al., 2003), doch in welchem Ausmaß solche in silico-Studien evolutionäre Ereignisse in lebenden Organismen widerspiegeln, bleibt unklar“ (S. 352; Hervorhebung im Original. Die Studien von Lenski et al. [2003] können in der Tat nicht als Modellierung von Makroevolution gelten, wie Bertsch & Waldminghaus [2005] gezeigt haben; vgl. Evolution virtueller Lebewesen, https://www.genesisnet.info/index.php?Artikel=41246&Sprache=de&l=1).
Aus paläontologischer Perspektive sieht Jablonski (2005, 511) die Sache ähnlich: „Das widersprechende Verhalten der Taxa, die einerseits als Gattungen eingestuft werden, andererseits als Ordnungen, legt nahe, dass der Ursprung neuer Designelemente, die das Potential zur Diversifikation und zur Ansammlung weiterer abgeleiteter Merkmale besitzen, von Faktoren gesteuert wird, die sich von denen unterscheiden, die den Ursprung der Arten verursachen und welche einfach nur weitere Arten und Gattungen hervorbringen.“
„Intelligent Design“ auf der Lauer? Angesichts der eingestandenen offenen Fragen zu den Mechanismen der Makroevolution stellt Theissen (2006, 365) etwas besorgt fest: „Es ist gefährlich, Aufmerksamkeit auf die Tatsache zu lenken, dass es keine befriedigende Erklärung für Makroevolution gibt. Man wird leicht eine Zielscheibe der orthodoxen Evolutionsbiologie und ein falscher Freund von Befürwortern nicht-wissenschaftlicher Konzepte. Nach Ansicht der ersteren kennen wir bereits alle relevanten Prinzipien, die die Komplexität und Verschiedenartigkeit des Lebens auf der Erde erklären; für die letzteren wird Wissenschaft und Forschung nie in der Lage sein, eine schlüssige Erklärung zu liefern, ganz einfach weil komplexes Leben keinen natürlichen Ursprung hat. Aus einer heuristischen Perspektive sind beide Positionen unbefriedigend“ (Hervorhebungen nicht im Original). Wie man dem Problem der Erklärung von Makroevolution aus dem Weg gehen kann (erste Alternative in Theissens Zitat), liefert beispielhaft folgende Definition Futuymas: Makroevolution: „ein vager Begriff für die Evolution großer phänotypischer Veränderungen, die gewöhnlich groß genug sind, um die die veränderte Abstammungslinie und ihre Nachfahren in eine abgegrenzte Gattung oder ein höheres Taxon abzuteilen“ (zit. in Carroll 2001). Diese Definition verschleiert das Problem „Makroevolution“ und verhindert dadurch Ansätze zu deren Lösung. Man kann Makroevolution viel schärfer definieren (s. o.). Nicht der Begriff ist vage, wie Futuyma behauptet, sondern seine Definition. Die zweite Alternative in Theissens Zitat liefe darauf hinaus, Forschung in Ursprungsfragen zu unterlassen.
Es gibt aber noch einen dritten Weg: Makroevolution möglichst scharf definieren, offen einräumen, was ungeklärt ist und ergebnisoffen weiterforschen. Ergebnisoffenheit schließt in der naturwissenschaftlichen Forschung die Möglichkeit ein, dass Makroevolution als Prozess tatsächlich nicht belegt werden kann oder dass Versuche einer natürlichen Erklärungen für die Hypothese „Makroevolution“ permanent scheitern. Ergebnisoffenheit heißt auch, die Option „Planung“ einzubeziehen, und Forschung zu betreiben, um (klar definierte) Indizien auf Planung festzustellen oder als unplausibel zu erweisen. Das wäre dann Forschung im Sinne des „Intelligent Design“-Ansatzes. Dieser Ansatz würgt Forschung weder ab (vgl. Ist „Intelligent Design“ wissenschaftsfeindlich?, https://www.genesisnet.info/index.php?Sprache=de&Frage=86&FG=4) noch führt er neue Methoden ein, sondern wirft Fragen auf, deren Beantwortung zu Erkenntnissen führen könnte, die man nicht gewinnen würde, wenn man diese Fragen nicht stellen würde.
Um abschließend den Bogen zur Erforschung der Makroevolution zu schlagen: Ergebnisoffene Forschung zu den Mechanismen der Makroevolution stellt strenggenommen nicht die Frage: „Welches sind die Mechanismen der Makroevolution?“ sondern: „Wie weit reichen die experimentell demonstrierten Evolutionsmechanismen?“
Literatur
Bertsch E & Waldminghaus T (2005) Evolution virtueller Lebewesen? Stud. Int. J. 12, 34-35.
Carroll SB (2001) The big picture. Nature 409, 669.
Jablonski S (2005) Evolutionary Innovations in the Fossil Record: The Intersection of Ecology, Development, and Macroevolution. J. Exp. Zool. 304B, 504-519.
Junker R (2006) Zur Abgrenzung von Mikroevolution und Makroevolution. Stud. Int. J. 13, 59-67.
Müller GB (2003) Homology: The Evolution of Morphological Organization. In: Müller GB & Newman SA (eds) Origination of Organismal Form. Beyond the Gene in Developmental and Evolutionary Biology. Vienna Series in Theoretical Biology. Cambridge, MA, pp 51-69.
Müller GB & Newman SA (2003) Origination of Organismal Form: The Forgotten Cause in Evolutionary Theory. In: Müller GB & Newman SA (eds) Origination of Organismal Form. Beyond the Gene in Developmental and Evolutionary Biology. Vienna Series in Theoretical Biology. Cambridge, MA, pp 3-12.
Theissen G (2006) The proper place of hopeful monsters in evolutionary biology. Theor. Biosci. 124, 349-369.
Autor dieser News: Reinhard Junker
© 2007, http://www.genesisnet.info/schoepfung_evolution/n93.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.03.07 Warum Intelligent Design in der Ursprungsfrage eine Existenzberechtigung hat
Thomas Waschke veröffentlichte auf dem Blog „Evolution und Schöpfung“ (http://evolution-schoepfung.blogspot.com/) am 4. März 2007 einen Gastbeitrag zum Thema „Warum ich davon ausgehe, dass ID im Bereich der Naturwissenschaften zumindest derzeit keine Existenzberechtigung hat“. Einen Tag später erschien dieser Beitrag unter dem Titel „Warum Intelligent Design (ID) im Bereich der Naturwissenschaften derzeit keine Existenzberechtigung hat“ bei der AG Evolutionsbiologie (http://www.evolutionsbiologen.de/waschke_id.html).
Zunächst stimme ich zu, dass die Kernaussage von „Intelligent Design“, dass manche Kennzeichen der Lebewesen auf einen intelligenten Urheber hinweisen (nicht: beweisen), über die Naturwissenschaft hinausgeht. Das gilt aber vice versa auch für die Evolutionslehre im Sinne der Gesamtanschauung einer allgemeinen Evolution, d. h. eines historischen Ablaufs einer evolutionären Entstehung aller Lebewesen, welche durch zahlreiche Evolutionstheorien wissenschaftlich repräsentiert wird. Dies kann man anhand des Trilemmas deutlich machen, in das Thomas Waschke den ID-Ansatz nach einem Zitat von Martin Neukamm führen möchte:
Das Trilemma. „Entweder, ID kann zeigen, dass es sich bei den postulierten Schöpfungsakten um Erfahrungstatsachen handelt. Die zweite Möglichkeit bestünde darin, ‘Intelligent Design so auszuformulieren, dass es über ein System von Gesetzesaussagen und Mechanismen verfügt, die sich derart konsistent und erklärungsmächtig in die Wissenschaftslandschaft eingliedert, dass es eine ernstzunehmende Konkurrenz zu evolutionären Betrachtungen darstellt.’“ Oder ID müsse drittens eine eliminative Induktion leisten, d. h.: „Man müsste alle denkbaren ‘Pfade’ der organismischen Entwicklung kennen und zeigen können, dass bestimmte Merkmale unter den einst herrschenden Randbedingungen nicht entstehen konnten“ (Zitat von Martin Neukamm (http://www.evolutionsbiologen.de/junker_axonas.pdf) in Waschkes Artikel).
Da ID weder zu den Eigenschaften des Designers noch zu dessen Methode der Schöpfung irgendwelche Aussagen mache, stünden die beiden ersten genannten Alternativen nicht zur Verfügung.
Das Trilemma evolutionstheoretisch. Doch wie sehen diese beiden ersten Möglichkeiten im Hinblick auf die verschiedenen Evolutionstheorien aus?
- Der postulierte Evolutionsablauf stellt sicher ebenfalls keine Erfahrungstatsache dar; er wird nur anhand von Indizien hypothetisch rekonstruiert. Die bezüglich Mechanismen einzigen derzeit beobachtbaren Tatsachen sind meist geringfügige Veränderungen, deren Extrapolation in Entwicklungssackgassen führt (das gilt auch für größere Änderungen im Sinne des Evo-Devo-Ansatzes). Darüber hinaus wird die postulierte Evolution nur durch unterschiedliche Analogieschlüsse unterstützt, nicht durch Extrapolation heutiger Prozesse (s. u.).
- Diese Evolutionstheorien verfügen ebenfalls nicht über ein System von Gesetzesaussagen und Mechanismen, wenn es um Makroevolution geht. Den Evolutionsalgorithmus, aus dem Makroevolution erschlossen werden kann, gibt es nicht. Stattdessen wird regelmäßig auf vergleichend-biologische Betrachtungen zurückgegriffen, wenn es um die Beschreibung oder gar Erklärung bei der Entstehung evolutionärer Neuheiten geht. Es geht in der anstehenden Frage um die Existenzberechtigung von „ID“ aber gerade um Mechanismen. Dass die bekannten Mechanismen keine Makroevolutionsprozesse begründen, habe ich zusammenfassend im Artikel „Was erklärt die Evolutionstheorie“ dargestellt (https://www.wort-und-wissen.org/wp-content/uploads/a03.pdf; Details dazu in „Evolution – ein kritisches Lehrbuch“: https://www.wort-und-wissen.org/publikationen/lehrbuch/).
Der Zwischenstand ist somit nach zwei Dritteln 0:0.
Design-Signale und Analogieschluss. Thomas Waschke wirft zu Beginn des dritten Teil des Trilemmas die Frage auf, „wie ID nun trotzdem als Wissenschaft betrieben werden kann“. Er verweist hier darauf, dass ID den Anspruch erhebt, eine Signalerkennungstheorie zu sein. Gemeint ist damit, dass es Kennzeichen von Lebewesen (sog. „Design-Signale“) gebe, die auf einen intelligenten Urheber hinweisen (s.o.). Eine Signalerkennungstheorie sei jedoch – so Waschke – nicht möglich, wenn man gar nicht wisse, was ein Signal sei.
Hier verweisen ID-Befürworter jedoch auf Kennzeichen von menschlichem Design, als da wären: Zweckmäßige Konstruktionen mit funktioneller Interdependenz und nichtreduzierbarer Komplexität, Modularität und Wiederverwendung von Bauelementen und vieles andere (was hier aus Platzgründen nicht dargestellt werden kann). Durch die Analogie mit technischem Design ist also sehr wohl klar, was ein Design-Signal ist. Damit ist zugleich ein Kriterienkatalog für die wissenschaftliche Prüfung vorgegeben. Dies erkennt Waschke aber nicht an. Begründung: Der Analogieschluss von technischem auf lebendiges Design sei „nicht möglich, denn die interessanten Systeme, nämlich die Lebewesen, haben eine Eigenschaft, die sie von allen genannten Systemen unterscheidet: sie sind zur Selbstvermehrung fähig, eben zu ‘descent with modification’.“ Das sei „genau der Punkt, der den Schluss von Artefakten auf Naturgegenstände unmöglich macht.“ Da Waschke meinen Genesisnet-Artikel über ID zitiert (Kontroverse um „Intelligent-Design“, https://www.genesisnet.info/schoepfung_evolution/p1622.php, dort der Abschnitt „Was bringt die Fortpflanzungs- und Evolutionsfähigkeit?“), wundert es mich, dass er nicht auf meine Entgegnung dieses Gegenarguments eingeht. Auch im Internet-Artikel „Wissenschaft im Rahmen des Schöpfungsparadigmas“ befasse ich mich mit dieser Frage (https://www.wort-und-wissen.org/wp-content/uploads/a02.a02.pdf, S. 50f.).
Die Antworten lauten dort:
- Die Tatsache, dass Lebewesen fortpflanzungs- und evolutionsfähig sind, stellt den Analogieschluss nicht in Frage, weil nie gezeigt wurde, dass diese Eigenschaften etwas mit der Entstehung von Design zu tun haben, und weil diese Eigenschaften das tertium comparationis daher nicht in Frage stellen.
- Das zusätzliche Merkmal der Fortpflanzungs- und Variationsfähigkeit erfordert nach allem, was wir über die biologischen Grundlagen sich fortpflanzender und entwickelnder Systeme wissen, eine hochvernetzte Interaktion zwischen Informationsträgern und den korrespondierenden morphologisch-funktionellen Merkmalen. Das biologische Design dafür verweist erst recht auf intelligente Planung. Der Schuss geht also nach hinten los. Auf dem Blog von Christoph Heilig hat Klaus Lange vor kurzem einen Beitrag veröffentlicht, in dem er zeigt, dass man programmiertechnisch mittlerweile in der Lage ist, diese Merkmale von Lebewesen ebenfalls teilweise „nachzumachen“ (http://evolution-schoepfung.blogspot.com/2007/03/klaus-lange-zur-vergleichbarkeit-von.html).
- Lebewesen besitzen nichtreduzierbar komplexe Strukturen, die nach gegenwärtigem Kenntnisstand durch natürliche Vorgänge nicht sukzessive entstehen können. Das Mehr-Generationen-Argument hilft in solchen Fällen nicht weiter. Hinzu kommt noch: Solange es Leben noch nicht gab, waren die Merkmale „Fortpflanzungs- und Evolutionsfähigkeit“ noch gar nicht etabliert.
Wir wissen also aus vielerlei Beispielen der menschlichen Kultur, dass Design durch intelligente Planung entsteht, während es keine Erfahrungstatsache ist, dass Design durch ungelenkte Prozesse entsteht.
Machen wir uns das Argument an einem Beispiel klar: Thomas Waschke schreibt: „Wir kennen die Eigenschaften beispielsweise von Stein hinreichend, um beurteilen zu können, dass die bekannten Mechanismen der Erosion keine Präsidentenköpfe aus dem Gestein des Mt. Rushmore formen können.“ Nach heutigem chemischem Wissen kann man mit Fug und Recht entsprechend argumentieren: Wir kennen die Eigenschaften kettenförmiger Makromoleküle wie Proteine und DNA hinreichend, um festzustellen, dass ihre Synthese und Existenz zwar im Einklang mit den bekannten physikalisch-chemischen Gesetzmäßigkeiten stehen, diese aber für eine plausible präbiotische (zufällige, ungesteuerte) Synthese der Makromoleküle nicht hinreichend sind. Das ist eine empirisch bestens begründete und vielfach bewährte Aussage.
Aus alledem folgt freilich nicht, wer das Design verwirklicht hat. Darum geht es bei ID aber auch gar nicht. Ohne Offenbarung kann es keine Identifikation des Designers der Lebewesen geben. Einen wissenschaftlich zwingenden Schluss auf einen göttlichen Designer ist daher unmöglich. (Daher stimmt dieser Satz: „Aber niemand käme auch auf die Idee, einen Faustkeil als Argument für Außerirdische zu verwenden.“ Er wird an ihm aber Design-Signale erkennen.)
Das positive Analogie-Argument bewährt sich und ist daher alles andere als ein gescheiterter Versuch wissenschaftlicher Deutung.
Argumentum ad ignorantiam. An dieser Stelle seien noch einige Anmerkungen zum „argumentum ad ignorantiam“ angefügt, welches besagt: Wir wissen nicht, wie lebendige Systeme ohne planerische Eingriffe entstehen konnten, also haben sie Planung benötigt. Thomas Waschke behauptet, die Naturwissenschaft würde eben doch Mechanismen kennen, die zumindest das Potenzial besitzen, Neuheiten in der Evolution entstehen zu lassen. Das bestreite ich. Wenn die gegenwärtig favorisierten Mechanismen, die nur zum Teil experimentell fassbar sind, extrapoliert werden (wie Waschke empfiehlt), gelangen wir nicht zu einer allgemeinen Evolution aller Lebewesen, sondern in phylogenetische Entwicklungssackgassen. Das Variationspotential, das in den Lebewesen (empirisch nachweisbar) steckt, wird zunehmend ausgereizt. (Dies wird im Detail an anderer Stelle begründet: „Artbildung“, https://www.genesisnet.info/index.php?Artikel=41244&Sprache=de&l=1 und „Genetisch polyvalente Stammformen von Grundtypen“, https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1.) Daher kann die Evolutionstheorie nicht mit dem Vorsprung einer Extrapolationsmöglichkeit gegenüber ID auftrumpfen, während ID nur einen vermeintlich fragwürdigen Analogieschluss anzubieten habe. Tatsächlich wird Makroevolution selbst mit Analogieschlüssen begründet, so vor allem durch vergleichend-biologische Daten (Ähnlichkeiten der Lebewesen). Warum dies ein Analogieschluss ist, habe ich an anderer Stelle kurz dargestellt (https://www.wort-und-wissen.org/wp-content/uploads/f03.pdf).
Immunisierung gegen Kritik? Thomas Waschke schreibt weiter: „Dazu kommt noch eine weitere Asymmetrie: wenn die Entstehung einer Struktur naturalistisch erklärt wurde, kann ID immer noch sagen, dass es durchaus noch andere Strukturen gibt, deren Genese eben noch nicht erklärt werden kann.“ In meinem Genesisnet-Artikel über „nichtreduzierbare Komplexität“ habe ich dazu Stellung genommen („Nichtreduzierbare Komplexität“, https://www.genesisnet.info/pdfs/Irreduzible_Komplexitaet.pdf). Selbstverständlich muss nicht für jede Struktur nachgewiesen werden, dass eine evolutive Entstehung möglich ist. Wenn dieser Nachweis einige Male gelingt, so steigt die Plausibilität, dass dies auch bei vergleichbar komplexen Strukturen möglich ist. Es sei in aller Deutlichkeit gesagt: Nicht „erst wenn alles erklärt wurde, ist kein Designer mehr plausibel“, sondern bereits dann, wenn eines der komplexesten Designs naturalistisch erklärt würde. Das ist eine Einladung zum Falsifizieren und alles andere als eine Immunisierungsstrategie. Waschke meint dagegen. „Ein einziges Beispiel für Design würde auf der anderen Seite den Naturalismus endgültig widerlegen. Daher ist die naturalistische Position viel leichter prüfbar.“ Dass Design durch intelligente Planung entsteht, ist aber längst gezeigt, und der Naturalismus zieht sich auf ein „wissen wir noch nicht“ zurück und immunisiert damit seine Position gegen Kritik.
Weshalb ID Forschung beleben könnte. Welchen Platz im Haus der Naturwissenschaften nehmen Theorien zur Herkunft des Lebens tatsächlich ein? Bilden sie das Fundament, auf dem alles Wissen ruht oder bestimmen sie eher die Architektur des Bauwerkes? ID hat wie die Evolutionslehre die Ursprungsfrage des Lebens zum Thema. Beide Ansätze greifen auf empirisches Vorwissen zurück (z. B. vergleichende Anatomie, Biochemie, Molekularbiologie, Paläontologie etc.). Für alle Bereiche der morphologischen, physiologischen oder molekularbiologischen Analyse biologischer Phänomene (z. B. des Sehvorganges oder der Nierenfunktion) gilt, dass eine Beschreibung gelingt, „ohne dass auf nur eine evolutionsbiologische Aussage zurückgegriffen werden müsste“ (M. Gutmann: Begründungsstrukturen von Evolutionstheorien. In: U. Krohs & G. Toepfer: Philosophie der Biologie. Frankfurt/M, 2005, S. 259). Interessanterweise führt auch das Fragen nach dem „Woher“ – egal ob durch die Evolutionsbiologie oder durch ID motiviert – ständig zu neuen Fragen nach dem „Wie“ des biologischen Seins. Aus diesem Grund hat ID eine Existenzberechtigung in der Wissenschaft. Aus dem ID-Ansatz folgen Fragestellungen, die zu Wissenszuwachs führen werden, wenn man sich ihnen widmen würde.
Für die Ursprungsfrage scheint es angemessen, einen breiteren Forschungsansatz gelten zu lassen als den, der nur naturgesetzmäßig Erfassbares berücksichtigt (siehe dazu „Ist Intelligent Design wissenschaftsfeindlich?“, https://www.genesisnet.info/index.php?Sprache=de&Frage=86&FG=4).
In der Ursprungsfrage geht es darum, auf der Grundlage gegenwärtig verfügbarer Befunde einen hypothetischen vergangenen Prozess zu rekonstruieren. Ein solcher Prozess kann nur beschreibend simuliert, nicht aber direkt untersucht werden. Wer die Option ID offenhält, wird nicht nur versuchen, das Potential gesetzmäßig beschreibbarer Prozesse auszuloten, sondern auch aktiv nach Indizien für Planung (Design-Signalen) suchen. Das Mittel dazu ist Naturwissenschaft, auch wenn der Schluss auf das Wirken eines Designers über Naturwissenschaft hinausgeht. Wer diese Option ID ausschließt, versagt sich möglicherweise Erkenntnisse, die er mit dieser Option gewinnen könnte.
Weiterer Link
Antwort auf den zitierten Artikel von Martin Neukamm: Kontroverse über Wissenschaft und „Intelligent Design“, https://www.genesisnet.info/index.php?News=76.
Autor dieser News: Reinhard Junker
© 2007, http://www.genesisnet.info/schoepfung_evolution/n78.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.11.06 Aktualisierter Text über „irreduzible Komplexität“
In der Diskussion um „Intelligent Design“ (ID) (Einführung in „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1621&Sprache=de&l=1) spielt irreduzible Komplexität („irreducible complexity“) bei Lebewesen eine besondere Rolle. Ein System ist irreduzibel komplex, wenn es notwendigerweise aus mehreren fein aufeinander abgestimmten, interagierenden Teilen besteht, die für eine bestimmte Funktion benötigt werden, so dass die Entfernung eines beliebigen Teils die Funktion restlos zerstört. Aus Erfahrung ist bekannt, dass eine zweckorientierte irreduzible Komplexität bei technischen Systemen nur durch Einsatz von Know how und Planung möglich ist.
Trifft dies auch auf die Lebewesen zu? Dazu müsste zum einen gezeigt werden, dass es tatsächlich irreduzibel komplexe Systeme bei den Lebewesen gibt. Zum anderen wäre der Nachweis erforderlich, dass solche Systeme durch bloße Mechanismen nicht entstehen können. Von Kritikern wird beides bestritten. Eine Reihe von Mechanismen wurde vorgeschlagen, wie irreduzible Komplexität auf natürlichem Wege aufgebaut werden kann, ohne Planung und ohne Wirken eines Designers.
Im aktualisierten Artikel Irreduzible Komplexität (https://www.genesisnet.info/pdfs/Irreduzible_Komplexitaet.pdf, PDF-Datei, ca. 170 KB) wird gezeigt, dass die vorgebrachten Kritikpunkte das evolutionskritische Argument der irreduziblen Komplexität nicht entkräften. Der Bogen spannt sich dabei von der vermeintlich reduzierbaren Mausefalle bis zu Themen wie „Kooption“, „Funktionswechsel“, „Genduplikation“, Vorstufen des berühmten Bakterien-Rotationsmotors bis zu einem behaupteten experimentellen Nachweis der evolutiven Entstehung von irreduzibler Komplexität (zu Letzterem vgl. auch: Wissenschaftliche Kontroverse über irreduzible Komplexität, https://www.genesisnet.info/index.php?News=64).
Außerdem wird auf wissenschaftstheoretische Aspekte im Zusammenhang von irreduzibler Komplexität und „Intelligent Design“ eingegangen. Es wird gefolgert, dass das Konzept der irreduziblen Komplexität als Impulsgeber für Forschung wirken kann. Je mehr ein System erforscht ist, desto eher kann irreduzible Komplexität und das daran ansetzende evolutionskritische Argument plausibel gemacht – oder auch zerstreut – werden. Dies ist nur ein Beispiel dafür, dass der ID-Ansatz Forschung nicht hemmt, sondern unter Umständen sogar mehr anregt, als wenn offene Fragen als scheinbar evolutionstheoretisch gelöst betrachtet werden. Angesichts der in den Medien verbreiteten unzutreffenden Behauptung, der ID-Ansatz sei wissenschaftsfeindlich, kann auf diesen Sachverhalt nicht genug hingewiesen werden.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2006, http://www.genesisnet.info/schoepfung_evolution/n74.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
04.05.06 Wissenschaftliche Kontroverse über irreduzible Komplexität
In der aktuellen Diskussion um „Intelligent Design“ (ID) ist eine der Standardbehauptungen, dass es zu diesem Thema keine wissenschaftliche Diskussion gebe, weil es nichts zu diskutieren gebe. Nun ist ausgerechnet in der angesehenen Wissenschaftszeitschrift Science ein Artikel erschienen (Bridgham et al. 2006), der sich anhand naturwissenschaftlicher Befunde mit einem Hauptargument für ID auseinandersetzt, nämlich dem Konzept von „irreduzibler Komplexität“ (IC). Ein biologisches System ist irreduzibel komplex, wenn es notwendigerweise aus mehreren fein aufeinander abgestimmten, interagierenden Teilen besteht, die für eine bestimmte Funktion benötigt werden, so dass die Entfernung eines beliebigen Teils die Funktion restlos zerstört (Behe 1996; 2001). Ein solches System wird als IC-System bezeichnet. Darauf kann ein sog. IC-Argument wie folgt aufgebaut werden: Es ist nicht möglich, ein IC-System schrittweise durch ungerichtete graduelle Prozesse aufzubauen.
Die Arbeit von Bridgham et al. wurde als so bedeutsam eingestuft, dass in derselben Science-Ausgabe auch ein Kommentar dazu von Christoph Adami erschien. Nachdem dieser Kommentator die Ergebnisse von Bridgham et al. erläutert hatte, kam er paradoxerweise zum Schluss: „Consequently, whatever debate remains must be characterized as purely political.“ Sein eigener Kommentar widerlegt dies jedoch, denn er geht auf das IC-Konzept und das IC-Argument mit ausschließlich naturwissenschaftlichen Argumenten ein.
Zur Sache: Adami behauptet, die Arbeit von Bridgham et al. habe gezeigt, wie ein IC-System durch evolutive Prozesse entstehen kann, nämlich durch eine Genduplikation und wenige Punktmutationen. Das IC-Argument wäre damit widerlegt. Bevor im Folgenden darauf eingegangen wird, sei darauf hingewiesen, dass damit bestätigt wird, dass ID-Argumente widerlegbar sind, und dass dies nichts mit „Politik“, sondern nur mit „Wissenschaft“ zu tun hat.
Was haben Bridgham et al. gezeigt? Es geht um die Bindung von Hormonen an Rezeptorproteine. Solche Rezeptoren triggern Signalkaskaden in der Zelle, die durch die Bindung der Hormone an Rezeptoren in Gang gesetzt werden. Dadurch werden spezifische physiologische Vorgänge ausgelöst. Die Ergebnisse der Experimente von Bridgham können in wenigen Sätzen beschrieben werden (nach Behe 2006): Die Autoren starteten ihre Experimente mit einem Rezeptorprotein, welches die Fähigkeit besaß, mit drei verschiedenen Steroidhormonen (Aldosteron, Cortisol and 11-Deoxycorticosteron [DOC]) stark zu interagieren. Durch Punktmutationen interagierte dieses Protein viel schwächer mit diesen drei Steroiden. Eine bereits vorhandene Fähigkeit wurde also abgeschwächt. Soweit der nackte experimentelle Befund. Er belegt offenkundig in keiner Weise die Entstehung von IC und es stellt sich angesichts dieses Befundes die Frage, weshalb die Autoren und der Kommentator die evolutive Entstehung eines IC-Systems gezeigt haben wollen. Dazu muss etwas ausgeholt werden.
Die Autoren rekonstruierten einen hypothetischen Vorläufer-Rezeptor (VR), von dem zwei heute vorkommende Rezeptor-Typen evolviert sein sollen, nämlich der Glucocorticoid-Rezeptor (GR) und der Mineralocorticoid-Rezeptor (MR). MR und GR üben verschiedene Signalfunktionen aus (Bridgham et al. 2006, 97). Die heutigen Rezeptoren sollen durch eine Genduplikation (ein hypothetisches Ereignis; das Vorkommen von Genduplikationen ist aber grundsätzlich experimentell belegt) und durch Punktmutationen aus dem VR entstanden sein. Der künstlich hergestellte hypothetische VR kann wie das heutige MR die strukturell ähnlichen Hormone Aldosteron, Cortisol and DOC (s.o.) stark binden (vgl. Abb. 1). Zur Zeit, als der VR (auf unbekannte Weise) entstand (nämlich nach evolutionstheoretischen Vorstellungen vor 450 Millionen Jahren), gab es aber noch kein Aldosteron (dies wird indirekt durch vergleichende Studien an heutigen Organismen erschlossen und soll hier zugunsten des evolutionstheoretischen Arguments nicht problematisiert werden). Damit muss die Aldosteron-Affinität des VR damals ein Nebenprodukt der Affinität zu den anderen Steroidhormonen gewesen sein.
Abb. 1: Mutationen an Rezeptor. Dieses Bild zeigt links oben schematisch den hypothetischen Vorläufer-Rezeptor, der Aldosteron, Cortisol und DOC bindet. Die Mutation L111Q führt zum Verlust der Bindungsfähigkeit des Rezeptors (dargestellt durch Verformung der Bindungstasche). Die Mutation S106P führt dazu, dass Aldosteron und Cortisol nur noch sehr schwach gebunden werden können, während DOC unverändert gebunden werden kann. Beide Mutationen zusammen erlauben wieder eine starke Bindung von Cortisol; Aldosteron passt jedoch nach wie vor nicht mehr. Nach Adami C (2006) Reducible Complexity. Science 312, 61-63.
Durch zwei Aminosäureaustausche gelangte man nun vom VR ausgehend zum GR, der Cortisol und DOC, nicht aber Aldosteron binden kann. Treten die beiden dazu erforderlichen Mutationen nacheinander auf, so führt dies entweder über eine Zwischenstation, in der der Rezeptor die drei Hormone gar nicht oder nur ganz schwach binden kann oder auf der er DOC und die anderen beiden Hormone nur schwach binden kann (Abb. 1 rechts oben bzw. links unten). Die jeweils zweite Mutation führte dann zum heutigen GR, das die (in diesem Szenario erneute!) Fähigkeit besitzt, Cortisol (stärker) zu binden (während Aldosteron nach wie vor nicht mehr gebunden wird) (in Abb. 1 die Schritte nach rechts unten). Das andere Duplikat des VR wurde zum heutigen MR, das wie der VR alle drei Hormone binden kann.
Bridgham et al. heben hervor, dass die Aldosteron-Sensitivität des MR schon in seinem VR-Vorläufer vorhanden war, bevor es das Aldosteron überhaupt gab (s. o.). Der VR und seine Nachfolger waren also präadaptiert für eine Sensitivität für Aldosteron. Dieser (nur indirekt erschlossene!) Vorgang ist es, der von Bridgham et al. und Adami als Beleg für die Evolution einer IC-Struktur gewertet wird: „This evolutionary scenario – recruiting an ancient receptor into partnership with a novel ligand …“ (Bridgham et al. 2006, 100).
Aus den vorstehenden Beschreibungen geht hervor (wie oben kurz dargestellt), dass die einzigen Vorgänge dieses Szenarios, die experimentell belegt sind, die Folgen der beiden beschriebenen Punktmutationen sind (wie in Abb. 1 dargestellt).
Gegen die Interpretation dieser Vorgänge als Belege für die evolutive Entstehung von IC gibt es schwerwiegende Einwände (die meisten Einwände im Folgenden nach Behe 2006):
- Selbst wenn der Erwerb der Affinität zu Aldosteron der VR (bzw. späteren MR) ein zufällig passendes Nebenprodukt war (wie oben beschrieben), handelt es sich doch ein ein strukturell ähnliches Molekül im Vergleich zu Cortisol und DOC.
- Es handelt sich nach der Definition von Behe (s. o.) gar nicht um ein IC-System, da es nur aus einem einzigen Protein (Rezeptor) und seinem Liganden (dem Hormon) besteht. Das Rezeptor-Hormon-Paar ist dabei nur ein kleiner Teil einer Signalübertragskaskade, die andere komplexe physiologische Prozesse reguliert. Rezeptor-Liganden-Paare besitzen für sich alleine selektierbare Funktionen (vgl. dazu auch Nelson 2006). Es wird also nur ein für sich alleine funktionsloser Teil eines IC-Systems betrachtet.
- Im Experiment wurden weder neue Komponenten hinzugefügt noch alte entfernt.
- Es wurde die schon vorhandene Fähigkeit eines Proteins, bestimmte (ähnliche) Moleküle zu binden, lediglich geschwächt. Die Experimente starteten mit einem Protein,welches mehrere strukturell sehr ähnliche Steroide stark binden kann und hatten als Ergebnis ein Protein, das zwei der Steroide höchstens 10malschwächer binden konnte.
Stephen C. Meyer vom Discovery Institut (Seattle) kommentiert diese Ergebnisse wie folgt: Wenn diese Experimente das Beste sind, das Michael Behes Kritiker nach zehn Jahren Widerlegungsbemühungen vorweisen können, dann ist der Neodarwinismus in großen Schwierigkeiten. Ob Science die Größe haben wird, eine Stellungnahme von Michael Behe zu publizieren?
Quellen
Adami C (2006) Reducible Complexity. Science 312, 61-63.
Behe MJ (1996) Darwin’s Black Box: the Biochemical Challenge to Evolution. New York.
Behe MJ (2001) Reply to my critics: A response to reviews of Darwin’s Black Box: the Biochemical Challenge to Evolution. Biol. Philos. 16, 685-709.
Behe M (2006) The Lamest Attempt Yet to Answer the Challenge Irreducible Complexity Poses for Darwinian Evolution. www.idthefuture.com/2006/04/the_lamest_attempt_yet_to_answ.html
Bridgham JT, Carroll SM & Thornton JW (2006) Evolution of Hormone-Receptor Complexity by Molecular Exploitation. Science 312, 97-100.
Nelson P (2006) How to Explain Irreducible Complexity – A Lab Manual (Step One; to be continued). www.idthefuture.com/2006/04/how_to_explain_irreducible_com_1.html
Meyer SC (2006) Responds to Research on Irreducible Complexity. www.discovery.org/scripts/viewDB/index.php?command=view&id=3406
Autor dieser News: Reinhard Junker
© 2006, http://www.genesisnet.info/schoepfung_evolution/n64.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
29.09.05 Sechs aktuelle Fragen zu Schöpfung und Intelligent Design beantwortet
Anlässlicher zahlreicher kritischer Artikel über Schöpfungslehre und Intelligent Design, die in jüngster Zeit veröffentlicht wurden, wurden weitere Antworten auf oft gestellte Fragen unter der Rubrik „Fragen und Antworten“ zusammengestellt. Es handelt sich um folgende Themen:
- Worin unterscheiden sich Schöpfungs- und Evolutionslehre?, https://www.genesisnet.info/index.php?Sprache=de&Frage=75&FG=3
- Widersprechen sich Schöpfung und Evolution?, https://www.genesisnet.info/index.php?Sprache=de&Frage=76&FG=5
- Ist „Intelligent Design“ wissenschaftlich testbar und widerlegbar?, https://www.genesisnet.info/index.php?Sprache=de&Frage=77&FG=4
- Fördert der Ansatz der Schöpfungslehre und des „Intelligent Design“ Forschung?, https://www.genesisnet.info/index.php?Sprache=de&Frage=78&FG=1
- Ist der Bezug auf einen Schöpfer in der Wissenschaft erlaubt?, https://www.genesisnet.info/index.php?Sprache=de&Frage=79&FG=1
- Worin unterscheiden sich Kreationismus und „Intelligent Design“?, https://www.genesisnet.info/index.php?Sprache=de&Frage=80&FG=1
Eine weitere Liste solcher Fragen ist unter Acht neue Fragen zur aktuellen Diskussion um Schöpfung und Intelligent Design zusammengestellt: https://www.genesisnet.info/index.php?News=43.
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n47.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
09.09.05 Neuer Artikel über Irreduzible Komplexität
In der Diskussion um „Intelligent Design“ (ID) (Einführung in „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1621&Sprache=de&l=1) spielt irreduzible Komplexität („irreducible complexity“) bei Lebewesen eine besondere Rolle. Jedes Lebewesen enthält irreduzibel komplexe Teilsysteme. Ein System ist irreduzibel komplex, wenn es notwendigerweise aus mehreren fein aufeinander abgestimmten, interagierenden Teilen besteht, die für eine bestimmte Funktion benötigt werden, so dass die Entfernung eines beliebigen Teils die Funktion restlos zerstört. Die evolutive Entstehung solcher Systeme auf der Basis ungerichteter Prozesse ist unbekannt. Aus Erfahrung ist jedoch bekannt, dass irreduzible Komplexität durch Einsatz von Know how und Steuerung von außen möglich ist.
Im neuen Artikel zu diesem Thema „Irreduzible Komplexität“ (https://www.genesisnet.info/index.php?Artikel=1624&Sprache=de&l=2) wird genauer erläutert, was irreduzible Komplexität (IC) ist und gezeigt, weshalb die Entstehung von IC durch ungerichtete evolutive Prozesse derzeit unbekannt ist. Außerdem wird gezeigt, weshalb IC auch positiv als Argument für einen intelligenten Urheber gewertet werden kann: Irreduzible Komplexität kann als tertium comparationis zwischen technischen und lebendigen Systemen dienen, so dass in einem Analogieschluss beim Nachweis von irreduzibler Komplexität auf einen Urheber geschlossen werden kann.
Gegen das evolutionskritische IC-Argument wurde eine Reihe von Kritikpunkten vorgebracht; im Artikel wird gezeigt, dass diese nicht stichhaltig sind. Außerdem wird auf wissenschaftstheoretische Aspekte im Zusammenhang von IC und „Intelligent Design“ eingegangen. Dabei werden folgende Fragen behandelt:
- Ist IC empirisch prüfbar?
- Ist IC widerlegbar?
- Kann eine IC-Struktur sprunghaft entstehen?
- Verhindert der Verweis auf IC Forschung?
- Beruht das IC-Argument auf Nichtwissen?
- Kann bewiesen werden, dass IC nicht evolvierbar ist?
- Fördert das IC-Konzept Forschung?
- Ist der Bezug auf einen Designer erlaubt?
Es wird gefolgert, dass das IC-Konzept als Impulsgeber für Forschung wirken kann: Das IC-Konzept regt an, genau hinzuschauen. Nur ein detailliertes Studium eines IC-verdächtigen Systems kann zeigen, ob es wirklich IC ist, und nur die weiter verbesserte Kenntnis der Evolutionsmechanismen kann die Plausibilität einer evolutiven Entstehung einer IC-Struktur begründen, erhöhen oder verringern. Das Ergebnis steht dabei nicht von vornherein fest.
Eine ausführlichere Version des Artikels kann auch als PDF (Irreduzible Komplexität, https://www.genesisnet.info/pdfs/Irreduzible_Komplexitaet.pdf) heruntergeladen werden.
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n45.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
11.08.05 Acht neue Fragen zur aktuellen Diskussion um Schöpfung und Intelligent Design
Diesen Sommer erschienen in der Tages- und Wochenpresse fast pausenlos kritische Artikel über Schöpfungslehre und Intelligent Design. Auf viele Einwände gegen diese Denkrichtungen gibt es längst publizierte Antworten. Diese wurden den Lesern jedoch fast immer vorenthalten. Die Autoren waren daher entweder nicht informiert und haben schlecht oder gar nicht recherchiert (jedenfalls nicht aus erster Hand) oder haben bekannte Informationen unterschlagen. Ein krasses Beispiel wird in „Desinformation über das evolutionskritische Lehrbuch“ (https://www.genesisnet.info/index.php?News=41) geschildert.
Einige Standardkritikpunkte stehen mittlerweile unter „Fragen und Antworten“ auf Genesisnet, im Einzelnen:
Ist die Evolutionslehre falsifizierbar?, https://www.genesisnet.info/index.php?Sprache=de&Frage=71&FG=101
Sind in der Schöpfungslehre konkrete, falsifizierbare Vorhersagen möglich?, https://www.genesisnet.info/index.php?Sprache=de&Frage=69&FG=1
Kann man unter der Vorgabe von Schöpfung ergebnisoffene Wissenschaft betreiben?, https://www.genesisnet.info/index.php?Sprache=de&Frage=67&FG=1
Ist die Schöpfungslehre forschungsfeindlich, weil alles mit dem Handeln eines Schöpfers erklärt wird?, https://www.genesisnet.info/index.php?Sprache=de&Frage=68&FG=1
Ist Gott als Schöpfer ein „Lückenbüßer“ für Unverstandenes?, https://www.genesisnet.info/index.php?Sprache=de&Frage=73&FG=1
Kann man Wissenschaft betreiben, wenn eine Schöpfung dabei nicht in Frage gestellt wird?, https://www.genesisnet.info/index.php?Sprache=de&Frage=70&FG=1
Ist das Fehlen eines Mechanismus ein Argument gegen „Intelligent Design“?, https://www.genesisnet.info/index.php?Sprache=de&Frage=72&FG=4
Außerdem wurde der Expertenteil des Artikels „Argumente gegen Design“ (https://www.genesisnet.info/index.php?Artikel=1641&Sprache=de&l=2) um zwei Abschnitte ergänzt: „Design-Fehler, Forschungsanreize und Testbarkeit“ sowie „Design und Constraints“.
Da in der Auseinandersetzung um Schöpfung und Evolution häufig beteuert wird, man wolle niemandes Glauben angreifen, wird in „Welches Gottesbild verträgt sich mit einer naturalistischen Evolution?“ (https://www.genesisnet.info/index.php?Sprache=de&Frage=74&FG=5) gezeigt, dass dies letztlich nicht den Tatsachen entspricht. Anhand einer Reihe von aktuellen Zitaten von drei deutschen Autoren wird gezeigt, dass es sehr wohl darum geht, schöpfungsgläubige Menschen durch Vermittlung eines konsequent evolutionistischen Weltbildes von der Unhaltbarkeit ihrer Glaubensbasis zu überzeugen. Das gehört selbstverständlich zur Ausübung der Meinungsfreiheit in unserem Land. Allerdings kann man sich des Eindrucks nicht ganz erwehren, dass diese Bemühungen den missionarischen Bestrebungen nicht unähnlich sind, welche die meisten Religionen auszeichnen.
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n43.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
18.07.05 Neue Texte zum Thema Schöpfung, Grundtypen und Wissenschaft
Der biblische Schöpfungsbericht und andere biblische Schilderungen über Gott als Schöpfer und über sein Handeln in der Geschichte motivieren einige Postulate, die der Grundtypenbiologie (Heutige Grundtypen, https://www.genesisnet.info/index.php?Artikel=1241&Sprache=de&l=1) und der Design-Theorie (Einführung in „Intelligent-Design“, https://www.genesisnet.info/index.php?Artikel=1621&Sprache=de&l=1) zugrundegelegt werden können. Aus diesen Postulaten kann eine Reihe von Hypothesen abgeleitet werden, die wissenschaftlich prüfbar sind und Forschung anregen. In dem neuen Artikel „Biblische Schöpfungslehre und Grundtypenbiologie“ (https://www.genesisnet.info/index.php?Artikel=1221&Sprache=de&l=1) werden folgende Postulate zusammengestellt und begründet:
- Alle Lebewesen waren ursprünglich optimal konstruiert.
- Alle Lebewesen starteten ihre Existenz als polyvalente, abgegrenzte Grundtypen.
- Die Lebewesen sind nur im Rahmen der vorgegebenen Polyvalenz zu Variation fähig.
- Es gibt in großem Ausmaß inkongruente Merkmalskombinationen bei den verschiedenen Grundtypen (modulares Baukastensystem)
- Alle Grundtypen sind geologisch gesehen gleichzeitig entstanden.
Aus diesen Postulaten können testbare und falsifizierbare Hypothesen abgeleitet werden – acht Hypothesen werden zusammengestellt.
Ein weiterer neuer Artikel „Schöpfung und Wissenschaft“ (https://www.genesisnet.info/index.php?Artikel=422&Sprache=de&l=1) befasst sich mit einigen grundsätzlichen Fragen zum Verhältnis von Schöpfungsparadigma und empirischer Wissenschaft. Es wird gezeigt, wie unter der Vorgabe von Schöpfung fruchtbare, erkenntnisfördernde Wissenschaft betrieben werden kann. Wichtig ist die Unterscheidung zwischen dem Rahmenparadigma einerseits und konkreten Hypothesen innerhalb von diesem Rahmen andererseits. Mit Hypothesen, die im Rahmen des Schöpfungsparadigmas aufgestellt werden, kann man genauso verfahren wie mit evolutionären Hypothesen: sie sind prüfbar, widerlegbar, regen Forschung an und führen zu neuen Erkenntnissen.
In diesem Artikel wird auch eine Reihe von Argumenten diskutiert, mit denen einer Wissenschaft auf der Basis von Schöpfung das Existenzrecht bestritten wird, und es wird gezeigt, weshalb diese Argumente nicht stichhaltig und daher zurückzuweisen sind. Zu diesem Thema gibt es sowohl einen Interessierten- als auch einen Expertentext.
Schließlich wurde mittlerweile der Artikel „Wissenschaft und Weltanschauung“ (https://www.genesisnet.info/index.php?Artikel=40463&Sprache=de&l=1) ausgebaut und ist nun auch als Expertentext verfügbar. Es wird dargelegt, dass in der Wissenschaft, insbesondere in der Ursprungsfrage unvermeidlich weltanschauliche Vorgaben den konkreten wissenschaftlichen Arbeiten zugrunde liegen. Neu sind darin die Abschnitte „Ist der Naturalismus falsifizierbar?“ und „Wird der Naturalismus methodologisch erzwungen?“. Es wird die Schlussfolgerung gezogen, dass der Naturalismus eine weltanschauliche Vorgabe über die Welt ist, die weder empirisch noch methodisch erzwungen wird und die nicht falsifiziert werden kann.
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n42.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
25.06.05 Neue Artikel zur Schöpfungslehre und zur Grundtypenbiologie
In zwei neuen Artikeln geht es um biblische Grundlagen einer schöpfungsorientierten Wissenschaft und um Kritik an der Grundtypenbiologie. Im Artikel „Biblische Grundlagen der Schöpfungslehre“ (https://www.genesisnet.info/index.php?Artikel=421&Sprache=de&l=1) sind einige grundlegende Aussagen der Bibel zum Thema „Schöpfung“ zusammengestellt. Daraus werden einige allgemeine Folgerungen für die bibelorientierte Rekonstruktion der Geschichte des Lebens gezogen.
Im Artikel „Kritik an der Grundtypenbiologie“ (https://www.genesisnet.info/index.php?Artikel=1246&Sprache=de&l=1)wird Kritik vorgestellt, die an der Grundtypenbiologie geübt wurde. Der Ansatz der Grundtypenbiologie wurde bisher von der Fachwelt nur wenig beachtet. Hier und da wurden in den letzten Jahren aber vermehrt Kritikpunkte geäußert. Manchmal zeigte sich, dass der Kritiker mit den Inhalten der Grundtypenbiologie praktisch überhaupt nicht vertraut war (ein Beispiel schildert „Grundtypenmodell der Schöpfungslehre widerlegt?“, https://www.genesisnet.info/index.php?News=23). Es wurde aber auch seriösere Kritik geäußert. Auf diese wird in diesem neuen Artikel ausführlich eingegangen. Es wird gezeigt, dass entgegen der Ansicht der Kritiker die Ansätze der Grundtypenbiologie prüfbar sind und Forschung anregen. Vermeintliche Widersprüche innerhalb der Grundtypenbiologie werden aufgelöst. Im Expertenteil („Kritik an der Grundtypenbiologie“, https://www.genesisnet.info/index.php?Artikel=1246&Sprache=de&l=2) wird dabei auch auf die Frage eingegangen, inwiefern Tests des Grundtypenmodells auch Tests auf Schöpfung sind.
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n39.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
05.05.04 Lesenswerte Sammlung von 15 persönlichen Berichten von Wissenschaftlern
Wie bringen wissenschaftlich arbeitende Christen den biblischen Glauben und das Wissen Ihres Fachgebietes zusammen? Und was führte sie dazu, sich im Rahmen der biblischen Schöpfungslehre wissenschaftlich zu engagieren? Dies schildern 15 Mitarbeiter der Studiengemeinschaft Wort und Wissen in persönlichen Zeugnissen erfrischend offen in einem sehr lesenswerten und kurzweilig geschriebenen Buch: Mit Kopf und Herz. Bekenntnisse zum Gott der Bibel im Zeitalter der Wissenschaft; hgg. von Henrik Ullrich (Pb., 158 S., 44 Abb., nur 5,95 Euro / 11,90 sfr). Nähere Informationen, eine Inhaltsübersicht und eine Bestellmöglichkeit gibt es unter: https://www.wort-und-wissen.org/produkt/mit-kopf-und-herz/.
Autor dieser News: Reinhard Junker
© 2004, http://www.genesisnet.info/schoepfung_evolution/n20.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
26.03.04 Drei neue Artikel zu „Intelligent Design“ online
Der Grundgedanke des „Intelligent Design“ (ID) ist, dass man an Strukturen der Lebewesen Eigenschaften („Signale“) erkennen könne, die auf das Wirken eines intelligenten, willensbegabten Urhebers (Designer, Schöpfer) hinweisen und andere Möglichkeiten ihrer Herkunft ausschließen (insbesondere die Möglichkeit einer Evolution). Dieses Thema ist seit einigen Jahren Gegenstand intensiver Diskussionen, vor allem im Internet. Drei neue Beiträge befassen sich mit dieser spannenden Thematik. In „Einführung in Intelligent-Design“ (https://www.genesisnet.info/index.php?Artikel=1621&Sprache=de&l=1) wird das ID-Konzept erklärt, während in „Kontroverse um Intelligent-Design“ (https://www.genesisnet.info/index.php?Artikel=1622&Sprache=de&l=1) Kritik am ID-Konzept erläutert und bewertet wird.
Es werden auch Argumente dagegen vorgebracht, dass ein intelligenter Urheber die Lebewesen erschaffen haben könnte. Der Artikel „Argumente gegen Design“ (https://www.genesisnet.info/index.php?Artikel=1641&Sprache=de&l=1) befasst sich mit diesen Argumenten und setzt sich kritisch mit ihnen auseinander. Für an Details Interessierte werden zum Thema „ID“ und „Argumente gegen Design“ PDF-Dateien angeboten.
Autor dieser News: Reinhard Junker
© 2004, http://www.genesisnet.info/schoepfung_evolution/n16.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
12.01.04 Artikel über polyvalente Stammformen von Grundtypen online
In der Grundtypenbiologie (https://www.genesisnet.info/index.php?Artikel=1200&Sprache=de&l=1) der Schöpfungslehre geht es neben der Frage nach der Abgrenzbarkeit von Grundtypen auch um die Variabilität innerhalb von Grundtypen. Als „Pool“ für die Vielfalt der Grundtypen wird eine genetische Vielseitigkeit (Polyvalenz) der Ursprungsformen angenommen.
Der neue Artikel „Genetisch polyvalente Stammformen von Grundtypen“ (https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1) erklärt, was unter Polyvalenz von Grundtypen zu verstehen ist und welche Indizien es für eine ursprüngliche Polyvalenz gibt. Vielfalt innerhalb von Grundtypen entstand demnach nicht in erster Linie durch Mutationen, sondern auf der Basis einer vorgegebenen programmierten Variabilität.
Autor dieser News: Torsten Poitzsch
© 2004, http://www.genesisnet.info/schoepfung_evolution/n12.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
10.01.04 Neuer Artikel über Mosaikformen und Baukastensysteme
Die Merkmale verschiedener „Grundtypen“ (https://www.genesisnet.info/index.php?Artikel=1240&Sprache=de&l=1) und höherer systematischer Gruppen sind oft so verteilt, dass netzartige – weniger baumförmige – Beziehungen resultieren. Die verschiedenen Grundtypen scheinen nach einem „Baukastensystem“ konstruiert zu sein.
Der neue Artikel „Mosaikformen als Grundtypen und Baukastensysteme“ (https://www.genesisnet.info/index.php?Artikel=1261&Sprache=de&l=1) erklärt, was unter „Mosaikformen“ zu verstehen ist und wie die Existenz von Mosaikformen im Rahmen des Grundtypmodells der Schöpfungslehre interpretierbar ist.
Autor dieser News: Torsten Poitzsch
© 2004, http://www.genesisnet.info/schoepfung_evolution/n11.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/