
(Paläo-)Anthropologie: Herkunft des Menschen
15.09.23 Hatte Homo erectus eine menschliche Kindheitsdauer?
Ein Merkmal, das Menschenaffen und Menschen nach typischer evolutionärer Lehrmeinung unterscheidet, ist die Dauer der Kindheit, die man Individualentwicklung nennt. Allerdings wird immer wieder behauptet, dass Homo erectus (der „aufrechte Mensch“) hinsichtlich der Individualentwicklung eine Übergangsform von großaffenähnlichen Vorfahren zu heutigen Menschen darstellen soll. Das Ergebnis einer Reihe von Methoden, die zur Altersbestimmung eingesetzt werden, zeigt aber: Homo erectus hatte wahrscheinlich eine menschliche Kindheitsdauer, wenn man ein Alter von ca. 10 Jahren ansetzt.
Einleitung
In der Evolutionsbiologie wird allgemein angenommen, dass sich Schimpanse und Mensch aus gemeinsamen Vorfahren vor ca. 7 Mio. radiometrischen Jahren (rJ) entwickelt haben. Der Weg zum Menschen soll über großaffenähnliche Australomorphen (z. B. Australopithecus) gelaufen sein und dann über Homo erectus zum modernen Menschen geführt haben. Ein Argument für diese Perspektive lautet, dass Homo erectus eine verkürzte Kindheitsdauer (Individualentwicklung) gehabt haben soll, die sich als eine Übergangsform zwischen heutigen Menschenaffen und Menschen eignet (vgl. Collard & Wood 2015). Andere Merkmale wie Körpergröße, Körperform, Fortbewegungsweise und Gebissmerkmale von Homo erectus seien hingegen typisch menschlich gewesen (vgl. ebd.).
Bei der These einer verkürzten Kindheit wird häufig auf das vollständigste Homo-erectus-Fossil namens Turkana Boy aus Kenia verwiesen. Ein Special Paper von Benjamin Scholl mit dem Titel „Hatte Homo erectus eine verkürzte Kindheit ähnlich derjenigen der Menschenaffen?“ (http://www.wort-und-wissen.de/artikel/homo-erectus-individialentwicklung/) setzt sich kritisch mit den zugrundeliegenden Vorannahmen einer vermeintlich nichtmenschlichen Individualentwicklung des Turkana Boy auseinander. Die Ergebnisse werden hier kurz zusammengefasst.
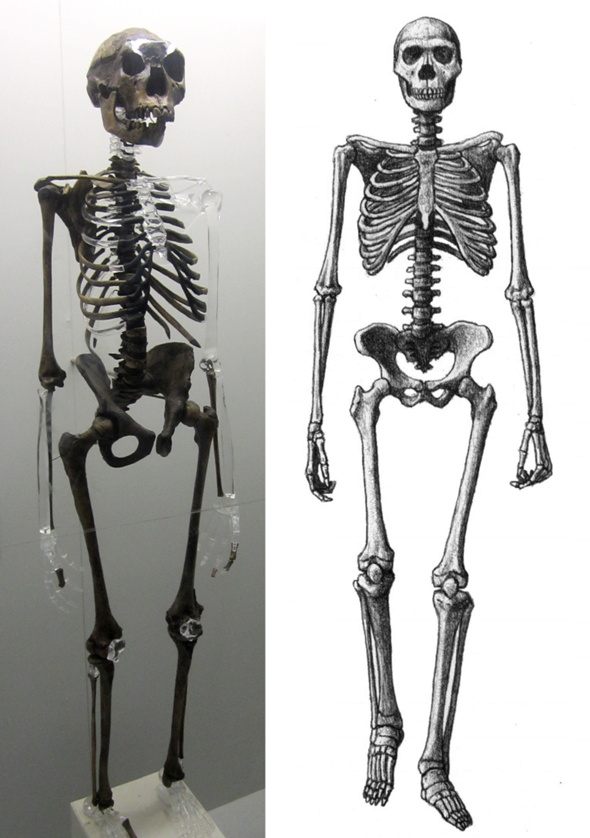
Abb. 1 Der Turkana Boy (Links: Skelett, Rechts: Skizze) besitzt insgesamt ein menschenähnliches Skelett. (Wikimedia: Rept0n1x, CC BY-SA 3.0; Mauricio Antón, published with Alan Turner: Africa – the Evolution of a Continent and its Large Mammal Fauna, CC BY 4.0)
Verschiedene Rekonstruktionsverfahren
Graves und Kollegen (2010, Tab. 1) haben einen Überblick über bisherige Studien zur Rekonstruktion des Sterbealters des Turkana Boy vorgelegt. Dabei kommt je nach Art, die als Vergleich verwendet wird, und je nach Kriterium zur Altersbestimmung ein ganz anderes Alter für den Turkana Boy heraus. Ein geringes Sterbealter trotz relativ großer Körpergröße des Turkana Boy wird in der Regel als Argument dafür verwendet, dass er eine affenähnliche Kindheitsdauer hatte. Daher sind gerade die Studien problematisch, in denen dem Turkana Boy ein niedriges Sterbealter zugemessen wird. Sieben Studien ergaben ein Sterbealter ca. 10,5 bis 15 Jahren, unterschieden sich aber im Teil erheblich.
Vier Studien kamen aber auf ein geringes Sterbealter des Turkana Boy (ca. 7,5 bis 9,9 Jahre). Sie setzen allerdings nicht ausschließlich heutige Menschen als Vergleichsart zur Altersrekonstruktion ein, sondern auch Großaffen oder Australomorphen. Da Homo erectus aus Grundtyp-Perspektive (vgl. Brandt 2017) ein echter von Gott erschaffener Mensch war (der von Adam und Eva abstammt), sind diese Rekonstruktionen außerhalb des evolutionären Denkrahmens problematisch. Da zwei dieser Studien neben Affen auch Menschen als Vergleichsarten verwendeten, sollen sie im Folgenden genauer betrachtet werden.
Die unterschiedlichen rekonstruierten Sterbealter für Menschen unterscheiden sich aufgrund verschiedener Kriterien zur Altersbestimmung. Dies liegt daran, dass der Zustand des Skeletts und der Zähne jeweils unterschiedliche Alter nahelegen können. Doch zwei bis drei Jahre Unterschied in der Individualentwicklung der Zähne und des Skeletts sind auch bei heutigen Menschen nicht ungewöhnlich (Graves et al. 2010; vgl. Dean & Smith 2009, Tab. 10.3), weil die Entwicklungsgeschwindigkeiten verschiedener Körperbereiche unterschiedlich schnell sein können (Dean 2016; vgl. Dean & Liversidge 2015).
Altersrekonstruktion mithilfe des Gebisses
Heutige Menschen und auch Schimpansen haben eine große individuelle Vielfalt bei der Entwicklung von Zähnen im Kindheitsverlauf, weil das Zahnwachstum ein ziemlich dynamischer Prozess ist (vgl. Smith et al. 2010). Dies wird von Wissenschaftlern, die das Alter menschlicher oder affenähnlicher Fossilien bestimmen wollen, oft nicht angemessen berücksichtigt, da sich diese oft zu stark an vereinfachten Faustregeln orientieren. Zum Beispiel wird häufiger behauptet, der Zahndurchbruch des 1. Backenzahns (M1) würde bei Schimpansen mit 4,1 Jahren erfolgen (ebd.). Studien zeigen aber, dass bei Schimpansen der 1. Backenzahn zwischen ca. 2,1 und 4,4 Jahren durchbricht (sichtbar wird), während dies bei heutigen Menschen zwischen ca. 3,7 und 7,8 Jahren geschehen kann (Ekstrand et al. 2003; Smith et al. 2010; Machanda et al. 2015; Rozzi 2016; Chaitanya 2018). Findet man nun ein Fossil eines angeblichen „Vormenschen“ mit einem solchen gerade durchgebrochenen Backenzahn, kann man also nicht ableiten, dass das Individuum mit genau 4,1 Jahren gestorben sei – selbst wenn man sicher sein könnte, dass es ein Großaffe oder ein Mensch war. Auch beim Durchbruchsalter anderer Zähne gibt es gewaltige Schwankungen und immer auch einen Bereich, in dem sich Schimpansen und Menschen überlappen. Die von Scholl (2023, Tab. 2) zusammengestellten Zahndurchbruchsalter bei modernen Menschen sprechen im Vergleich mit dem Zahnzustand beim Turkana Boy (alle Zähne außer Weisheitszähne und obere Eckzähne durchgebrochen) für ein Alter des Turkana Boy zwischen ca. 7,3 bis 12,9 Jahren.
Auch anhand der mikroskopisch kleinen Zahnwachstumslinien auf den Zähnen kann man ein Alter abschätzen (ähnlich wie bei Baumringen). Allerdings ist ihre Wachstumsgeschwindigkeit nicht immer gleich, sondern unterscheidet sich zwischen ca. 6 bis 12 Tagen je nach Population und Zahntyp (vgl. Reid & Dean 2006, Tab. 2; Hu et al. 2011; Tam et al. 2017). Legt man diese jeweils minimale bzw. maximale menschliche Bildungsdauer der Zahnwachstumslinien zugrunde und korrigiert die methodischen Fehler in der Rechnung von Dean & Smith (2009), ist es möglich, das Zahn-Alter des Turkana Boy unter modern-menschlichen Bedingungen auf ca. 7,3 bis 10,1 Jahre zu rekonstruieren (vgl. Scholl 2023, Anhang Tab. 4).
Betrachtet man den konkreten Bildungszustand der Zahnkronen und der Zahnwurzeln des Turkana Boy und legt man heutige menschliche Entwicklungszeiten zugrunde, könnte der Turkana Boy minimal 7,5 bis 9,4 Jahre und maximal 11,8 bis 14,4 Jahre alt gewesen sein (vgl. Scholl 2023, Anhang Tab. 3). Insgesamt weist der Gesamteindruck des Gebisses – abgesehen von den Zahnwachstumslinien (s. o.) – auf ein Alter von ca. 10 bis 11 Jahren hin (nach Dean & Smith 2009).
So kommen auch Dean & Liversidge (2015; vgl. Dean 2016) bei den Vor- und Backenzähnen zu dem Schluss, dass der Turkana Boy ein frühes, aber menschliches Bildungsmuster der Zähne hatte. Auch die Bildungsdauer der vorderen Zähne kann im frühen menschlichen Bereich gelegen haben, wenn man das gesamte menschliche Merkmalsspektrum beim Zahnwachstum betrachtet (vgl. Scholl 2023, Abb. 2).
Übrigens gibt es nicht nur bei Menschen, sondern auch bei heutigen Schimpansen deutliche Unterschiede bei Zahndurchbruch, Bildungsdauer von Zahnwachstumslinien sowie von Zahnkrone und -wurzel (vgl. Smith et al. 2010), was eine konkrete Altersrekonstruktion auch bei großaffenähnlichen Australomorphen erschwert.
Altersbestimmung und Körperwachstum
Letztlich herrscht auch bei der Bestimmung des Skelettalters eine Unsicherheit bei einigen Wissenschaftlern (vgl. Antón & Snodgrass 2012), wie menschenähnlich der Turkana Boy diesbezüglich war. Dies sollte aber nicht überraschen, da auch das Körperwachstum, die maximale Erwachsenengröße und auch das Eintreten der Geschlechtsreife bei Schimpansen und bei heutigen Menschen extrem schwankt. Ursachen sind zum Beispiel die Herkunft und unterschiedliche Lebensbedingungen (vgl. Marson et al. 1991; Hamada et al. 1996; Ji 2001; Ilieva et al. 2002; Smith et al. 2010; Antón & Snodgrass 2012; Brandt 2015, 206; Song et al. 2016; Walker et al. 2018). Dies kann ein Vergleich aus der Kriminalistik verdeutlichen: Wenn beispielsweise bei einem Skelettfund eines modernen Menschen die Herkunft nicht klar wäre, könnte man allein aufgrund der durchschnittlichen Körpergröße entweder einen 13 Jahre alten Jungen aus den Niederlanden oder einen 19 Jahre alten Jungen von den Solomon Islands vor sich haben (vgl. Rodriguez-Martinez et al. 2020).
Laron (2010) stellte außerdem fest, dass bei Jungen in der Pubertät das Skelettalter im Durchschnitt um 9 Monate zu jung bestimmt worden war. Bei Jungen mit verzögerter Pubertät lag das geschätzte Alter des Skeletts sogar im Durchschnitt 2,85 Jahre niedriger als ihr tatsächliches Alter (ebd.).
Ebenso gibt es beim Wachstum der Epiphysen, also dem Zuwachsen der Wachstumsfugen in der Nähe der Knochenenden, große individuelle Unterschiede und auch Überlappungsbereiche bei Schimpansen und modernen Menschen (vgl. Graves et al. 2010; Dean 2016). Hinzu kommt der schlecht erhaltene Zustand des körperfernen Oberarmknochens (vgl. Ruff & Burgess 2015). Die Angabe von Dean (2016), dass der Turkana Boy mit nur teilweise geschlossener Epiphyse des körperfernen Oberarmknochens und der Beckenknochen höchstens 11 bis 14 Jahre alt gewesen sei, muss daher mit Vorsicht betrachtet werden.
Auch die Rekonstruktionsversuche zur Körpergröße des Turkana Boy zum Sterbezeitpunkt auf der Grundlage der Skelettfunde schwanken zwischen ca. 141 und 169 cm – liegen in der Mehrheit aber bei ca. 160 cm (Graves et al. 2010). Nimmt man heutige Jungen ohne extreme Wachstumskurven aus Deutschland bzw. Mosambik als Vergleichsgrundlage (Robert Koch-Institut 2011; FANTA 2018) hätte der Turkana Boy selbst bei vorsichtigen Größenschätzungen und unter Ausschluss von Erkrankungen zwischen 10 und 18 Jahren alt gewesen sein können.
Diese Beispiele verdeutlichen, wie schwierig es ist, ein konkretes Sterbealter beim Turkana Boy abzuleiten. Daher ist bei Schätzungen von Größe und Alter des ausgewachsenen Zustandes beim Turkana Boy, wie beispielsweise bei Graves et al. (2010), große Zurückhaltung geboten.
Fazit
Das Sterbealter des Turkana Boy ist aufgrund vieler unsicherer Faktoren nicht eindeutig bestimmbar. Die Bestimmungsmerkmale an Gebiss und Körperskelett bei heutigen Schimpansen und Menschen weisen in der Regel eine viel größere Vielfalt und auch Überlappung auf, als dies in der Fossilienforschung oft berücksichtigt wird. Die Behauptung, dass Homo erectus keine menschliche Kindheitsentwicklung besaß, ist daher als evolutionär bedingte Spekulation zu betrachten. Davon abgesehen würde der Turkana Boy bei jedem von Scholl (2023) untersuchten Kriterium zur Altersbestimmung als voll menschlich gelten, wenn er ungefähr 10 Jahre alt gewesen sein sollte. Somit kann für Homo erectus am Beispiel des am besten erhaltenen Skeletts Turkana Boy geschlussfolgert werden, dass Homo erectus nicht nur hinsichtlich der Kriterien Körpergröße, Körperform, Fortbewegungsweise und Gebissausprägung von Collard & Wood (2015) menschlich war, sondern wahrscheinlich auch bezüglich der Individualentwicklung (zu den durchschnittlich kleineren Gehirn von Homo erectus siehe Scholl 2022).
Interessant ist auch, dass Cofran (2019) – bei aller Vorsicht bezüglich der Altersrekonstruktion bei Fossilien – zu dem Schluss kam, dass Homo erectus vor 1 MrJ (wie auch der Neandertaler in späterer Zeit) wahrscheinlich ein langsames menschenähnliches Gehirnwachstum aufwies; während es bei Australopithecus schimpansentypisch schnell war.
Literatur
Antón SC & Snodgrass JJ (2012) Origins and Evolution of Genus Homo: New Perspectives. Curr. Anthropol. 53, S479–S496, doi: 10.1086/667692.
Brandt M (2015) Wie alt ist die Menschheit? Demographie und Steinwerkzeuge mit überraschenden Befunden. 5. erw. Aufl. SCM Hänssler.
Brandt M (2017) Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special. SCM Hänssler.
Chaitanya P et al. (2018) Time and Eruption Sequence of Permanent Teeth in Hyderabad Children: A Descriptive Cross-sectional Study. Int. J. Clin. Pediatr. Dent. 11, 330–337, doi:10.5005/jp-journals-10005-1534.
Cofran Z (2019) Brain size growth in Australopithecus. J. Hum. Evol. 130, 72–72, doi: 10.1016/j.jhevol.2019.02.006.
Collard M & Wood B (2015) Defining the Genus Homo. In: Henke W, Hardt T & Tattersall I (Ed.) Handbook of Paleoanthropology. Springer‐Verlag, 2107–2144, doi: 10.1007/978-3-642-39979-4_51.
Dean MC et al. (2001) Growth processes in teeth distinguish modern humans from Homo erectus and earlier hominins. Nature 414, 628–631, https://www.nature.com/articles/414628a.
Dean MC (2016) Measures of maturation in early fossil hominins: events at the first transition from australopiths to early Homo. Phil. Trans. R. Soc. B 371, 20150234, http://dx.doi.org/10.1098/rstb.2015.0234.
Dean MC & Liversidge HM (2015) Age estimation in fossil hominins: comparing dental development in early Homo with modern humans. Ann. Hum. Biol. 42, 415–429, doi:10.3109/03014460.2015.1046488.
Ekstrand KR, Christiansen J & Christiansen MEC (2003) Time and duration of eruption of first and second permanent molars: a longitudinal investigation. Community Dent. Oral Epidemiol. 31, 344-350, doi: 10.1034/j.1600-0528.2003.00016.x.
FANTA: Food And Nutrition Technical Assistance (2018) Mozambique Growth Charts and Tables for Children and Adolescents. aufgerufen am 26.09.2022, https://www.fantaproject.org/sites/default/files/resources/Moz-Growth-TABLES-BOY-Mar2018.pdf.
Graves RR (2010) Just how strapping was KNM-WT 15000? J. Hum. Evol. 59, 542–554, https://doi.org/10.1016/j.jhevol.2010.06.007.
Hamada Y, Udono T, Teramoto M & Sugawara T (1996) The growth pattern of chimpanzees: Somatic growth and reproductive maturation in Pan troglodytes. Primates 37, 279–295, https://doi.org/10.1007/BF02381860.
Hu R, Zhao LX, Wu XZ (2011) Periodicity of Retzius lines in fossil Pongo from South China. Chin. Sci. Bull 57, 790–794, doi: 10.1007/s11434-011-4883-3.
Ilieva E, Veleganova VK & Belcheva AB (2002) Eruption of first permanent molars in 4- to 8-year-old children. Folia Med. (Plovdiv) 44, 70–73.
Ji C-Y (2001) Age at spermarche and comparison of growth and performance of pre- and post-spermarcheal Chinese Boy. J. Hum. Biol. 13, 35–43, https://doi.org/10.1002/1520-6300(200101/02)13:1<35::AID-AJHB1005>3.0.CO;2-E.
Laron Z (2010) Age at first ejaculation (spermarche) – The overlooked milestone in male development. Pediatr. Endocrinol. Rev. 7, 256–257. PMID: 20526238.
Machanda Z et al. (2015) Dental eruption in East African wild chimpanzee. J. Hum. Evol. 82, 137–144, http://dx.doi.org/10.1016/j.jhevol.2015.02.010.
Marson J, Meuris S, Cooper RW & Jouannet P (1991) Puberty in the male chimpanzee: progressive maturation of semen characteristics. Biol. Reprod. 44, 448-55, doi: 10.1095/biolreprod44.3.448.
Robert Koch-Institut (2011) Beiträge zur Gesundheitsberichterstattung des Bundes: Referenzperzentile für anthropometrische Maßzahlen und Blutdruck aus der Studie zur Gesundheit von Kindern und Jugendlichen in Deutschland (KiGGS) 2003 bis 2006. erw. Aufl., https://www.rki.de/DE/Content/Gesundheitsmonitoring/Gesundheitsberichterstattung/GBEDownloadsB/referenzperzentile/anhang.pdf?__blob=publicationFile.
Rodriguez-Martinez A et al. (2020) Height and body-mass index trajectories of school-aged children and adolescents from 1985 to 2019 in 200 countries: Pooled analysis of 2,182 population-based studies with 65 million participants. Lancet 396, 1511–1524, doi: 10.1016/S0140-6736(20)31859-6.
Rozzi FR (2016) Diversity in tooth eruption and life history in humans: illustration from a Pygmy population. Sci. Rep. 6, 27405, https://doi.org/10.1038/srep27405.
Ruff CB & Burgess ML (2015) How much more would KNM-WT 15000 have grown? J. Hum. Evol. 80, 74–82, doi: 10.1016/j.jhevol.2014.09.005.
Smith TM et al. (2010) Dental development of the Taï Forest chimpanzees revisited. J. Hum. Evol. 58, 363–373.
Scholl (2023) Hatte Homo erectus eine verkürzte Kindheit ähnlich derjenigen der Menschenaffen? W+W Special Paper B-22-3, https://www.wort-und-wissen.org/artikel/homo-erectus-individialentwicklung/.
Scholl B (2022) Homininen-Schädel: „Stolpersteine“ des Grundtypmodells? Eine schöpfungstheoretische Deutung der Funde von Dmanisi. W+W Special Paper B-22-1, https://www.wort-und-wissen.org/artikel/homininen-schaedel/.
Autor dieser News: Benjamin Scholl
© 2023, http://www.genesisnet.info/schoepfung_evolution/n320.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
15.06.23 „Syntaxähnliche“ Struktur bei Schimpansen nachgewiesen
Bisher schien es eine gut begründete Tatsache zu sein, dass wildlebende Schimpansen nicht zu Grammatik in Lautäußerungen fähig sind, weil der Nachweis fehlte, dass Rufkombinationen von Affen tatsächlich Bedeutungen kommunizieren (vgl. Scholl 2018, 2023a). Eine neue Studie von Leroux und Kollegen vom Mai 2023 liefert allerdings gute Argumente für eine „syntaxähnliche“ Struktur bei Schimpansen (Syntax = Satzlehre). Lässt sich daraus auf eine Evolution der menschlichen Sprachen aus schimpansenähnlichen Vorfahren schlussfolgern?
Der bisherige Forschungsstand zur „Schimpansen-Grammatik“
Es ist es faszinierend, dass es durch gründliche und systematische Beobachtung von Lautäußerungen bei Tieren und deren Verknüpfung mit möglichen Bedeutungen durch Menschen möglich ist, gewisse Analogien zur menschlichen Sprache zu formulieren. Doch wie weit geht diese Ähnlichkeit zwischen der akustischen Kommunikation von Schimpansen und Menschen im Detail?
In einem Online-Artikel „Beherrschen Schimpansen etwa doch Grammatik?“[1] hatte Scholl (2023a) kürzlich eine Studie von Girad-Buttoz et al. (2022) analysiert, welche sich mit einem möglichen Nachweis einer Grammatik bei Schimpansen beschäftigt. In manchen populären Medien war daraus –fälschlicherweise – auf das Vorhandensein einer Grammatik als evolutionäre Vorstufe zur menschlichen Sprachfähigkeit geschlussfolgert worden (Scholl 2023a).
Die Forscher um Girad-Buttoz hatten ca. 5000 Rufe von Schimpansen in freier Wildbahn aufgezeichnet und dabei ca. 390 Kombinationen von Einzelrufen festgestellt. Dabei wurden in sehr seltenen Fällen bis zu zehn Rufe aneinandergereiht (meist waren es nur zwei bis fünf Rufe in Folge). Anhand von statistischen Analysen hatten die Autoren aufgezeigt, dass manche Rufe deutlich häufiger am Anfang, am Ende oder in Kombination mit anderen Rufen bei Zweier- und Dreier-Rufeinheiten verwendet wurden. Im besten Fall könnte man so zwanzig einfache grammatikalische Regeln zum Bilden von Rufkombinationen bei Schimpansen vermuten – im Gegensatz zu mindestens 3.500 grammatikalischen Regeln im relativ „simplen“ modernen Englisch (vgl. Scholl 2023a).
Der Nachweis einer auch nur rudimentär ausgebildeten Grammatik bei Schimpansen war von Girad-Buttoz et al. (2022) allerdings nicht erbracht worden, da die Forscher bei den überdurchschnittlich häufig vorkommenden Rufkombinationen mögliche inhaltliche Bedeutungen gar nicht untersucht hatten. Dagegen können Menschen in ihren ca. 7000 verschiedenen Sprachen durch feine Nuancen im Satzbau (Syntax) eine unglaubliche Fülle an Bedeutungen vermitteln. Es war also bisher nicht bekannt, ob Schimpansen – im Gegensatz zu einigen anderen Tieren wie sonstigen Säugetieren und Vögeln, die keine engere Verwandtschaft zum Menschen aufweisen – überhaupt Informationen mit ihren Rufkombinationen vermitteln. Damit klafft eine gewaltige Lücke zwischen den kommunikativen Fähigkeiten von Schimpansen und den ganz unterschiedlichen, äußerst komplexen menschlichen Sprachen auf der anderen Seite.
Außerdem zeigte Scholl (2023a) weitere schwerwiegende Probleme zur Evolution von Kommunikationsfähigkeiten auf – wie z. B. eine völlig unsystematische Verteilung von Fähigkeiten zu Rufkombinationen (wie auch Konsonantenverwendung) unter Primaten, Fledermäusen, Walen und Singvögeln und auch von auditiven Kommunikationsfähigkeiten und -organen im Tierreich generell. Daraus lässt sich die Schlussfolgerung ableiten, dass evolutionäre Erklärungsmodelle zur Herkunft menschlicher Sprache im Besonderen und auch zur Herkunft genialer, vielfältiger und unsystematisch verteilter Kommunikationsfähigkeiten im Tierreich im Allgemeinen bisher gescheitert sind (vgl. Scholl 2023a; Scholl 2018, 13–20). Dieser Befund könnte stattdessen als Argument für ein biblisches Ursprungsmodell von tierischen Kommunikationsfähigkeiten und menschlichen Sprachen vor allem auf der Basis von 1. Mose Kapitel 1–2 und 10–11 gedeutet werden.
Allerdings hat sich die Datenlage bezüglich der Kommunikationsfähigkeit von Schimpansen mit einer Studie vom Mai 2023 grundsätzlich geändert. Leroux und Kollegen (2023) haben nämlich die allererste Studie über wildlebende Schimpansen vorgelegt, die sich mit einer Rufkombination mit inhaltlicher Bedeutung auseinandersetzt. Doch was bedeuten die neuen Forschungsergebnisse für die Frage nach der evolutionären Herkunft der menschlichen Sprachen?
Neue Studie zur „kompositionellen Syntax“ bei Schimpansen
Leroux et al. (2023, 1) halten zu Beginn ihres Artikels fest, dass die menschliche Sprache etwas Besonderes ist: „Die menschliche Sprache ist ein äußerst produktives Kommunikationssystem, bei dem durch die Kombination vorhandener bedeutungstragender Einheiten (oder Wörter) syntaktisch neue Bedeutung geschaffen werden können“. Dabei können Menschen nicht nur Kombinationen hervorbringen, deren Bedeutungen sich (kompositionell) aus den Bedeutungen der Einzelbestandteile (wie z. B. Wörtern) ergeben. Menschen sind darüber hinaus auch in der Lage, durch Kombinationen völlig neue Bedeutungsinhalte zu erzeugen, was man kombinatorische Syntax nennt (ebd.). Dies erfordert sicherlich eine höhere Abstraktionsleistung als die kompositionelle Syntax, die wahrscheinlich im nachfolgend beschriebenen Einzelfall bei Schimpansen nachgewiesen worden ist. Es ist allerdings noch umstritten („more contentious“), welches der Ursprung der Syntax nach evolutionstheoretischen Vorstellungen ist (ebd.). Auch Leroux et al. (2023, 2) bestätigen die Feststellung von Scholl (2023a), dass noch keine Syntax bei Großen Menschenaffen nachgewiesen wurde – weil nämlich noch gar keine systematischen Arbeiten zu syntaktischen Strukturen bei ihnen durchgeführt worden waren.
Die dem Menschen vermeintlich am nächsten verwandten[2] Schimpansen benutzen zwölf verschiedene Ruftypen (Girad-Buttoz et al. 2022; vgl. zur Verwandtschaft: Scholl 2023b). Dazu gehören auch „Huu“-Rufe (Alarmruf) und „Waa“-Rufe (Ruf zur Unterstützung bei Jagd oder Gefahr, aber auch bei anderen sozialen Kontexten wie Aggression; vgl. Leroux et al. 2023, 2).
Wie sind die Wissenschaftler um Leroux vorgegangen?
Sie erschreckten mittels einer Schlangen-Attrappe dreizehn wildlebende Schimpansen. In neun von 21 Fällen kombinierten die erschrockenen Schimpansen die Rufe „Huu“ und „Waa“ zu „Huu Waa“[3] (ebd.; vgl. Podbregar 2023). Wenn dies geschah, eilten überdurchschnittlich häufig Schimpansen zum Rufenden herbei: In sieben von neun Fällen eilten beim „Huu Waa“ Artgenossen zu dem Rufenden; dies geschah hingegen nur in einem von zwölf Fällen, als nur der Ruf „Huu“ ohne „Waa“ ausgestoßen worden war (Leroux et al. 2023, 2, Fig. 1).
Daher schlussfolgerten die Forscher, dass der Inhalt einen Rekrutierungsruf darstellt, der Gruppenmitglieder zugleich warnen und rekrutieren soll. Die Bedeutung der Rufkombinationen ergibt sich hierbei aus der Bedeutung der beiden Einzelbestandteile (Alarm und Ruf zur Unterstützung), was Leroux et al. (2023, 1) als „eine der kompositionellen Syntax ähnliche Struktur“ deuten.
Um zu auszuschließen, dass der Ruf „Waa“ alleine dieselbe Wirkung haben könnte, wie die Kombination „Huu-Waa“, spielten die Forscher vier Schimpansen per Lautsprechern den aufgenommenen Ruf „Waa“ vor. Fünf Schimpansen wurde „Huu“ und sechs Schimpansen „Huu-Waa“ vorgespielt. „Huu-Waa“ sorgte, wie erwartet, für eine schnellere Reaktion als „Huu“ (nicht aber schneller als bei „Waa“ alleine) sowie für eine deutlich längeren Blickkontakt der Schimpansen als bei „Waa“ oder gar „Huu“ (ebd., Fig. 2). Zudem war der Blickkontakt, den die Forscher als verstärkte Aufmerksamkeit deuten, bei der Rufkombination noch länger, als wenn man die Werte bei den einzelnen Rufen „Huu“ und „Waa“ einfach addieren würde. Weiterhin berichten die Forscher, dass sich bei „Huu-Waa“ – im Gegensatz zu den Einzelrufen – drei von sechs zuhörenden Schimpansen den Lautsprechern näherten. Außerdem kletternden zwei Schimpansen auf einen Baum und schauten herunter, wie dies typisch beim Kontakt mit Schlangen ist (S. 3). Die Autoren schlussfolgern (S. 3): „Es besteht also die Möglichkeit, dass die Kombination der Rufe den Zuhörern spezifischere Informationen bietet als die einzelnen Rufe für sich genommen.“
Syntaktische Fähigkeiten im sonstigen Tierreich
Ähnlich wie Scholl (2023a) stellen Leroux et al. (2023) die bei Schimpansen nachgewiesene Fähigkeit zur kompositionellen Syntax in einen breiteren evolutionären Rahmen im Tierreich.
Fähigkeiten zur Kombination von Rufen – z. T. auch mit inhaltlich neuer Bedeutung im Vergleich zu den Einzelrufen – treten nämlich weit verbreitet in der Systematik der Primaten auf. Nach (Leroux et al. 2023, 2) könnte diese Fähigkeit evolutionär bereits vor der vermuteten evolutionären Aufspaltung in Altwelt- und Neuweltaffen vor 45 Millionen Jahren entstanden sein. Daher sollten solche Fähigkeiten nicht als Beleg für eine Evolution der menschlichen Sprache aus Fähigkeiten von Schimpansen (bzw. schimpansenähnlichen Vorfahren) verwendet werden. Wenn man schon von einem evolutionären Paradigma ausgeht, müsste diese Fähigkeit viel älter sein, wie auch Leroux et al. in Bezug auf die Primaten zugeben – oder mehrfach parallel entstanden sein, wie die Autoren in Bezug auf solche Fähigkeiten bei Vögeln vermuten.
Zu den syntaktisch begabten Primaten gehören die Große Weißnasenmeerkatzen. Diese kombinieren zwei Rufe, die mit Feinden bzw. Störungen zusammenhängen, in eine längere Sequenz, die mit der Bewegung der Gruppe zu tun hat (Leroux et al. 2023, 2). So werden die Rufe für „Adler-Warnung“ und „Störung“ zu einem Ruf kombiniert Ruf für „Bewegung!“ (ebd.). Da der Inhalt der Rufkombination nicht direkt mit der Bedeutung der beiden Einzelrufe zusammenhängt, repräsentiert dies einen Fall der der anspruchsvollen kombinatorischen Syntax (vgl. ebd.). Mit einer Zusatzsilbe („Oo“) können Campbell-Meerkatzen außerdem zwei verschiedene spezielle Alarmrufe in weniger dringliche bzw. spezifische Alarmrufe umwandeln, die sich aber auf denselben Bereich (am Boden bzw. in der Luft) wie im ursprünglichen Ruf beziehen. Diese Fähigkeit wurde von Wissenschaftlern wiederholt als „rudimentäre“ kompositionelle Syntax interpretiert (ebd.).
Allerdings lassen Leroux et al. (2023, 2) solche Kombinationsfähigkeiten nicht generell als Hinweise auf eine gemeinsame Abstammung gelten, da sie vergleichbare Fähigkeiten bei Vögeln stattdessen als „konvergente“ (also unabhängige) Evolution deuten wollen (vgl. auch Scholl 2023a). Denn Drosselhäherlinge (Turdoides) und japanische Meisen können ebenfalls einen Alarmruf mit einem Rekrutierungsruf kombinieren, wenn sie bei Gefahr Unterstützung benötigen (Leroux et al. 2023, 2; Scholl 2023a). Dies entspricht exakt der „rudimentären“ kompositionellen Syntax, die die Forscher bei Schimpansen sehen, tritt aber im Stammbaum der Lebewesen ungefähr 300 Millionen Jahre vermeintliche Evolutionsgeschichte entfernt auf.

Abb. 1 Nicht nur Schimpansen, auch andere, nicht nahe verwandte Tiere können die Bedeutung von einzelnen Rufen ändern, wenn sie diese kombinieren (v. l.): Große Weißnasen-Meerkatzen, Campbell- bzw. Lowe-Meerkatzen, japanische Meisen und Drosselhäherlinge. (Wikimedia: LaetitiaC, CC BY-SA 3.0; Badgernet, CC BY-SA 3.0; Gerrie van Vuuren, CC BY-SA 4.0)
Schlussfolgerungen
Fasst man die Befunde zusammen, so wurde somit erstmals eine Kombination zweier Rufe bei Schimpansen nachgewiesen, die wahrscheinlich eine eigenständige Information übermittelt. Diese Rufkombination zur Rekrutierung von Artgenossen ist inhaltlich allerdings der Bedeutung der Einzelrufe (Alarm für „Huu“ und Rekrutierung für „Waa“) sehr nahe, was man daher als einen Fall von kompositioneller Syntax deuten kann. Kompositionelle Syntax tritt aber auch bei anderen, nicht näher mit Menschen verwandten Primaten und auch Vögeln auf. Und bei Campbell-Meerkatzen ist sogar die noch anspruchsvollere Stufe der kombinatorischen Syntax belegt, obwohl diese Affen durch 20 bis 47 Millionen radiometrische Jahre Evolutionsgeschichte von den großen Menschenaffen getrennt sein sollen (je nach Autor; s. Scholl 2023a, Tab. 1). Somit ist auch der neue Nachweis für eine wahrscheinliche Verwendung von kompositioneller Syntaxbei Schimpansen kein Argument, das die Hypothese einer Abstammung der menschlichen Sprache und vor allem Syntax von den Rufen bei Schimpansen nahelegt. Es bleibt aus evolutionärer Perspektive rätselhaft, dass die syntaktischen Fähigkeiten bei Großen Weißnasenmeerkatzen nach aktuellem Kenntnisstand den Menschen stärker ähneln als jene wildlebender Schimpansen. Somit bleibt es dabei, dass eine gewaltige Kluft zwischen den kommunikativen Fähigkeiten von Schimpansen und Menschen besteht (vgl. Scholl 2018; 2023a), die besser durch ein biblisches Schöpfungsmodell als durch ein Evolutionsmodell erklärt werden kann.
Quellen
Girard-Buttoz C et al. (2022) Chimpanzees produce diverse vocal sequences with ordered and recombinatorial properties. Commun. Biol. 5, 410, https://doi.org/10.1038/s42003-022-03350-8.
Leroux M et al. (2023) Call combinations and compositional processing in wild chimpanzees. Nat. Commun. 14, 2225, https://doi.org/10.1038/s41467-023-37816-y.
Podbregar N (2023) Schimpansen kombinieren Rufe zu neuen Bedeutungen, natur.de vom 05.05.2023, https://www.wissenschaft.de/erde-umwelt/schimpansen-kombinieren-rufe-zu-neuen-bedeutungen/.
Scholl B (2018) Affe = Mensch? Ein Überblick über verhaltensbiologische Unterschiede zwischen Affen und Menschen. W+W Special Paper B-18-1, https://www.wort-und-wissen.org/wp-content/uploads/b-18-1_affe-mensch.pdf.
Scholl B (2022) „Totales Chaos“: Unklare Abstammungsverhältnisse bei Menschenaffen und Menschen. W+W Special Paper B-22-2, https://www.wort-und-wissen.org/wp-content/uploads/b-22-2_menschenaffen.pdf.
Scholl B (2023a) Beherrschen Schimpansen etwa doch Grammatik? W+W Online-Artikel vom 23.02.2023, https://www.wort-und-wissen.org/wp-content/uploads/Schimpansengrammatik.pdf.
Scholl B (2023b) „Totales Chaos“: Unklare Abstammungsverhältnisse bei Menschenaffen und Menschen. W+W Special Paper B-22-2, https://www.wort-und-wissen.org/wp-content/uploads/b-22-2_menschenaffen.pdf.
Anmerkungen:
[1] https://www.wort-und-wissen.org/artikel/schimpansengrammatik/.
[2] Solche Verwandtschaftsverhältnisse sind lediglich evolutionstheoretische Postulate und können nicht empirisch bewiesen werden. Empirisch sind hingegen nur abgestufte Ähnlichkeiten; diese kann man sachlich beschreiben. Ähnlichkeiten können im Gegensatz zu gemeinsamer Abstammung immer auch als Hinweise auf einen gemeinsamen Schöpfer gedeutet werden, der ähnliche Merkmale zugeteilt hat (vgl. Scholl 2022).
[3] Die Kombination „Huu-Waa“ tauchte unter den ca. 5000 in der Wildnis aufgezeichneten Rufen von Girad-Buttoz et al. (2022, Suppl. Data 2: „Hoo“+“Bark“ = „HO-BK“) acht Mal auf. Diese Kombination war dort insgesamt nicht besonders häufig oder auffällig. Allerdings beträgt die (A-posteriori-) Wahrscheinlichkeit 83 %, dass ein „Huu“ allgemein am Anfang einer Zweiereinheit auftritt; „Waa“ („Bark“) tritt zu 72 % am Ende einer Zweiereinheit auf (ebd., Tab. 3). Da „Huu“ nach Leroux et al. (2023) einen Alarm darstellt, der auch ohne „Waa“ im selben Kontext (Schlangen-Sichtung) von Schimpansen verwendet wird, ist generell zu erwarten, dass beide Rufe typischerweise in dieser Reihenfolge abgeben werden. Die Reihenfolge an sich erfordert also nicht die Annahme einer zusätzlichen grammatikalischen Regel (s. Scholl 2023a).
Autor dieser News: Benjamin Scholl
© 2023, http://www.genesisnet.info/schoepfung_evolution/n316.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
10.05.23 Banyoles-Unterkiefer: Kein Neandertaler und doch nicht der älteste moderne Mensch in Europa?
Wie alt ist der älteste fossile moderne Mensch (Homo sapiens) in Europa? Keeling et al. (2023) schlagen vor, dass der Banyoles-Unterkiefer ein früher Homo sapiens mit einem Alter von 45.000 bis gar 66.000 rJ gewesen sei. Moderne Menschen wären demzufolge bereits zu einer Zeit in Europa gewesen, aus der wesentliche archäologische Funde stammen, die man bisher dem Intellekt der Neandertaler zugeschrieben hatte. Eine gründliche Analyse der Daten zeigt jedoch, dass die Hypothese, der Unterkiefer von Banyoles gehöre zu einem modernen Menschen, spekulativ ist. Stattdessen unterstützen die netzwerkartig verteilten Merkmale die Grundtypenhypothese, die von einem einzigen erschaffenen Grundtyp der Menschen ausgeht.
- Einleitung
Das erste Erscheinen des modernen Menschen in Europa hat sich durch neuere Funde nach hinten verschoben und wird nun älter als ca. 45.000 rJ eingestuft. So schreiben Keeling et al. (2023, 4): „Obwohl ein möglicherweise viel früheres H. sapiens-Fossil aus Apidima in Griechenland mit 210.000 Jahren gemeldet wurde […], wurde der früheste fossile Nachweis für H. sapiens aus dem Spätpleistozän kürzlich in der Grotte Mandrin in Frankreich auf 56.800 bis 51.700 Jahre [rJ] datiert […].“ Allerdings besteht bei den beiden genannten Homo–sapiens-Funde aus Griechenland[1] und Frankreich[2] noch kein Konsens.
Keeling et al. (2023, 1, 14–16) schlagen vor, dass der seit 1887 bekannte Unterkiefer von Banyoles (Nordost-Spanien) kein Neandertaler, sondern wahrscheinlich ein sehr früher Homo sapiens aus Europa mit einem Alter von 45.000 bis gar 66.000 rJ gewesen sei. So titelte auch scinexx.de „Ist dies das älteste Homo-sapiens-Fossil Europas?“ (Bernhard 2022). Die Behauptung, der Banyoles-Unterkiefer sei ein moderner Mensch gewesen, könnte in Zukunft als Argument verwendet werden, um archäologische Hinterlassenschaften, die bisher als Belege der intellektuellen Ebenbürtigkeit von Neandertalern mit uns modernen Menschen angesehen wurden (vgl. Frayer & Radovčić 2022a/b; Scholl 2023), z. T. als potenzielle Hinterlassenschaften moderner Menschen umzudeuten. Im Folgenden wird aufgezeigt, warum der Vorschlag von Keeling et al. (2023), dass Banyoles ein modern-menschlicher Unterkiefer sei, aus mehreren Gründen äußerst problematisch ist.
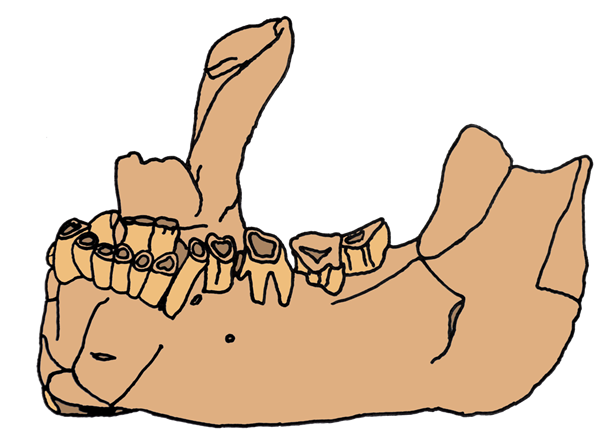
Abb. 1 Skizze des Banyoles-Unterkiefers (Eigene Darstellung)
- Probleme der Zuordnung des Banyoles-Unterkiefers zu Homo sapiens
Erstens „sind sich die Fachleute über die taxonomische Zugehörigkeit dieses Fossils uneins“ (Keeling et al. 2023, 1). Dies zeigt auch der Blick in die über ein Jahrhundert zurückreichende Forschungsgeschichte mit kontroverser Diskussion seiner Merkmale und Zuordnung. So sind beim Unterkiefer von Banyoles von vielen Forschern Ähnlichkeiten mit Neandertalern oder Mittelpleistozänen Menschen festgestellt worden[3]; von letzteren gab es solche mit typischen Neandertaler-Merkmalen (z. B. Sima de los Huesos und Ehringsdorf) und andere ohne (wie z. B. Mauer).
Zweitens fehlt der klare archäologische Kontext sowie eine eindeutige Datierung (Keeling et al. 2023, 3). Man kennt den genauen Fundort nicht, weiß aber, dass der Unterkiefer von Baynoles in einem Steinbruch Pla de la Fromgiga etwa vier Meter unter der Oberfläche in einem Kalkstein-Travertin-Tuff gefunden wurde. Die „heterogenen“ Datierungsergebnisse reichen von ca. 16.000 bis 66.000 rJ (vgl. Anhang 1), wobei Keeling und Kollegen (ebd.) ein Alter von ca. 45.000 bis 66.000 rJ bevorzugen (bei ± 4.000 / 7.000 rJ). Selbst wenn dies stimmen sollte, ist also keineswegs belegt, dass der Unterkiefer älter ist als die bisher ältesten Funde von Homo sapiens in Europa, da bereits Funde an der Untergrenze der möglichen Datierung (ca. 45.000 rJ) aus Europa bekannt sind.
Das dritte Problem: Die erhaltenen Merkmale des Unterkiefers (z. B. ein fehlendes vorstehendes Kinn) „erschweren ebenfalls eine Zuordnung“ und lassen die taxonomische Entscheidung „aktuell offen“ (Keeling et al. 2023, 16).
Die Schwierigkeiten bei der Zuordnung von Banyoles werden auch anhand der folgenden Tabelle 1 ersichtlich: Es gibt kaum Merkmale des Unterkiefers, die nur bei Homo sapiens, nicht aber bei Neandertaler oder dem Homo aus dem Mittelpleistozän (bzw. Homo heidelbergensis) auftreten. Selbst vermeintlich typische Merkmale einer Menschenform finden sich in geringerer Häufigkeit auch bei anderen Menschenformen. Harvati & Reyes-Centeno (2022, 3) halten fest, dass es in Paläoanthropologie sogar gängige Praxis ist, solche unscharfen Merkmale (vgl. Junker & Scherer 2013, 28f, 32f, 240) wie Häufigkeitsunterschiede statt einzigartiger abgeleiteter Merkmale als Artzuschreibungskriterien bei fossilen Menschen zu verwenden: „Unterschiede in der Häufigkeit bestimmter Merkmalsausprägungen oder Merkmalskombinationen werden daher häufig als definierende Merkmale für die Diagnose akzeptiert.“
Tabelle 1: Zusammenstellung einiger Merkmale bei anderen Menschenformen, die exakt den Merkmalen des Unterkiefers von Banyoles entsprechen. Die Farben zeigen die größten (quantitativen) Ähnlichkeiten (von dunkelgrün) bis hin zu fehlende Merkmale (rot) bei den jeweiligen untersuchten Menschenformen. (Eigene Darstellung nach Keeling et al. 2023, Tab. 3–10)
Keeling et al. (2023, 7) erstellten Flächenvergleiche mithilfe von 22 Messpunkten („Landmarken“). Anschließend erstellten sie fünf Procrostus-Analysen (PC), welche ca. 68 % der Varianz erklären. Allerdings tragen PC 1, 3, 4 und 5 überhaupt nicht zur taxonomischen Klarheit bei (vgl. ebd. 12; Fig. 6). Lediglich bei PC 2, die ca. 16 % der Varianz erklärt, fielen die Banyoles-Rekonstruktion in den Variationsbereich von Homo sapiens.[4] In Bezug auf die gesamte Formvariation bestehen aber signifikante Unterschiede von Banyoles zu allen untersuchten Menschenformen (S. 13).
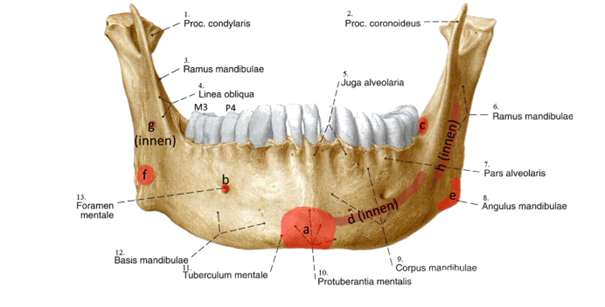
Abb. 2: Unterkiefer eines heutigen Menschen mit den markierten Vergleichsregionen beim Banyoles-Unterkiefer. (nach Wikimedia: Partynia nach Kankoreziu103; CC BY-SA 4.0)
- Schlussfolgerungen
Insgesamt schlussfolgern Keeling et al. (2023, 15): „Auch wenn die genaue taxonomische Einordnung unsicher ist, handelt es sich bei Banyoles eindeutig nicht um einen Neandertaler“. Allerdings bedeutet dies nicht automatisch, dass Banyoles ein moderner Mensch gewesen ist. Schließlich halten die Autoren (S. 14) die meisten Merkmale von Unterkiefern beim modernen Menschen generell für Beibehaltungen („retentions“) primitiver bzw. ursprünglicher Merkmale älterer Menschenformen. Daher ist die taxonomische Zuordnung eines Unterkiefers zum modernen Menschen generell schwierig.
Ähnlich halten auch Harvati & Reyes-Centeno (2022, 3) fest, dass ein allgemeines Problem in der Abgrenzung verschiedener Menschenformen – und insbesondere bei Homo sapiens – besteht: „Die Diagnose von Vorfahren des modernen Menschen im Fossilbericht ist nicht immer einfach.“ Dies sei bei Neandertalern leichter, weil sie durch viele abgeleitete Merkmale/Merkmalskombinationen einfacher zuzuordnen sind. Andererseits können Merkmale des modernen Menschen, die als diagnostisch gedeutet wurden, die tatsächliche geografischen und zeitlichen Vielfalt des modernen Menschen nicht wirklich erfassen, weil sie zu eng gefasst („restriktiv“) sind (ebd.).
Dies zeigt sich beispielhaft beim Unterkiefer, da die Existenz eines knöchernen Kinns das „einzige eindeutig abgeleitete Merkmal im Unterkiefer von H. sapiens“ darstellt (Keeling et al. 2023, 14). Und selbst dies gilt nur für die gesamte Kombination der Merkmale des knöchernen Kinns, weil einzelne Merkmale bei Neandertalern und sogar bei verschiedenen Formen von Homo erectus vorkommen (vgl. S. 14).[5] Allerdings ist „bemerkenswert“, dass die Mehrzahl der modern-menschlichen Unterkiefer-Merkmale bei Banyoles eben nicht vorhanden ist (ebd.).
Dennoch bevorzugen die Autoren die Hypothese, dass Banyoles einer der frühsten modernen Menschen in Europa war. Und so spekulieren sie über einen Zusammenhang der fehlenden sapiens-Merkmale mit starker Abnutzung der vorderen Zähne (S. 14); oder verweisen auf eine abgeschwächte Form bzw. Variabilität der Kinnmerkmale bei frühen Funden von Homo sapiens (S. 15), die aber auch 30.000 bis 200.000 rJ älter sind als Banyoles.[6] Es besteht also bezüglich der Merkmale des Unterkiefers von Banyoles ein weiterer, nicht gut einzuordnender Merkmalsmix. Die Autoren halten es sogar für möglich, dass eine völlig unbekannte archaische Menschenform, die keine Neandertaler waren, sich mit Homo sapiens gekreuzt haben könnte und so das Merkmalsmosaik von Banyoles erzeugt hätte (S. 14f).
Dies alles bleibt aber Spekulation. Denn vom morphologischen Befund her gilt: „Banyoles weist also keine abgeleiteten Neandertaler- oder H. sapiens-Merkmale auf. Vielmehr scheint die Morphologie von Banyoles die einer verallgemeinerten, primitiven Form der Gattung Homo zu sein“ (Keeling et al. 2023, 14). Dies wiederrum spricht eher für den – ebenfalls spektakulären – Befund, dass Banyoles auf solche mittelpleistozänen Menschen zurückgehen könnte, die – entsprechend ihrer Skelettmerkmale – nicht in den Neandertalern aufgegangen sind.
- Der Grundtyp Mensch als Netzwerk
Es zeigt sich wieder einmal, dass zwischen Homo erectus als frühestem fossilen Menschen und dem heutigen Homo sapiens ein nicht zu entwirrendes Netzwerk von fossilen Menschenformen mit ganz unterschiedlichen Merkmalsmosaiken besteht (vgl. Brandt 2020, 216–230; Brandt 2018). Dies scheint insbesondere durch Ergebnisse des Vergleichs fossiler DNA- und Eiweiß-Sequenzen für die wissenschaftliche Fachwelt immer deutlicher zu werden (vgl. Anhang 2), wie auch das folgenden Zitats von Schlott (2021; Hervorh. hinzug.) von spektrum.de aufzeigt: „Doch die Paläogenetik hat auch gezeigt, dass sich die Trennung von verschiedenen Menschenarten nicht mehr so scharf aufrechterhalten lässt. Auch deshalb ist inzwischen häufiger von Menschenformen die Rede als von -arten, zumal sich anatomisch moderner Mensch, Neandertaler und Denisovaner [die Schwestergruppe der Neandertaler aus dem östlichen Asien] mehrfach miteinander vermischt und sich im Lauf der Zeit in Unterpopulationen aufgespalten haben“ (vgl. Anhang 2).
Diesen Befund nennen Wissenschaftler in Bezug auf das Mittelpleistozän[7] „the muddle in the middle“ (z. B. Roksandic et al. 2021, 1f; Harvati & Reyes-Centeno 2022, 1f) – das „Wirrwarr“ bzw. „undurchdringliche Geflecht“ der Merkmale der Menschenformen. Abgesehen von Homo erectus, sapiens und neanderthalensis haben Wissenschaftler schon mehr als ein Dutzend verschiedene Menschenformen in diesem Übergangsbereich vorgeschlagen (vgl. Harvati & Reyes-Centeno 2022, Tab. 1)[8] – allerdings, ohne dabei einen breiten Konsens zu erzielen. Letztlich scheint sich auch die Aufstellung des neuen Taxons Homo bodosiensis (statt Homo heidelbgergensis/rhodensis) und des neuen Taxons des „Drachenmenschen“ namens Homo longi (evtl. ein Denisova, vgl. Line 2021, 122f; George 2021; Schlott 2021) nicht wirklich durchzusetzen (s. die Diskussion von Roksandvic et al. 2021; Sarmiento & Pickford 2022; Delson & Stringer 2022; Roksandvic et al. 2022; vgl. Ji et al. 2021; Handwerk 2021; Harvati & Reyes-Centeno 2022, 2; Line 2022, 89f, 98).

Abb. 3: Andere Menschenformen mit Merkmalsmosaiken: Skizze des Harbin-Schädels („Homo longi”), Bodo-Schädel („Homo bodoensis”) und Nesher-Ramla-Homo. (Wikimedia: nach Hemiauchenia, CC BY-SA 4.0; Ryan Somma, CC BY-SA 2.0; ד“ר יוסי זיידנר – Yossi Zaidner; CC BY-SA 4.0)
So schreiben Harvati & Reyes-Centeno (2022, 2) im Kontext der Diskussion um die Umbenennung einiger Funde von Homo heidelbergensis zu Homo longi bzw. Homo bodoensis: „In diesem Wirrwarr von vorgeschlagenen Homininen-Arten ist kein Taxon umstrittener als Homo heidelbergensis […].“ Dies liegt zum einen daran, dass das Typusexemplar aus Mauer in Deutschland nur ein einziger Unterkiefer ist. Zum anderen wird Homo heidelbergensis nicht durch „eigene, eindeutig abgeleitete [bzw. moderne] Merkmale definiert“, sondern durch eine Schädelmorphologie, die zwischen dem frühen Homo erectus und späteren Menschenformen überbrücken soll. Roksandvic et al. (2022, 2) bezeichnen Homo heidelbergensis sogar einfach als „Abfalleimer“ („wastebasket“), in den alles geschmissen wird, was nicht H. erectus, sapiens oder neanderthalensis ist. Und nach Hartwig-Scherer (2002) ist Homo heidelbergensis „ein einzigartiger Schmelztiegel unterschiedlicher Morphologiemosaike aus erectus-, heidelbergensis-, neanderthalensis– und sapiens-anmutenden Merkmalen, die sich kaum in eine evolutiv-lineare Abfolge bringen lassen“.
Harvati & Reyes-Centeno (2022, 8) beenden ihren Artikel über die Vielfalt der Menschen im Mittelpleistozän mit folgendem Statement: „Abschließend möchten wir betonen, dass das verwirrende, komplexe Bild, das die menschliche Evolution im mittleren bis späten Pleistozän bietet, als eine Besonderheit und nicht als ein Fehler angesehen werden sollte, da es die evolutionären Prozesse in all ihrer Unordnung widerspiegelt.“ Dem ist aus Schöpfungsperspektive zuzustimmen, wenn statt darwin’scher Makroevolution von mikroevolutionären Prozessen (innerhalb des erschaffenen Grundtyps Mensch) ausgegangen wird.
Auch morphologische und genetische Befunde (vgl. Tab. 1; Anhang 2 sowie Brandt 2017, 109–115; Brandt 2018; Brandt 2020, 216–230; Scholl 2020; Scholl 2022; Borger 2021; Binder & Borger 2022) sprechen für ein evolutionär nicht aufzulösendes Netzwerk der Vermischung verschiedener Menschenformen. Daher ist es kein Wunder, dass die Suche nach dem letzten gemeinsamen Vorfahren von modernem Mensch, Neandertaler und Denisova „gemischte und sogar widersprüchliche Ergebnisse“ geliefert hat und somit ungelöst bleibt (Harvati & Reyes-Centeno 2022, 6f).
Statt ständig den Fossilfunden neue eigene Menschenarten zuzuweisen, erscheint es deutlich besser begründet, sie alle als Ausprägungen und Spezialisierungen innerhalb des Grundtyp-Netzwerkes der echten Menschen zu sehen (vgl. Brandt 2020, 216–230). Die echten Menschen wurden laut Genesis 1–2 (die Bibel) von Gott als Menschen erschaffen. Dem Grundtyp-Modell zufolge passten sie sich dann vor allem aufgrund von Rekombination (Durchmischung des Erbgutes bei der sexuellen Vererbung) in ganz verschiedenen Merkmalskombinationen an ihre Umweltbedingungen an (vgl. Crompton 2019). Dementsprechend überrascht es auch nicht, wenn ständig neue menschliche Fossilien bekannt werden, die sich wie der Unterkiefer von Banyoles durch ein einzigartiges menschliches Merkmalsmosaik auszeichnen, das sich weder evolutionär noch systematisch klar einordnen lässt.
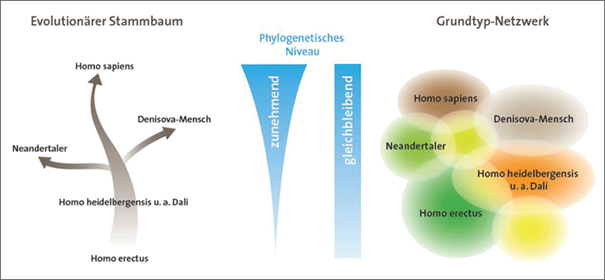
Abb. 4 Verwandtschaftsverhältnisse des Menschen nach dem phylogenetischen Modell (links) und dem Grundtypmodell. (Quelle: Brandt 2018, Abb. 2)
Anhang
Anhang 1: Verschiedene und widersprüchliche Datierungsergebnisse des Unterkiefers von Banyoles (zitiert nach Keeling et al. 2023, 3):
Radiokarbondatierung auf 17.000 ± 1.000 rJ (Berger & Libby 1966); Stratigrapische Schätzungen: Mittel- bis Spät-Pleistozön (Hernandez-Pacheco & Obermaier 1915; Bonarelli 1916; Sole Sabarís 1957; Bech 1971) – entsprechendes ergaben auch die Tierarten der Travertinmatrix (Maroto & Soler 1993); Uran-Thorium-Datierung: 16.200 ± 3.200 rJ des Unterkiefers und 70.000 bzw. 110.000 rJ (geschlossenes bzw. offenes System) für den Travertin (Yokoyama et al. 1987); Uran-Thorium-Datierung vom Travetin, der direkt vom Unterkiefer entfernt worden war: 45.000 ± 4.000 rJ; Travetinmatrix aus Pla de la Mata: 45.000–50.000 rJ (Julia & Bischoff 1991). Aus letzterem wurde von Julia & Bischoff (1991) geschlossen, dass der Unterkiefer „sehr wahrscheinlich“ aus der Travetinmatrix von Pla de la Mata stammt und dass die Fundstelle wahrscheinlich ein geschlossenes (d. h. geologisch nicht umgearbeitetes) System darstellt. Schließlich führten Grün et al. (2006) Uran-Thorium-Datierung / Elektronenspin-Resonanz-Datierung durch und kamen beim Zahnschmelz des 3. Backenzahns auf 66.000 ± 7.000 rJ und bei der dem Unterkiefer anhaftenden Travetinmatrix auf 42.500 ± 4.100 rJ. Solche „heterogenen“ Ergebnisse könnten für eine geologische Umarbeitung/Umlagerung des Unterkiefers sprechen, was Keeling et al. (2023, 3) aber für unwahrscheinlich halten. Aus diesen widersprüchlichen Ergebnissen leiten Keeling und Kollegen (ebd.) ein Alter von ca. 45.000 bis 66.000 rJ ab (bei ± 4.000 / 7.000 rJ).
Anhang 2: Hinweise auf Vermischung von Homo sapiens mit älteren Menschenformen nach Keeling et al. (2023, 14; abgekürzt durch n. K.) und anderen Autoren:
- Oase 1: früher Homo sapiens aus Peștera cu Oase (Rumänien) vor 37.000–42.000 rJ: 6–9 % Neandertaler-Genvarianten, also ungefähr 4–6 Generationen vorher ein Neandertaler-Vorfahre (Fu et al. 2015; n. K.)
- Drei moderne Homo sapiens aus Vindija (Kroatien): 3,4 bis 7,9 % Neandertaler-Varianten (Lohse & Frantz 2014; n. K.)
- mt-DNA von Knochenfragmenten des modernen Homo sapiens aus Bacho-Kiro-Höhle (Bulgarien) mit 46.000 rJ: kein Hinweis auf Neandertaler-Genfluss (Hublin et al. 2020; n. K.)
- Moderner Homo sapiens aus Ust-Ishim (Westsibirien) vor 45.000 rJ: 2,3 % Neandertaler-Varianten (Fu et al. 2014; n. K.)
- Moderner Homo sapiens aus Zlatý Kun (Tschechien) vor mind. 45.000 rJ: ca. 3 % Neandertalervarianten (Prüfer et al. 2021; n. K.)
- Moderner Homo sapiens im Nahen Osten vor 50.000–80.000 rJ: Vermischungsereignis mit Neandertaler-Varianten (Green et al. 2010; Smith 2013; Wall et al. 2013; n. K.) – nach Green et al. (2010) einmalige Vermischung; Keeling et al. (2023, 14) sprechen insgesamt von wiederholen Vermischungsereignissen zwischen Neandertaler und modernem Menschen (nicht nur im Nahen Osten).
- Der letzte Genfluss zwischen Neandertalern und nichtafrikanischen Homo sapiens geschah vermutlich zwischen 47.000–65.000 (Sankararaman et al. 2012; n. K.).
- Bei Neandertalern vor 270.000 rJ evtl. Genfluss von Afrikanern (Posth et al. 2017)
- Verschiedene Studien wiesen auf Hybride innerhalb der Gattung Homo hin aufgrund von abgeleiteten Körpermerkmalen (Duarte et al. 1999; Tattersall & Schwartz 1999; Trinkaus et al. 2003; Gunz & Harvati 2011; Smith 2013; Ackermann et al. 2016; Smith et al. 2017; n. K.; vgl. Brandt 2020, 216–230; Brandt 2018). Keeling et al. (2023, 14) kommentieren: „die Mischung von Neandertaler- und H.-sapiens-Merkmalen in Oase 1 steht im Einklang mit dieser Perspektive“.
- Heutige Menschen haben im Durchschnitt 1,4–2,1 % Neandertaler-Genvarianten– basierend auf dem Altai-Neandertaler-Genom (Prüfer et al. 2014; n. K.).
- Neandertaler und Denisova könnten sich vor 381.000–473.000 rJ getrennt haben (Kuhlwilm et al. 2016, 429; Meyer et al. 2016, 504+506). Die Trennung von Homo sapiens und älteren Menschenformen könnte bereits vor 430.000 rJ oder gar 550.000–765.000 rJ geschehen sein (ebd.; vgl. Posth et al. 2017, 2). Das Y-Chromosom spricht für eine Trennungszeit von Neandertaler und modernem Mensch vor 806.000–447.000 rJ und die mitochrondriale DNA für 489.000–295.000 rJ (wahrscheinlich ca. 400.000 rJ); Posth et al. (2017, 2, Tab. 1) kommen auf 467.720–360.230 rJ (Ø 412.930 rJ) für die mtDNA. Dies ist gar nicht so viel, wenn man bedenkt, dass sie bei den heute lebenden San eine Aufspaltungszeit (Divergenzzeit) von 169.520–123.650 rJ (Ø 146.730 rJ) errechnet haben (ebd., Tab. 1).
- Slon et al. (2018, 113) berichten davon, dass Denisova 11 aus der Denisova-Höhle (Altaigebirge, Sibirien) eine Neandertaler-Mutter und einen Denisova-Vater hatte; bereits bei Denisova 3 war man von einem Vermischungsereignis der beiden frühen Menschen ausgegangen. Kuhlwilm et al. (2016, 429) schreiben außerdem: „Das Genom des Altai-Neandertalers hat 5,4 % mehr abgeleitete Allele mit den heutigen Afrikanern gemeinsam als das Genom der Denisovaner.“
- Browning et al. (2018, 59f) berichten davon, dass Ostasiaten sogar von zwei verschiedenen Denisova-Populationen Genvarianten gerbt haben und dass diese wahrscheinlich einen positiven Einfluss auf das Immunsystem besitzen.
- Meyer et al. (2016, 504) schreiben, dass die Genvarianten des Zellkerns der 430.000 rJ alten Heidelberger Menschen aus Sima de los Huesos (Spanien) eher Neandertalern gleicht, obwohl ein Individuum dieser Gruppe in den Genvarianten des Mitochondriums stärker den Denisova ähnelt.
- Heutige Menschen aus Papua besitzen ca. 5 % Genvarianten der Denisova-Menschen; auch die Einwohner anderer südostasiatischer Inseln sowie von Australien besitzen bis zu 5 % Denisova-Varianten (Vespasiani et al. 2022, 1; vgl. Reich et al. 2010, 1: Melanesier haben ca. 4–6 % Denisova-Genvarianten).
- Lorente-Galdos et al. (2019, 1+10) vermuten, dass eine noch unbekannte Menschenform (ungefähr gleich alt wie Neandertaler und Denisova) ihre Genvarianten in Afrikanern aus Subsahara hinterlassen hat (Khoisan: ca. 4 %, Mbuti Pygmäen: ca. 4,3 % und Mandenka: 5,8 %). Mittlerweile gibt es einige solcher Hypothesen.
- Ragsdale et al. (2023) haben aufgrund umfassender Genomanalysen modelliert, dass Menschen seit ca. 1.000.000 rJ in Afrika ein Fortpflanzungsnetzwerk bilden; daher könnte man sie nach Coleman (2023) in nature news alle als eine einzige Art bezeichnen. Ragsdale et al. (2023) sind der Meinung, ihr Modell erkläre die Daten besser, als archaische „Geisterlinien“ – also unbekannte ausgestorbene Menschenformen in Afrika.
Quellen:
Bernhard E (2022) Ist dies das älteste Homo-sapiens-Fossil Europas? scinexx.de, vom 09.12.2022, https://www.scinexx.de/news/archaeologie/ist-dies-das-aelteste-homo-sapiens-fossil-europas/.
Binder H & Borger P (2022) Nobelpreis für Forschungen über das Erbgut aus menschlichen Fossilien. Genesisnet.info News, vom 20.10.2022, http://www.genesisnet.info/schoepfung_evolution/n306.php.
Borger P (2021) Kaum einzigartige DNA-Sequenzen im Erbgut des Menschen. Stud. Integr. J. 28, 134–135.
Brandt M (2017) Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special. SCM Hänssler.
Brandt M (2018) Merkmalsnetz statt Stammbaum. Neues Vernetzungsmodell in der Paläanthropologie ähnelt Verwandtschaftsverhältnissen im Grundtyp Mensch. Stud. Integr. J. 25, 47–51, https://www.si-journal.de/jg25/heft1/sij251-9.html.
Brandt M (2020) Wie alt ist die Menschheit? Demographie und Steinwerkzeuge mit überraschenden Befunden. 6., erw. Aufl. Studium Integrale Vor- und Frühgeschichte. SCM Hänssler.
Browning SR et al. (2018) Analysis of Human Sequence Data Reveals Two Pulses of Archaic Denisovan Admixture. Cell 173, 53–61, https://doi.org/10.1016/j.cell.2018.02.031.
Coleman J (2023) Human-evolution story rewritten by fresh data and more computing power. nature news, vom 18.05.2023, https://www.nature.com/articles/d41586-023-01664-z.
Crompton N (2019) Mendel’sche Artbildung und die Entstehung der Arten. W+W Special Paper B-19-3, https://www.wort-und-wissen.org/artikel/mendelsche-artbildung-und-die-entstehung-der-arten/.
Delson E & Stringer C (2022) The naming of Homo bodoensis Roksandic et al., 2021 does not resolve issues surrounding Middle Pleistocene human evolution. Evol. Anthropol. 31, 233–236.
Frayer DW & Radovčić D (2022a) Rockshelter Discoveries Show Neandertals Were a Lot like Us. Scientific American, vom 01.02.2022, https://www.scientificamerican.com/article/rockshelter-discoveries-show-neandertals-were-a-lot-like-us/.
Frayer DW & Radovčić D (2022b) Neandertaler. Menschen wie wir. Spektrum der Wissenschaft 11.22, 78–83, spektrum.de/artikel/2057463.
Hartwig-Scherer S (2002) Wurde Europa doch früher besiedelt? Überraschungen aus Ost und West. Stud. Int. J. 9, 67–73.
Harvati K & Reyes-Centeno H (2022) Evolution of Homo in the Middle and Late Pleistocene. J. Hum. Evol. 173, 103279, https://doi.org/10.1016/j.jhevol.2022.103279.
Ji Q et al. (2021) Late Middle Pleistocene Harbin cranium represents a new Homo species. The Innovation 2, 100132, https://doi.org/10.1016/j.xinn.2021.100132.
Junker R & Scherer S (Hrsg) (2013) Evolution. Ein kritisches Lehrbuch. 7., aktual. und erw. Aufl. Weyel Biologie.
George A (2021) ‚Dragon man‘ claimed as new species of ancient human but doubts remain. New Scientist, vom 25.06.2021, https://www.newscientist.com/article/2282223-dragon-man-claimed-as-new-species-of-ancient-human-but-doubts-remain/.
Handwerk B (2021) A 146,000-Year-Old Fossil Dubbed ‘Dragon Man’ Might Be One of Our Closest Relatives. A mysterious Middle Pleistocene skull from a Chinese well has inspired debate among paleoanthropologists. Smithsonian Magazine, vom 25.06.2021, https://www.smithsonianmag.com/science-nature/146000-year-old-fossil-dubbed-dragon-man-might-be-one-our-closest-relatives-180978062/.
Keeling BA et al. (2023) Reassessment of the human mandible from Banyoles (Girona, Spain). J. Hum. Evol. 17, 103291, https://doi.org/10.1016/j.jhevol.2022.103291.
Kuhlwilm M et al. (2016) Ancient gene flow from early modern humans into Eastern Neanderthals. Nature 530, 429–433, https://doi.org/10.1038/nature16544.
Line P (2021) Developments in paleoanthropology. Journal of Creation 35, 116–124.
Line P (2022) Developments in paleoanthropology no. 2. Journal of Creation 36, 89–98.
Lorente-Galdos B et al. (2019) Whole-genome sequence analysis of a Pan African set of samples reveals archaic gene flow from an extinct basal population of modern humans into sub-Saharan populations. Genome Biology 20, 77, https://doi.org/10.1186/s13059-019-1684-5.
Marshall M (2021) New human species has been named Homo bodoensis – but it may not stick. NewScientist, vom 28.10.2021, https://www.newscientist.com/article/2295406-new-human-species-has-been-named-homo-bodoensis-but-it-may-not-stick/.
Metz L, Lewis JE & Slimak L (2023) Bow-and-arrow, technology of the first modern humans in Europe 54,000 years ago at Mandrin, France. Sci. Adv. 9, 8, doi: 10.1126/sciadv.add4675.
Meyer M et al. (2016) Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins. Nature 531, 504–507, prepub.: https://www.eva.mpg.de/documents/Nature/Meyer_Nuclear_Nature_2016_2259387.pdf.
Posth C et al. (2017) Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals. Nat. Commun. 8, 16046, https://doi.org/10.1038/ncomms16046.
Ragsdale AP et al. (2023) A weakly structured stem for human origins in Africa. Nature, https://doi.org/10.1038/s41586-023-06055-y.
Reich D et al. (2010) Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468, 1053–1060, https://doi.org/10.1038/nature09710.
Roksandic M et al. (2022) Resolving the “muddle in the middle”: The case for Homo bodoensis sp. nov. Evol. Anthropol., doi: 10.1002/evan.21929.
Roksandic M et al. (2022) Homo bodoensis and why it matters. Evol. Anthropol., doi: 10.1002/evan.21954.
Sarmiento SS & Pickford M (2022) Muddying the muddle in the middle even more. Evol. Anthropol. 31, 237–239, https://doi.org/10.1002/evan.21952.
Schlott K (2021) »Drachenmensch«: Neue, längst bekannte Menschenform? Spektrum.de, vom 28.06.2021, https://www.spektrum.de/news/drachenmensch-neue-laengst-bekannte-menschenform/1888939.
Schlott K (2023) Homo sapiens in Europa: Kamen die ersten Pioniere viel früher? Spektrum.de, vom 23.02.2023, https://www.spektrum.de/news/homo-sapiens-in-europa-kamen-die-ersten-pioniere-viel-frueher/2112693.
Scholl B (2020) Frauen mit „Neandertal-Genvarianten“ haben weniger Fehlgeburten. Stud. Integr. J. 27, 126.
Scholl B (2022) Homininen-Schädel: „Stolpersteine“ des Grundtypmodells? Eine schöpfungstheoretische Deutung der Funde von Dmanisi. W+W Special Paper B-22-1, https://www.wort-und-wissen.org/artikel/homininen-schaedel/.
Scholl B (2023) Schöpfungsperspektive gestärkt. factum 23/1, 34f.
Slimak L et al. (2022) Modern human incursion into Neanderthal territories 54,000 years ago at Mandrin, France. Sci. Adv. 8, 6, doi: 10.1126/sciadv.abj9496.
Slon Vet al. (2018) The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature 561, 113–116, https://doi.org/10.1038/s41586-018-0455-x.
Vespasiani DM et al. (2022) Denisovan introgression has shaped the immune system of present-day Papuans. PLoS Genet. 18, e1010470, https://doi.org/10.1371/journal.pgen.1010470.
Anmerkungen:
[1] Die Schädelreste von Apidima 1 aus Griechenland wurden von Harvati et al. (2019) auf ca. 210.000 rJ datiert und zudem digital rekonstruiert. Sie besitzen einen Mix von Merkmalen des Homo sapiens sowie von älteren Menschenformen (ebd.). Es fehlen typische Neandertalermerkmale und die größte Nähe in multimetrischen Analysen besteht zu den modernen Menschen Nazlet Khater 2 und Dolní Věstonice 3 statt zum mittelpleistozänen Homo. Daher sprechen sich die Autoren für eine Zugehörigkeit zum frühen modernen Menschen aus. Es sind aber nicht alle Wissenschaftler völlig von der Anwesenheit von Homo sapiens in Griechenland vor ca. 210.000 rJ überzeugt; so halten Slimak et al. (2022, 1) dies nur für „möglich“ („possibly“), aber nicht für eindeutig belegt.
[2] Die morphologischen (körperbaulichen) Belege von Homo sapiens in Frankreich beruhen auf einem einzigen Backenzahn (M2 oben rechts) aus der Grotte Mandrin, der dem archaischen Homo sapiens und nicht den anderen dortigen Neandertaler-Zähnen gleichen soll (vgl. Slimak et al. 2022; Schlott 2023). Die Funde wurden auf 56.800 und 51.700 rJ datiert (ebd.). Die in der Studie als Belege angeführten multimetrischen Analysen (Slimak et al. 2022, 3, Fig. 6+7, S20–S22) beziehen sich nur auf die Form des Kronenumrisses sowie des Schmelz-Dentin-Übergangs (EDJ). Außerdem wurde bei den 3D-Rekonstruktionen, die die Mehrheit der Analysen ausmachen (Fig. 6+7, S20–22), mit rekonstruierten Werten für die Spitzen der Dentinhöcker gearbeitet, da diese dem betreffenden unvollständigen Backenzahn fehlen (vgl. Slimak et al. 2022, 4, Fig. S13+S14). Insgesamt liegt der Backenzahn in keiner multimetrischen Analyse im Variationsbereich des Neandertalers. Der Zahn ist zwar mehrheitlich im Variationsbereich des oberpleistozänen modernen Menschen angesiedelt und ähnelt diesem im Vergleich mit Neandertalern und Homo sapiens aus dem Holozän (11.700 rJ bis heute) auch am stärksten, dennoch erweist er sich in vier von neun Analysen als einzigartig (ebd., Fig. 6+7, vgl. S20–S22). Außerdem wurden die fossil älteren Funde von Homo heidelbergensis und Homo erectus in den Vergleich gar nicht einbezogen, sodass sowieso morphologisch keine zwingende Zugehörigkeit zu Homo sapiens nachgewiesen werden kann (s. o.). Als zweiter Beleg für eine Zugehörigkeit zum modernen Menschen werden kleine Steinwerkzeugspitzen aus der Fundschicht des Backenzahns angeführt, die laut experimentellen Studien mutmaßlich Pfeilspitzen waren, die entsprechenden Funden aus dem Nahen Osten ähneln (vgl. Schlott 2023; Slimak et al. 2022; Metz et al. 2023). Letztere stammen nach bisherigem Wissen vom modernen Menschen (ebd.). Insgesamt sind nicht alle Paläoanthropologen (wie Jean-Jacques Hublin) von den vorgestellten Belegen für Homo sapiens vor ca. 54.000 rJ in der Grotte Mandrin überzeugt (Schlott 2023).
[3] Keeling et al. (2023, 14) nennen folgende Studien: Hernandez-Pacheco & Obermaier 1915; Bonarelli 1916; Sergi 1917; Keith 1931; Hoyos-Sainz 1947; de Lumley, 1971–1972; Roth 1982; Sanchez-Lopez 1993; Roth et al. 1993; Rosas 1993.
[4] Bei einer multivariaten PC-Analyse von PC 1 und PC 2 lag Banyoles sogar ausschließlich im Bereich heutiger – merkwürdigerweise aber nicht fossiler – Unterkiefer von Homo sapiens.
[5] UR 501, OH 7, OH 13, KNM-ER 730, Dmanisi, D211 and D2735, ATE9-1, Sangiran 9 und 22.
[6] Allerdings besitzt bspw. der Unterkiefer von Nesher Ramla mit ca. 140.000 rJ zwar ein paar Gemeinsamkeiten mit Banyoles, zeigt aber doch mehr Ähnlichkeiten mit Neandertalern (S. 14f).
[7] Das Mittelpleistozän wird auch Chibanium genannt und reicht von ca. 781.000 bis 126.000 rJ.
[8] Homo altaiensis (Denisova), antecessor, bodoensis, cepranensis, heidelbergensis, helmei, longi, mauritanicus, narmadensis, njarasensis, rhodesiensis, saldanensis, steinheimensis und tsaichangensis.
Autor dieser News: Benjamin Scholl
© 2023, http://www.genesisnet.info/schoepfung_evolution/n314.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.03.23 Australische DNA in Südamerika
Seltsame Wege der Migration oder falsche Annahmen über genetische Mutationen?
Je tiefer man in die Genetik des Menschen eindringt, umso schwieriger wird es die Geschichte der Menschheit zu rekonstruieren. Statt klarer zu werden, wird sie eher undurchsichtiger. Vor allem die Möglichkeit von genetischen Analysen von DNA, die aus den Knochen von Neandertalern und fossil erhaltenen Denisova-Menschen aus Sibirien isoliert wurden, hat für einen Paradigmenwechsel gesorgt. Neue genetische Funde in alten Knochen von Menschen aus Südamerika werfen weitere Fragen über unsere Herkunft auf. Wie sind die genetischen Fingerabdrücke von Neandertalern, verschiedenen Denisova-Menschen und sogar australischen Signaturen dorthin gelangt? Erfolgte dies durch Migration oder gibt es eine andere Erklärung?
Vor dem Aufkommen der sogenannten Paläogenetik glaubte die Mehrheit der Forscher, dass sich Neandertaler nie mit modernen Menschen gekreuzt haben, sondern man sah sie als ausgestorbenen Seitenzweig der Menschheit. Man vermutete, dass sich die Abstammungslinien des modernen Menschen und des Neandertalers vor der Entstehung des heutigen Menschen auseinanderentwickelten (Noonan 2010). Außerdem schienen die frühen paläogenetischen Analysen der mitochondrialen DNA dies zu bestätigen.
Diese Ansicht wurde bis zum Jahr 2010 vertreten, als der Paläogenetiker Svante Päböö nachwies, dass moderne Menschen, insbesondere Europäer und Asiaten, einen geringen Anteil an Neandertaler-DNA besitzen. Das bedeutet, dass in Neandertalern Varianten von Genen (Allele) vorhanden sind, die man auch bei heute lebenden Menschen findet. Für die Entwicklung der paläogenetischen DNA-Forschung erhielt Päböö im Jahr 2022 den Nobelpreis für Medizin. Der genetische Beitrag der Neandertaler zum Genpool des modernen Menschen wird heute allgemein vor allem mit Europäern, insbesondere Nordeuropäern, in Verbindung gebracht. Denisova-Menschen hingegen werden im Allgemeinen mit Populationen in Asien verbunden.
Zur Überraschung der Biologen wurde jedoch nachgewiesen, dass auch afrikanische Menschen Genvarianten besitzen, die auch bei Neandertalern zu finden sind (Price 2020). Man war nämlich bislang davon ausgegangen, dass die Neandertaler Afrika schon vor längerer Zeit verließen und nie zurückkehrten, um sich mit den zurückgebliebenen Menschen zu vermischen. Sie seien also ihren eigenen evolutionären Weg unabhängig von den afrikanischen Menschen gegangen. Eine ähnliche Überraschung wurde neulich in der DNA von indigenen Völkern Südamerikas aufgedeckt.
Eine Gruppe brasilianischer Archäologen und US-amerikanischer Genetiker publizierte ihre Befunde zu der genetischen Abstammung einiger der frühesten Menschen Südamerikas in den Proceedings der Royal Society (Campelo dos Santos 2022). Die Wissenschaftler isolierten DNA aus alten menschlichen Genomen aus mindestens ca. 900 Jahre alten Knochen, die an archäologischen Stätten im Nordosten Brasiliens, Panamas und Uruguays gefunden wurden, und verglichen die Sequenzen mit Genomdatenbanken heutiger und ausgestorbener Menschen. Ursprünglich wollten die Forscher, die an diesen Stätten arbeiteten, die Muster alter Wanderungen auf dem gesamten südamerikanischen Kontinent kartieren. Dabei bestätigte sich, dass die Wanderungen entlang des Kontinents überwiegend von Norden nach Süden verliefen (Micu 2023; Campelo dos santos et al. 2022, Fig. 5).
Erstaunlicherweise wies die untersuchte DNA der Menschen aus Südamerika aber darauf hin, dass mehrere Individuen aus Uruguay und Panama offenbar von Denisova-Menschen und von Neandertalern abstammen. Dabei hatten zwei Individuen, die vor ca. 500–700 Jahren gelebt haben (PAPV173 und CH19B; vgl. ebd., Fig. 2), mehr Denisova-Anteile, während bei den anderen südamerikanischen Genomen der Neandertaler-Anteil überwog. Ein höherer Denisova-Anteil kommt auch bei Melanesiern in Südostasien vor: Sie haben ca. 4,8% Denisova-DNA und ca. 2,5 % Neandertaler-DNA (Brandt 2020, 226). Vielleicht teilen sie also einen gewissen Anteil gemeinsamer Vorfahren aus Ostasien, die mehr Denisova-Allele in sich trugen, mit manchen Indianervölkern?
Und als ob das noch nicht genug wäre, fanden sie auch noch eine starke australische genetische Signatur in einem ca. 500 Jahre alten Genom aus Panama (Micu 2023; Campelo dos Santos et al. 2022).
Es ist das erste Mal, dass Denisova- und Neandertaler-DNA in alten südamerikanischen Populationen nachgewiesen wurde. Diese Menschen werden daher als „Vorfahren“ interpretiert. Andre Luiz Campelo dos Santos, ein Archäologe an der Florida Atlantic University und Hauptautor der Studie, sagte: „Das Vorhandensein dieser Vorfahren in alten indianischen Genomen kann durch Episoden der Kreuzung zwischen anatomisch modernen Menschen und Neandertalern und Denisova-Menschen erklärt werden, die Jahrtausende vor der Ankunft der ersten menschlichen Gruppen in Amerika über die Beringstraße stattgefunden haben sollten.“ Die Autoren kamen zu dem Schluss, dass diese Befunde Licht auf die komplizierte demografische Geschichte des östlichen Südamerikas wirft und einen Ausgangspunkt für künftige kleinräumige Studien auf regionaler Ebene bietet (Campelo dos Santos et al. 2022).
Bringen diese überraschenden Daten wirklich Licht ins Dunkel der menschlichen Geschichte und der Ausbreitung der Menschheit? Die Forscher stellen fest, dass die Ergebnisse unser Verständnis des genetischen Hintergrunds der alten südamerikanischen Völker verkomplizieren (Micu 2023; Campelo dos Santos et al. 2022). Die populärwissenschaftliche Presse kommentierte: „Alte Völker in Südamerika hatten sowohl Denisovaner als auch Neandertaler-DNA – und wir haben keine Ahnung, wie sie dorthin gekommen ist“ (Micu 2023).
Abstammung und Kreuzung oder wiederkehrende Mutationen?
Haben wir es bei den Menschen aus Südamerika aber wirklich mit alter DNA von Neandertalern und Denisova-Menschen zu tun, oder sogar mit australischer DNA? Könnte es stattdessen sein, dass die Annahme über die zugrunde liegenden Mutationen falsch sind?
Den vorgebrachten Schlussfolgerungen der Autoren liegt die Annahme zugrunde, dass Mutationen generell weder vorhersehbar sind noch mit dem Verhalten, dem Lebensstil oder den Umweltbedingungen zusammenhängen. Mutationen sind demnach reine Zufallsereignisse in DNA-Sequenzen. Diese Ansicht wird von Futuyma (2005, 178f) in seinem weltweit verbreiteten Lehrbuch der Evolutionsbiologie wie folgt dargestellt:
„Obwohl wir die Wahrscheinlichkeit vorhersagen können, dass eine irgendeine Mutation auftreten wird, können wir nicht vorhersagen, welche von einer großen Anzahl an Genkopien die Mutation durchlaufen wird.“
Nach dem gegenwärtigen Konsens treten alle Mutationen nach dem Zufallsprinzip auf und daher in der Regel nur einmal vor – mit Ausnahme einiger „Hotspots“. Neuere Daten legen jedoch nahe, dass diese allgemeine Annahme falsch ist: Gleichartige Mutationen treten an Hotspots häufig auf und können so eine Illusion von Abstammung hervorrufen (Borger 2019). Dass dies bei den genetischen Befunden der untersuchten Menschen aus Südamerika eine Rolle spielen könnte, geht aus mehreren Beobachtungen hervor.
Zunächst einmal weisen die Allele (Erscheinungsformen) der betreffenden Gene in der Regel nur eine Punktmutation auf. Und es handelt sich um Allele, die man auch bei modernen Menschen vorfindet. Die australasiatischen Allele wurden außerdem zuvor auch im Südosten Brasiliens gefunden und sind heute im Volk der Surui (auch bekannt als Paiter) in Amazonien vorhanden (Micu 2023; Campelo dos Santos et al. 2022).
Das Vorkommen des australischen Allels in Südamerika lässt vermuten, dass man es möglicherweise mit wiederkehrenden Mutationen zu tun hat. Mit anderen Worten: Sie könnten als genetische Hotspots interpretiert werden, die unabhängig voneinander in verschiedenen Populationen entstanden sind, über Generationen vererbt wurden und heute eine Illusion von Abstammung erzeugen. Die Hotspot-Mutationen können also als Konvergenzen (d. h. als unabhängig vorkommend) verstanden werden, ein Phänomen, das zunehmend in biologischen Systemen zu beobachten ist. Es liegt auf der Hand, dass solche Mutationen nicht nur die Rekonstruktion von Abstammungslinien erschweren oder gar verunmöglichen, sondern auch die Vorstellungen über Migrationsrouten der Menschen über die Erde verzerren.
Quellen
Borger P (2019) Artübergreifende wiederkehrende Mutationen. Oder: Die Illusion der Verwandtschaft Stud. Integr. J. 26, 77–87.
Brandt M (2020) Wie alt ist die Menschheit? Demographie und Steinwerkzeugbefunde mit überraschenden Befunden. 6. erw. Aufl. SCM Hänssler, Holzgerlingen.
Campelo dos Santos AL, Owings A, Sullasi HSL, Gokcumen O, DeGiorgioM & Lindo J (2022) Genomic evidence for ancient human migration routes along South America’s Atlantic coast. Proc. R. Soc. B289: 20221078, https://doi.org/10.1098/rspb.2022.1078.
Futuyma DJ (2005) Evolutionary Biology, 3rd ed., Sinauer Associates, Sunderland, MA, S. 178–179.
Micu A (2023) Ancient peoples in South America had both Denisovan and Neanderthal DNA — and we have no clue how it got there. https://www.zmescience.com/science/south-america-ancient-neanderthal-denisovan-dna-92352342/
Noonan JP (2010) Neanderthal genomics and the evolution of modern humans. Genome Res. 20, 547–553; doi:10.1101/gr.076000.108.
Price M (2020) Africans carry surprising amount of Neanderthal DNA. doi: 10.1126/science.abb0984.
Autor dieser News: Peter Borger
© 2023, http://www.genesisnet.info/schoepfung_evolution/n311.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
09.01.23 Beobachtungen bei Schimpansen widersprechen „Savannen-Hypothese“
Eine Studie von Drummond-Clarke et al. (2022, 1) ist dem Ziel gewidmet, anhand der Beobachtung heute lebender Schimpansen die Plausibilität der berühmten „Savannen-Hypothese“ zu prüfen, die sich bis heute in Fachliteratur und Schulbüchern findet. Demnach soll der Erwerb des zweibeinigen Ganges durch eine baumärmere Savannen-Umgebung begünstigt gewesen sein. Die Ergebnisse der Autoren widersprechen der Savannen-Hypothese jedoch deutlich.
Die Savannenhypothese
Die bekannte „Savannen-Hypothese“ besagt, dass sich im späten Miozän bis Pliozän (ca. 10 bis 2,5 MrJ [Mio. radiometrische Jahre]) die Wälder in Afrika zurückgezogen haben sollen, was die Homininen (vermeintliche Vormenschen seit Abspaltung der Schimpansen-Linie) dazu gebracht hätte, sich viel häufiger zweibeinig fortzubewegen (Drummond-Clarke et al. 2022, 1). Auf diese Weise sei schrittweise der zweibeinige menschliche Gang entstanden. So heißt es zum Beispiel im aktuellen Oberstufen-Biologiebuch Biosphäre Gesamtband SII (Becker 2020, 509): „Aus dem ursprünglich zusammenhängenden Regenwald entwickelte sich eine Savannenlandschaft mit Galeriewäldern, in der die Nahrung ungleichmäßig verteilt war. Genau in dieser Region befinden sich die meisten Fundstellen aufrecht gehender Vormenschen. Obwohl sie sich weniger schnell fortbewegen konnten als Vierbeiner, mussten sie in dieser Landschaft dennoch einen Vorteil gehabt haben. So wurde berechnet, dass sich mit der zweibeinigen Fortbewegung bei vergleichbarem Energieaufwand [mindestens] doppelt so weite Strecken zurücklegen lassen wie mit vierbeiniger Fortbewegung.“ Ähnlich schreibt der Professor für Biologiegeschichte Thomas Junker (2021, 16): „Bei Menschen dagegen sind die abweichenden Merkmale entstanden, weil sie sich an andere ökologische Bedingungen – an das Leben in Baum- und Grassavanne – angepasst haben.“

Abb. 1: Die Savannen-Hypothese besagt, dass durch einen klimabedingten Rückgang der Wälder Homininen (wie z. B. Australopithecus) dazu gezwungen gewesen sein könnten, den aufrechten Gang zu entwickeln. Dies widerspricht empirischen Befunden bei heute lebenden Schimpansen. (Symbolbild nach Pixabay)
Drummond-Clarke et al. (2022) halten zu Beginn ihres Artikels allerdings erst einmal fest, dass keine direkten fossilen Belege für die Savannen-Hypothese existieren, und dass es Schwierigkeiten dabei gibt, „die Beziehung zwischen Verhalten und Lebensraum allein aus der Morphologie [Körperbau] zu rekonstruieren“. Daher halten sie verhaltensbiologische Studien bei Schimpansen als den nächstverwandten lebenden Arten für den entscheidenden „Schlüssel“ zum Verständnis zur Evolution des aufrechten Ganges (S. 1).
Die Freilandbeobachtungen bei Schimpansen
Die Wissenschaftler unternahmen einen Test dieser Hypothese, indem sie das Verhalten einer Schimpansen-Gruppe untersuchten, die sich erst seit 2018 in der Issa-Talregion (im Westen Tansanias) angesiedelt hatte. Diese Region enthält ein sogenanntes Savannen-Mosaik aus offenem woodland und kleinen geschlossenen Waldregionen (forest). Die Landschaft soll darin dem Lebensraum der frühen Homininen wie Orrorin, Ardipithecus ramidus und Australopithecus afarensis vergleichbar gewesen sein (S. 2). Übrigens soll auch die Ernährung von Bäumen bei Homininen vor ca. 4 MrJ der Ernährung heutiger Savannen-Schimpansen geähnelt haben (Drummond-Clarke et al. 2022, 1+5).
Die Forscher konnten ca. 13.700 Beobachtungen von 13 erwachsenen Schimpansen aufzeichnen (S. 3). Dabei fanden sie heraus, dass die Schimpansen sich in offener Vegetation „signifikant“ häufiger am Boden fortbewegten als in geschlossener Vegetation (82 % vs. 61 % der Fortbewegung). Die Sensation war, dass Zweibeinigkeit hauptsächlich in Bäumen (zu 86 %) – also beim Klettern – aber nur zu 14 % am Boden auftrat. Schimpansen nutzen Zweibeinigkeit am Boden in der Savanne also erstaunlich selten. Meistens war die Zweibeinigkeit mit der Nahrungssuche assoziiert (73 % aller Beobachtungen). Ein Vergleich mit Schimpansen aus anderen Lebensräumen (in Mahale, Gombe und Bwindi) ergab weiterhin (S. 4): „Issa hat den höchsten Prozentsatz von zweibeinigem Verhalten zum Zweck der Fortbewegung, aber es scheint keinen Zusammenhang zwischen der Gesamthäufigkeit der Zweibeinigkeit und dem Lebensraumtyp zu geben.“
Insgesamt waren die Schimpansen Issa nur in 0,77 % der Zeit mit zweibeinigem Stehen oder Gehen beschäftigt. Davon machte das zweibeinige Gehen wiederrum nur ein Viertel der Fälle aus, weshalb sich die Schimpansen insgesamt nur in ca. 0,2 % der Zeit zweibeinig fortbewegten. Dies widerspricht den Erwartungen der Savannen-Hypothese, wie auch die Autoren konstatieren (S. 5): „Unsere Ergebnisse stellen die seit Langem bestehende Assoziation zwischen zunehmender Terrestrialität [Leben am Boden] und der Evolution des Zweibeinertums als Fortbewegungsverhalten bei frühen Homininen in Frage. […] Issa-Schimpansen blieben in hohem Maße baumbewohnend und nutzten in offener Vegetation nicht mehr Zweibeinigkeit […].“
Dementsprechend hat sich die Zweibeinigkeit der Homininen nicht in der Savanne entwickelt – die Autoren vermuten stattdessen eine Entstehung in den Bäumen (S. 4). Dies ist eine Theorie, die seit dem Fund von Ardipithecus ramidus häufiger diskutiert wird – obwohl ähnliche Anpassungen an Zweibeinigkeit (sofern diese nicht fehlinterpretiert wurden; vgl. Scholl 2022c, 23) bei anderen miozänen Affen als Parallelevolution zur Entwicklung des Menschen gedeutet werden (vgl. Hartwig-Scherer 2011; Scholl 2022c, 9, 12, 19, 26). Es sei außerdem angemerkt, dass beim balancierenden Stehen oder Laufen in Bäumen ganz andere biomechanische Ansprüche und Selektionsdrücke im Detail angenommen werden müssen als beim ausdauernden, schreitenden menschlichen Gang am Boden. Dieser Sachverhalt untermauert die schon früher gemachte Feststellung: „Somit ist nach Prahabat et al. (2021) noch kein bisschen klar, wann und wie überhaupt ein Übergang „vom Baumleben zur dauerhaften Zweibeinigkeit‘ stattgefunden haben könnte.“ Außerdem war H. erectus auch als ältester Hominine eindeutig an einen gewohnheitsmäßigen menschlichen Gang angepasst (vgl. Ruff 2009; Brandt 2017a).“ (Scholl 2022a)
Ein weiterer Versuch zur Rettung der Savannen-Hypothese ist das Argument, dass Schimpansen nur begrenzt als Modell dienen könnten (vgl. Vieweg 2022), weil man voraussetzt, dass der letzte gemeinsame Vorfahr von Schimpanse und Mensch dem Schimpansen gar nicht so sehr geähnelt habe. Allerdings ist unter Wissenschaftlern sehr umstritten, wie der letzte gemeinsame Vorfahr überhaupt zu rekonstruieren sei (vgl. Scholl 2022c, v. a. 4f); und auch Drummond-Clarke et al. (2022, 1+5) betonen die Eignung von Schimpansen als den besten Test für die Savannen-Hypothese.
Schlussfolgerungen
Überaus bemerkenswert ist die Einschätzung von Drummond-Clarke et al. (2022, 5) über die Unsicherheiten in der vermeintlichen Evolution des aufrechten Ganges: „Unklar bleibt jedoch, welche Art von Selektionsdruck auf die Homininen aufgrund dieses Übergangs in offene Lebensräume wirkte, da die gegensätzlichen Signale von Terrestrialität und Arborealität [Baumleben] viel Unsicherheit darüber lassen, wie genau die Homininen diese Lebensräume nutzten […]. Mit anderen Worten, die bloße Anwesenheit von Homininen sagt nichts darüber aus, wie sie in ihren Paläohabitaten [Lebensräumen] interagierten. Darüber hinaus zeigen biomechanische Modelle […] und die innere Knochenstruktur […] eine größere Variation im Haltungsrepertoire der Homininen als bisher angenommen.“
Außerdem schließen die Autoren aus ihren Beobachtungen, dass die Anpassungen an das Baumleben bei „vielen frühen“ (z. B. Sahelantropus, Orrorin, Ardipithecus und Australopithecus afarensis) und „sogar späten“ Homininen (sie nennen Australopithecus sediba, den vermeintlichen „Homo“ namens naledi und den rätselhaften Homo floresiensis) „funktionell bedeutende“ Anpassungen waren (S. 1+5). Zu diesen fossil untersuchbaren Körpermerkmalen, die als funktional bedeutende Kletteranpassungen zu werten sind, zählen die Autoren gebogene Zehen- und Fingerglieder, lange obere Gliedmaßen sowie bewegliche Gelenke in Schulter, Ellbogen und Handgelenk (S. 1).
Die tatsächlichen Beobachtungen bei Schimpansen sprechen damit gegen die von vielen Evolutionsbiologen favorisierte Savannen-Hypothese zum Erwerb des aufrechten Ganges. Exemplarisch wird hier deutlich, dass die Savannen-Hypothese eine evolutionär – und im Wesentlichen eben nicht empirisch – motivierte Spekulation darüber darstellt, wie der menschliche Gang hätte entstanden sein können. Wie oben von Drummond-Clarke et al. (2022, 1) beschrieben wurde, wird der spekulative Charakter daran ersichtlich, dass direkte fossile Beweise und eindeutige Beziehungen zwischen Körperbau, Verhalten, Lebensraum bei der Savannen-Hypothese fehlen. Stattdessen werden mittels „story telling“ (eine „Geschichte erzählen“) theoretische Spekulationen über Knochenfunde an fossilen Homininen und deren ökologisches Umfeld mit plausibel klingenden – aber oft rein hypothetischen – Wirkungen von Evolutionsfaktoren zu einer gut klingenden Geschichte verbunden (vgl. Scholl 2022c, v. a. 23f). Wird diese Geschichte oft genug in der Fachliteratur und in populären Darstellungen wiederholt, weil plausiblere Alternativen innerhalb des evolutionären Paradigmas fehlen, setzt sie sich schließlich auch in Schulbüchern und den Köpfen fest.
Aufgrund der Beobachtungen von Schimpansen in der Savanne bleibt festzuhalten: Die Entstehung des ausdauernd schreitenden Ganges bei echten Menschen, wie er fossil seit Homo erectus nachweisbar ist, ist weiterhin ungeklärt (vgl. Brandt 2017a; Scholl 2022b, S. 9f, 14, 18; Scholl 2022c, 14f, 22, 25f, 29; Brandt 2023, in Vorbereitung). So konstatiert auch Vieweg (2022) auf wissenschaft.de zum vorliegenden Artikel von Drummond-Clarke und Kollegen: „Nach wie vor bleibt ein Rätsel, warum die Vorfahren des Menschen schließlich eine aufrechte Fortbewegungsweise annahmen.“
Literatur
Becker J (Hrsg.) (2020) Biosphäre Sekundarstufe, II Gesamtband. 1. Aufl. Cornelsen Verlag Berlin.
Brandt M (2017a) Wie sicher sind Deutungen in der Paläanthropologie? Australopithecus sediba und sein merkwürdiges Merkmalsmosaik. In: Brandt M (Hg) Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special. SCM Hänssler, S. 9–49.
Brandt M (2017b) Frühmensch war ein „Missing Link“? Die Schulter von Homo erectus. In: Brandt M (Hg) Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special. SCM Hänssler, S. 93–103.
Brandt M (2023) Australopithecus ein effizienter Zweibeiner? W+W Special Paper, in Vorbereitung.
Drummond-Clarke RC et al. (2022) Wild chimpanzee behavior suggests that a savanna-mosaic habitat did not support the emergence of hominin terrestrial bipedalism. Aci. Adv. 8, eadd9752, doi: 10.1126/sciadv.add9752.
Hartwig-Scherer S (2011) Ardipithecus: Ein Astgänger sägt am Lehrbuchwissen. Stud. Integr. J. 18, 68–77.
Junker T (2021) Die Evolution des Menschen. 4. Aufl. C.H.Beck.
Prabhat AM et al. (2021) Homoplasy in the evolution of modern human-like joint proportions in Australopithecus afarensis. eLife 10:e65897, https://doi.org/10.7554/eLife.65897.
Ruff C (2009) Relative Limb Strength and Locomotion in Homo habilis. Am. J. Phys. Anthropol. 138, 90–100, doi: 10.1002/ajpa.20907.
Scholl B (2022a) Genesisnet, Eine modern menschliche Wirbelsäule bei Australopithecus sediba?, https://www.genesisnet.info/index.php?News=298.
Scholl B (2022b) Homininen-Schädel: „Stolpersteine“ des Grundtypmodells? Eine schöpfungstheoretische Deutung der Funde von Dmanisi. W+W Special Paper B-22-1, https://www.wort-und-wissen.org/artikel/homininen-schaedel/.
Scholl B (2022c) „Totales Chaos“: Unklare Abstammungsverhältnisse bei Menschenaffen und Menschen. W+W Special Paper B-22-2, https://www.wort-und-wissen.org/artikel/miozaene-affen/https://www.wort-und-wissen.org/artikel/miozaene-affen/.
Vieweg M (2022) Aufrechter Gang: Fragliche Ursprungs-Annahme. wissenschaft.de, veröffentlicht am 14.12.2022, https://www.wissenschaft.de/erde-umwelt/aufrechter-gang-fragliche-ursprungs-annahme/.
Autor dieser News: Benjamin Scholl
© 2023, http://www.genesisnet.info/schoepfung_evolution/n310.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
20.10.22 Nobelpreis für Forschungen über das Erbgut aus menschlichen Fossilien
Von Harald Binder & Peter Borger
Svante Pääbo, ein schwedischer Biowissenschaftler, der einen großen Teil seiner wissenschaftlichen Karriere in deutschen Forschungseinrichtungen verbracht hat, ist mit dem Nobelpreis ausgezeichnet worden. In der Öffentlichkeit ist Pääbo bekannt für seine Beiträge zur Erforschung des Erbguts des Neandertalers. In populären Medien ist viel davon zu lesen, dass damit die Evolution des Menschen besser verstanden sei. In einer kritischen Würdigung der Arbeiten von Pääbo wird hier darauf verwiesen, dass die genetischen Daten die Neandertaler im Variationsbereich des modernen Menschen (Homo sapiens) verorten (eine Position, die in der Paläanthropologie unabhängig von genetischen Daten vertreten wird). Die genetischen Daten aus menschenähnlichen Fossilien können auch aus einer Schöpfungsperspektive sinnvoll interpretiert werden.

Abb. 1 Svante Pääbo (Wikimedia: Duncan.Hull, CC BY-SA 4.0)
Der Nobelpreis für Physiologie und Medizin 2022 wurde dem schwedischen Immunologen Svante Pääbo (*1955 in Stockholm) zugesprochen. Das Nobelkomitee zeichnet den Wissenschaftler aus „für seine Entdeckungen, die das Erbgut ausgestorbener menschenähnlicher Lebewesen und die Evolution des Menschen betreffen“1. Svante Pääbo ist ein Sohn des schwedischen Mediziners und Biochemikers Sune Bergström (1916–2004), der 1982 gemeinsam mit zwei anderen Wissenschaftlern (B. I. Samuelsson und J. R. Vane) den Nobelpreis für Physiologie und Medizin für bahnbrechende Arbeiten über Prostaglandine erhalten hatte. Der Familienname geht auf die Mutter, die estnische Chemikerin, Karin Pääbo zurück.
Für seine Promotion bearbeitete Svante Pääbo immunologische Fragestellungen an der Universität Uppsala. Sein eigentliches Interesse galt jedoch der Ägyptologie und allgemein der Geschichte des Menschen. So besorgte er sich ohne Kenntnis seiner Betreuer Proben von ägyptischen Mumien und begann darin nach Spuren von ihrer DNA zu suchen. Seine erste Arbeit über Hinweise auf Überreste von DNA in ägyptischen Mumien veröffentlichte er in einer Zeitschrift, die in der DDR herausgegeben wurde. Diese Arbeit fand allerdings keine Beachtung. Ganz im Gegensatz zu einem Beitrag, der ein Jahr später von der renommierten Zeitschrift „Nature“ akzeptiert wurde. Sogar das Titelbild der Ausgabe wurde dem Thema gewidmet.
Nach kurzen Forschungsaufenthalten in Zürich und London trat Pääbo eine Postdoc-Stelle im Labor von Allan Wilson in Berkeley an, wo er die Technik der Polymerase-Kettenreaktion (PCR) nutzte – eine Methode, um DNA-Moleküle durch einen Kopiermechanismus zu vervielfältigen – um Erbinformation aus den fossilen Überresten ausgestorbener Tiere zu gewinnen. So konnte Pääbo zusammen mit anderen Autoren Daten zum Erbgut z. B. vom Quagga, Riesenfaultier und Mammut publizieren. Den Großteil seines Forscherlebens verbrachte Svante Pääbo in deutschen Forschungseinrichtungen: an der LMU in München (1990–1997) und seit 1997 am Max-Planck-Institut für evolutionäre Anthropologie in Leipzig, wo er als Direktor die Abteilung Evolutionäre Genetik leitet.
Paläogenetik
Mit den Untersuchungen von DNA-Bruchstücken mit Erbinformationen mumifizierter Überreste von Menschen hatte Pääbo den Grundstein für einen neuen Forschungsbereich gelegt, der Paläogenetik. In der Paläogenetik, also für die Untersuchung von Erbinformationen alter Lebewesen, sind keine heute lebenden Organismen verfügbar, sondern man ist auf Präparate aus Museumsbeständen oder auf entsprechende Fossilien angewiesen. Aufgrund der Geschichte der Proben, wie den an der Fossilisierung beteiligten Prozessen, ihrer Präparation und sonstigen Handhabungen enthalten die Proben wenig und massiv veränderte DNA neben einer unbestimmten Menge an DNA, die aus anderen Quellen stammt.
Mit den Mitarbeitern seiner Arbeitsgruppen in München und Leipzig, z. T. auch mit internationaler Zusammenarbeit, konnte Pääbo zunächst DNA-Sequenzen aus Mitochondrien des Neandertalers bestimmen. Diese mitochondriale DNA (mtDNA) konnte in Proben aus dem namengebenden Typusexemplar des Neandertalers gewonnen werden. Die gewonnen Daten interpretierten die Autoren in dem Sinne, dass die Neandertaler ausgestorben waren, ohne genetische Spuren im modernen Menschen hinterlassen zu haben. Dies wurde oft in dem Sinne aufgenommen, als würde es sich um zwei verschiedene „menschliche Spezies“ handeln, die keine Fortpflanzungsgemeinschaft bildeten. Unter großem medialem Aufsehen wurden DNA-Sequenzen aus Proben von weiteren Neandertalerfossilien z. B. aus Osteuropa präsentiert, und schließlich konnte 2010 anhand der analysierten Sequenzdaten ein Genom von Neandertalern – unter Nutzung bereits bekannter Genome von Menschen und Großaffen – weitgehend rekonstruiert werden. Großes Aufsehen erregte auch die Veröffentlichung von DNA-Sequenzen aus einem fossilen Fingerglied bzw. später auch einem Backenzahn, die in der Denisova-Höhle im Altai-Gebirge in Sibirien gefunden worden waren. Diese Fragmente der Erbinformation wurden von Pääbo und seinen Kollegen so interpretiert, dass es sich um einen weiteren, bisher unbekannten und sowohl vom modernen Menschen als auch vom Neandertaler verschiedenen Menschentyp handeln müsse. In weiteren Veröffentlichungen schlossen die Autoren aus Vergleichen von Sequenzdaten, dass sowohl Neandertaler als auch der Denisova-Mensch mit Homo sapiens in Fortpflanzungsgemeinschaft standen und damit derselben Art, Homo sapiens, zugehörig sind. Diese Sicht über die Neandertaler kam aber nicht unerwartet, da archäologische Erkenntnisse eine zunehmende Anzahl von Fachleuten zu derselben Einschätzung führten (vgl. z. B. Frayer und Radovčić 2022).
Sequenzvergleich einzelner Gene – gibt es Neandertaler-Gene?
Svante Pääbo und seine Mitarbeiter konnten zeigen, dass in Neandertalern Varianten von Genen (Allele) vorhanden sind, die man auch bei heute lebenden Menschen findet. Wenn sie diese Allele addieren, dann kommen die Autoren auf ca. 4 % des Genoms, in denen sich im modernen Menschen Allele finden, die auch im Neandertaler nachgewiesen werden konnten. In seinem Buch „Die Neandertaler und wir – Meine Suche nach den Urzeit-Genen“ (Pääbo 2014) prägt der Autor bereits im Untertitel die Vorstellung von Genen, die für Neandertaler typisch sind. Die Sequenzdaten zeigen aber Varianten von Genen, die sich bei modernen Menschen und bei Neandertalern finden, also Allele. Die Bezeichnung „Neandertaler-Gene“ ist daher missverständlich und intendiert spezifische Gene, wobei die Auffassung naheliegend ist, dass es sich um Varianten von Genen handelt, die Neandertalern und Menschen gemeinsam sind. Es ist wichtig, die Begriffe „Gene“ und „Allele“ (Genvarianten) auseinanderzuhalten: Ein Gen ist die Erbinformation für bestimmtes Merkmal, ein Allel ist eine Variante eines Gens.
Pääbo hat gezeigt, dass beim modernen Menschen teilweise die gleichen Allele zu finden sind wie beim Neandertaler. Es wurde festgestellt, dass einige dieser Allele mehr oder weniger mit Krankheit und Gesundheit korreliert sind (Gibbons 2016). Frei nach dem Motto: Leiden Sie unter Depressionen? Das könnte an Ihren Neandertaler-Genen liegen. Blasenschwäche? Neandertaler-Gene! Selbst einen schwereren Verlauf von Covid-19 konnte Pääbo mit „Neandertaler-Genen“ erklären (Zeberg & Pääbo 2020). Meldungen über positiv wirksame Einflüsse von „Neandertaler-Genvarianten“ gibt es zwar auch, erfahren aber weniger Aufmerksamkeit (vgl. Scholl 2020). Die Attraktivität solcher Erklärungen ist bei der Art und Weise von Diskussionen, wie sie derzeit häufig in der Öffentlichkeit geführt werden, nicht verwunderlich. Woher das „Neandertaler-Gen“ (bzw. das betreffende Allel) stammt, das für schwere Covid-Verläufe verantwortlich sein soll, bleibt ein großes Rätsel, da dieselbe Studie mit Sicherheit ausschloss, dass es von einem gemeinsamen Vorfahren stammen könnte.
DNA des modernen Menschen und des Neandertalers fast identisch
Im Jahr 2021 wurde durch den Vergleich tausender Sequenzen des modernen Menschen mit denen der Neandertaler deutlich, dass der moderne Mensch kein ganz anderes, einzigartiges Genom besitzt (Sánchez-Quinto et al. 2012). In seiner DNA-Sequenz wurden nur 1,5 % der Allele des modernen Menschen als einzigartig eingestuft (Borger 2021). Bedeutet das, dass 98,5 % der modernen Menschen Neandertaler sind? Nein, es bedeutet, dass 98,5 % der im Neandertaler-Genom gefundenen Variationen auch anderswo in der Population des modernen Menschen vorkommen. Möglicherweise erhöht sich dieser Prozentsatz, wenn man noch mehr DNA-Sequenzen einbezieht. Beim modernen Menschen ist kaum noch einzigartige DNA zu finden. Dies ist ein genetischer Beleg dafür, dass der Neandertaler – bei den bekannten Unterschieden in der bei ihm ausgeprägten Merkmalskombination – zum modernen Menschen Homo sapiens gehört.
Wenn es keine spezifischen Neandertaler-Gene gibt, sondern nur Allele, worin besteht dann der Unterschied zwischen modernen Menschen und diesen Urzeitmenschen? Das ist eine berechtigte Frage! Beim Vergleich der Gesamt-DNA innerhalb moderner menschlicher Populationen konnten genetische Unterschiede von bis zu 12 % festgestellt werden (Borger 2021). Selbst innerhalb einer Population können sich zwei Personen, die miteinander Kinder haben, um mehrere Prozent unterscheiden. Dieser enorme Unterschied, der viele Millionen DNA-Buchstaben betragen kann, ist eher das Ergebnis von Indel-Mutationen, d. h. Mutationen, bei denen sich große Abschnitte der DNA verdoppeln oder verschwinden (Duplikationen und Deletionen), als eine Folge von Punktmutationen. Diese Indel-Mutationen, die durch genetische Mechanismen angetrieben werden, liegen superschnellen Anpassungsprozessen zugrunde (Borger 2018).
Das Gen FOXP2, das beim Menschen einen Zusammenhang mit seiner Sprachfähigkeit zeigt, liegt auch beim Neandertaler vor und wird als genetischer Hinweis auf dessen Sprachfähigkeit gewertet (Krause 2007). Die von vielen Forschern dem Neandertaler zunächst nur eingeschränkt zugestandene Sprachfähigkeit war aber bereits zuvor aus verschiedenen archäologischen Fundzusammenhängen erwartet worden und spätestens seit dem Nachweis eines Zungenbeins gilt sie als gut begründet. Außerdem gibt es klare Anzeichen dafür, dass der Neandertaler wie der moderne Mensch 46 Chromosomen hatte, während alle Menschenaffen 48 Chromosomen besitzen (Miga 2017). Die gleiche Anzahl von Chromosomen, die gleichen Gene wie beim Menschen und die gleichen pathogenen Allele? Nichts Menschliches war den Neandertalern offenbar fremd.
Svante Pääbo hat mit seiner Forschung einen neuen Forschungsbereich eröffnet, in dem Erbinformationen aus Überresten von Lebewesen zugänglich gemacht worden sind. Seine Arbeitsgruppe hat viel dazu beigetragen, dass die erhaltenen Daten an Vertrauenswürdigkeit gewonnen haben. Was die Interpretation der DNA-Sequenzen aus menschlichen Fossilien betrifft, so folgt Pääbo darin unkritisch den etablierten Evolutionsvorstellungen. Dies führt dazu, dass sich das, was er als Denkrahmen voraussetzt, dann auch in seiner Interpretation der genetischen Daten wiederfindet. Er hat in seiner Forschung keinen neuen, unabhängigen Hinweis darauf gefunden, der eine Entwicklung des Menschen aus affenähnlichen Vorfahren heraus beweisen würde. Einige Sichtweisen, die durch paläoanthopologische Forschungen bereits vorhanden waren, konnte er mit seinen Ergebnissen untermauern. Insgesamt hat er vor allem mit den Ergebnissen von Neandertalern deren große genetische Ähnlichkeit zum modernen Menschen unterstrichen. Die Ähnlichkeit interpretiert Pääbo mit vielen seiner Kollegen im Sinne einer menschlichen Evolution. Dies wurde in den Darstellungen der populären Medien noch verstärkt. Es bleibt jedoch festzuhalten, dass seine Resultate dem Gedanken einer Schöpfung der Menschen nicht widersprechen und auch von diesem Standpunkt aus interpretiert und verstanden werden können. Denn lange bevor entsprechende genetische Befunde vorlagen, konnte man aus Grundtyp-Perspektive bereits davon ausgehen, dass sich moderner Mensch und Neandertaler miteinander hatten fortpflanzen können, weil sie beide als regional unterschiedlich angepasste Nachfahren der von Gott erschaffenen Menschen sind. Wahrscheinlich unfreiwillig hat Pääbo damit einen wichtigen Beitrag zur Stützung des Grundtypenmodells in Bezug auf den Grundtyp Mensch geleistet, welcher moderne Menschen, Neandertaler, Denisova, Heidelberger Menschen, Homo floresiensis und Homo erectus und vielleicht noch andere unentdeckte Menschenformen umfasst.
Anmerkung
1 „… for his discoveries concerning the genomes of extinct hominins and the human evolution”.
Quellen
Borger P (2018) Darwin Revisited – How to understand biology in the 21st century. Scholars Press.
Borger P (2021) Kaum einzigartige DNA-Sequenzen im Erbgut des Menschen. Stud. Integr. J. 28, 134–135.
Frayer DW & Radovčić D (2022) Rockshelter Discoveries Show Neandertals Were a Lot like Us. Scientific American, veröffentlicht am 01.02.2022, https://www.scientificamerican.com/article/rockshelter-discoveries-show-neandertals-were-a-lot-like-us/.
Gibbons A (2016) Neanderthal genes linked to modern diseases. Science 351, 648-849. https://www.science.org/doi/10.1126/science.351.6274.648
Krause J, Lalueza-Fox C, Orlando L, Enard W, Green RE, Burbano HA, Hublin J-J, Hänni C, Fortea J, De la Rasilla M, Bertranpetit J, Rosas A & PääboS (2007) The derived FOXP2 variant of modern humans was shared with Neandertals Curr Biol 17(21):1908-12. https://doi.org/10.1016/j.cub.2007.10.008
Miga KH (2017) Chromosome-Specific Centromere Sequences Provide an Estimate of the Ancestral Chromosome 2 Fusion Event in Hominin Genomes. Journal of Heredity, 108, 45–52; https://doi.org/10.1093/jhered/esw039
Pääbo S (2014) Neanderthal Man. In Search of lost Genomes. 2914 Basic Books, New York. (deutsch: Die Neandertaler und wir. Meine Suche nach den Urzeit-Genen. S. Fischer, Frankfurt a. M.)
Sánchez-Quinto F, Botigue LR, Civit S, Arenas C, Ávila-Arcos MC, Bustamante CD, Comas D & Lalueza-Fox C (2012) North African populations carry the signature of admixture with Neanderthals. PLoS ONE 7: e7765. https://doi.org10.1371/journal.pone.0047765
Scholl B (2020) Frauen mit „Neandertal-Genvarianten“ haben weniger Fehlgeburten. Stud. Integr. J. 27, 126.
Zeberg H & Pääbo S (2020) The major genetic risk factor for severe COVID-19 is inherited from Neanderthals. Nature 587, 610–612. https://www.nature.com/articles/s41586-020-2818-3
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2022, http://www.genesisnet.info/schoepfung_evolution/n306.php
26.08.22 Wie fundiert sind Nasenrekonstruktionen bei Homininen?
Campbell et al. (2022) diskutierten die Rekonstruktion von Nasenweichteilpartien bei Homininen. Dabei stellten sie fest, dass in diesem Forschungsbereich nur wenige Studien vorliegen, obwohl Nasenpartien in vielen Medien in Rekonstruktionen dargestellt werden. Campbell et al. (2022) erstellten daher ein eigenes Rekonstruktionsmodell. Die Gruppe der echten Menschen, die aus Schöpfungsperspektive einen abgrenzbaren Grundtyp darstellt, besitzt in diesen Rekonstruktionen vielfältige, aber durchaus menschlich gestaltete Nasen. Die Nasen-Rekonstruktionen der großaffenähnlichen Australomorphen ist aber auch in diesem Modell mit methodischen Mängeln behaftet, so dass bis heute nicht geklärt ist, wie zum Beispiel die Nase der berühmten „Lucy“ (Australopithecus afarensis) tatsächlich ausgesehen hat.
- Einleitung
Unter den Homininen* wird eine Reihe von Formen aus evolutionärer Perspektive als mutmaßliche Vorfahren des Menschen seit der Abspaltung des Schimpansen interpretiert. Überall in Museen, Filmen oder Büchern werden diese Homininen mit Nasenpartien rekonstruiert. Dabei kann beim unvoreingenommenen Betrachter zum Teil der Eindruck entstehen, dass sie einen Übergang zwischen Großaffen und Menschen darstellen. Doch wie gut sind solche Nasen-Rekonstruktionen eigentlich begründet?
*Homininen (= aus evolutionärer Perspektive Menschen und menschenähnliche Vorfahren, die sich nach der Abspaltung der Schimpansen-Linie entwickelt haben)
In einer neuen Studie aus dem Jahr 2022 haben sich Campbell et al. mit der Rekonstruktion von Nasenweichteilpartien bei Homininen näher beschäftigt. Ihre Ergebnisse werden hier kurz diskutiert.

Abb. 1: Links: Die Rekonstruktionen der Nasenpartie in Museen und Lehrbüchern von nichtmenschlichen Homininen – wie hier von „Lucy“ (Australopithecus afarensis) – sind bisher wissenschaftlich erstaunlich wenig fundiert. Rechts: Die Nasenrekonstruktionen von menschlichen Homininen wie Neandertalern (hier ein Mädchen) sind zwar empirisch besser begründet, aber ohne Weichteilstrukturen trotzdem mit Unsicherheiten behaftet. (Wikimedia: Neanderthal-Museum, Mettmann – Pressebilder, CC BY-SA 4.0; Fährtenleser, CC BY-SA 4.0)
- Der bisherige Stand der Nasenrekonstruktionen
Campbell et al. (2022) sehen bisher einen Mangel bei der Erforschung der Weichteilstrukturen wie der Nase bei Menschenaffen – insbesondere bei der innerartlichen Vielfalt: „Die meisten evolutionären Studien zur Nasenregion haben sich auf moderne Menschen und Neandertaler […] konzentriert. Im Gegensatz dazu wurde dem Weichteilgewebe der Nasen von Menschenaffen nur sehr wenig Aufmerksamkeit geschenkt. Wir erkennen zwar an, dass die Nasen von Schimpansen auf ein gewisses Forschungsinteresse gestoßen sind, aber mit Ausnahme einer Studie gibt es praktisch keine Studien über Gorillas und Orang-Utans“.
Mit anderen Worten: Für heutige Großaffen und großaffenähnliche Australomorphen gibt es zwar viele Darstellungen, aber wenig wissenschaftliche Daten zur Rekonstruktion der Nasenpartie. Die Knochenstrukturen ermöglichen dabei bestenfalls eine grobe Orientierung für die Nasenrekonstruktionen. Für Menschen und Neandertaler liegen hingegen deutlich mehr Untersuchungen vor, so dass die Nasenpartien menschlicher Homininen auf einer breiteren Datenbasis fußen. Allerdings muss erwähnt werden, dass auch bei Neandertalern bisher keine Weichteilstrukturen bekannt sind (vgl. De Azevedo et al. 2017), welche definitive Aussagen ermöglichen.
Auf der Grundlage mehrerer möglichst einfach durchführbarer Messungen an Schädel und Nase bei heutigen Schimpansen und Menschen haben nun Campbell et al. (2022) eigene Gleichungen zur Rekonstruktion der Nase aufgestellt. Diese Rekonstruktionen basieren auf den Messungen der bei Fossilien häufig erhaltenen Winkel zwischen Hinterhauptsloch und Nasenansatz, Nasenspitze oder Oberkiefer an den Schädeln.1
- Die Rekonstruktion nach Campbell und Kollegen
Die Ergebnisse der Rekonstruktionen von Campbell et al. (2022) sind in der folgenden Abbildung dargestellt:

Abb. 2: Nasenrekonstruktionen von Campbell et al. (2022) Nasenrekonstruktionen basierend auf Schädelmessungen von Campbell et al. (2022) bei heutigen Menschenaffen (oben), ausgestorbenen Australomorphen (Mitte) und echten Menschen (unten). Problematisch an diesen Rekonstruktionen ist aber, dass eine evolutionäre Linie unkritisch vorausgesetzt wird. (Eigene Darstellung nach Campbell et al. 2022, Fig. 5, 6)
Es fällt auf, dass bei den nichtmenschlichen Arten „Homo“ habilis und „Homo“ naledi eine Nasenmorphologie rekonstruiert wird, die den Autoren zufolge „weder affenartig noch menschenähnlich ist“. Campbell und Kollegen halten deren Nasenmorphologie aber für „nicht außerhalb des Bereichs des Möglichen“ für Übergangsformen zum Menschen. Die Australopithecinen weisen interessanterweise im Gegensatz zu den echten Menschen weder menschliche noch intermediäre, sondern affenähnliche Nasenstrukturen auf (s. Abb. 2).
Ähnlich schrieben auch der Paleo-Artist (Rekonstrukteur) Viktor Deak und der Paläoanthropologe G. J. Sawyer (2008, 186): „Bei den Australopithecinen lässt der Aufbau des Gesichtsskeletts darauf schließen, dass Lippen und Nase denen der heutigen Menschenaffen ähnelten und in ihrer Form ganz und gar nicht menschlich waren; die kleinen Eckzähne und die übrigen, ebenfalls recht menschenähnlichen Zähne lassen dagegen an ein Gesicht denken, dass nicht vollkommen affenartig war und sich von dem aller heutigen Affen unterschied.“
- Methodische Probleme dieser Rekonstruktion
Doch leider sind auch die Rekonstruktionen von Campbell et al. (2022) – obwohl sie den aktuellen Forschungsstand darstellen – mit einigen methodischen Mängeln behaftet.
Erstens überlappen insgesamt vier von fünf Schädel-Nase-Messwerten von Mensch und Schimpanse, auf welche die Autoren ihre Rekonstruktionen aufbauen, stark (vgl. ebd., Tab. 2). Solche Befunde erschweren die Rekonstruktion der Merkmale.
Zweitens konnten überhaupt nur kleine Stichproben von Menschenaffen untersucht werden – was die Autoren auch ausdrücklich bedauern.
Drittens sind insbesondere beim Zwergschimpansen, Sumatra-Orang-Utan und dem Mantelpavian die mit dem Modell errechneten Vorhersagen für die Nasenhöhlenlänge zunehmend ungenau (ca. 1 cm bis 4 cm Abweichung; ebd., Fig. 4A). Auch die vorhergesagten Winkel der Nase unterscheiden sich von dem tatsächlich gemessenen Winkeln bei Gemeinem Schimpansen, Gorilla und Mantelpavian (1,0° bis 5,5° Abweichung; ebd., Fig. 4B).
Viertens geben Campbell et al. (2022) zu, dass die die Genauigkeit ihrer Vorhersage mit der „phylogenetischen Entfernung dieser Arten zu H. sapiens“ – also deren evolutionären Verwandtschaft zum modernen Menschen – abnimmt. Sollte es sich bei Australomorphen aber um einen oder mehrere nichtmenschliche Grundtypen handeln, die unabhängig von Menschen oder Großaffen erschaffen wurden, ist dementsprechend mit einer deutlich größeren Fehlerquote der Rekonstruktion zu rechnen. Schließlich setzen Campbell et al. (2022) aufgrund des evolutionären Modells – im Unterschied zum Grundtypenmodell – implizit voraus, dass die Australomorphen in ihren Merkmalen zwischen Schimpansen und Menschen stehen, und dass damit Rekonstruktionen weniger fehleranfällig seien.
Und fünftens ist die oben zitierte Aussage über die zunehmende Ungenauigkeit bei phylogenetisch größerer Distanz zu Homo sapiens nicht ganz mit den Ergebnissen der Rekonstruktionen von Campbell et al. (2022) vereinbar. Der Zwergschimpanse steht dem Menschen nämlich aus evolutionärer Perspektive genauso nahe wie der Gemeine Schimpanse, hat aber eine größere Ungenauigkeit in der Vorhersage der Nasenlänge als selbst der Gorilla (ebd., Fig 4A). Der Gorilla wiederum ist in der Vorhersage des Nasenwinkels ungenauer als der Orang-Utan, der seinerseits evolutionär gesehen noch weniger mit dem Menschen verwandt ist (ebd., Fig 4B). Campbell et al. (2022) stellen folgerichtig fest, dass die Ergebnisse der 3D-Modellierung bei Zwergschimpanse sowie Sumatra-Orang-Utan und Siamang (ein großer Gibbon) zu ungenau im Vergleich mit den tatsächlichen Messwerten der Nase sind.
Insgesamt legen die beschriebenen methodischen Mängel nahe, dass nach wie vor keine gut begründeten Nasen-Rekonstruktionen von nichtmenschlichen Australomorphen vorliegen.
- Exkurs: „Neandertaler, wieso hast du eine so große Nase?“
Doch woher kommt es eigentlich, dass ausgestorbene Menschenformen andere Nasenformen aufwiesen – wie zum Beispiel die besonders große Nase der Neandertaler (vgl. Abb. 2)?
Lacruz et al. (2019) diskutieren den Bau des Mittelgesichts bei Homininen. Sie schlagen vor, dass das große vorstehende Mittelgesicht beim Mittelpleistozänen Homo (Homo heidelbergensis, Sima de los Huesos und Neandertaler) im Vergleich zum modernen Menschen vor allem auf eine große Nasenhöhle und oft auch auf große Nasennebenhöhlen zurückzuführen ist. Dies ist neben anderen Faktoren (z. B. Hormonspiegel) wahrscheinlich auch durch eine Anpassung an kaltes Klima bedingt, so dass eine große Nasenpartie die Luft entsprechend erwärmen und befeuchten kann.2 Lacruz und Kollegen (2019) sehen nur bei Homo erectus, Mittelpleistozänem Homo und modernem Menschen ein menschliches Mittelgesicht, nicht aber bei den sogenannten „Homo“-Arten habilis und rudolfensis, die nach Collard & Wood (2015) auch gar keine Menschen (Homo) waren. Lacruz et al. (2019) zufolge ist für den jüngeren „Homo“ naledi nur wenig vom Gesichtsschädel rekonstruierbar, außer dass er „transversal [in Querrichtung] flach, aber insgesamt sehr prognathisch“ war – also eine vorstehende Schnauze besaß.
Die Deutung, dass die großen Nasen der Neandertaler und anderer Menschenformen aus dem Mittelpleistozän Anpassungen an kalte Umgebungen in Europa, West- oder Zentralasien waren (vgl. Fabre et al. 2009, Fig. 2), findet auch bei heutigen Menschen eine Parallele. Bei diesen zeigt sich nämlich nach Zaidi et al. (2017, v. a. Tab. 1) ein deutlicher Zusammenhang von Nasenform und absoluter Feuchtigkeit sowie Temperatur des Lebensraumes. So haben Nord-Europäer in kälteren Klimaten zur Erwärmung der Luft die längsten und schmalsten Nasen (durchschnittlich 4,78 cm lang und 3,43 cm breit). West-Afrikaner in heißeren Klimaten besitzen hingegen die kürzesten und breitesten Nasen (durchschnittlich 4,63 cm lang und 4,54 cm breit).
- Fazit
Die evolutionären Rekonstruktionen der Nasenpartien von nichtmenschlichen Homininen wie „Lucy“, die immer wieder in Museen, Medien und sogar Lehrbüchern gezeigt werden, sind äußerst spekulativ. Ursache hierfür sind die mangelnden empirischen Daten über heute lebende Menschenaffen und fossile Homininen sowie methodische Probleme bei Rekonstruktionsmodellen. Dies gilt leider auch für die neue Studie von Campbell et al. (2022), bei der die Australomorphen – passend zum Grundtypmodell – keine menschlichen Nasenpartien aufweisen.
Anmerkungen:
1 Winkelmessungen: Winkel Nr. 1: Basion – Nasion zu Basion – Pronasale. Winkel Nr. 2: Basion – Pronasale zu Basion – Prosthion. Basion: Der hinterste Punkt des Hinterhauptlochs (midsaggitale Ebene). Nasion: Schnittpunkt der nasofrontalen Naht in der Medianebene. Pronasale: Der vorderste Punkt der Nasenspitze. Prosthion: Der vorderste Punkt des Oberkiefers (midsaggitale Ebene).
2 Lacruz et al. (2019) stellen die Frage, ob auch eine zunehmende Sozialfunktion der Grund für eine „komplexere Topografie“ des Mittelgesichts beim modernen Menschen im Vergleich zum Mittelpleistozänen Menschen gewesen sein könnte. Diese Hypothese basiert aber nur auf evolutionären Annahmen und bleibt ohne vorliegende Weichteilstrukturen pure Spekulation. Das biblisch motivierte Grundtypmodell hingegen setzt voraus, dass der Mensch von Anfang an als Gottes Ebenbild und intelligentes, soziales Wesen erschaffen wurde, bevor er sich über die Welt verbreitete und sich im Rahmen von Mikroevolution an die Umwelt anpasste.
Literatur
Campbell RM, Vinas G & Henneberg M (2022) Relationships between the hard and soft dimensions of the nose in Pan troglodytes and Homo sapiens reveal the positions of the nasal tips of Plio-Pleistocene hominids. PLoS ONE 17, e0259329; https://doi.org/10.1371/journal.pone.0259329.
Collard M & Wood B (2015) Defining the Genus Homo. In: Henke W & Tattersall I (Ed.) Handbook of Paleoanthropology, doi: 10.1007/978-3-642-39979-4_51.
De Azevedo S et al. (2017) Nasal airflow simulations suggest convergent adaptation in Neanderthals and modern humans. PNAS 114, 12442–12447, doi: 10.1073/pnas.1703790114.
Fabre V & Condemi S & Degioanni A (2009) Genetic Evidence of Geographical Groups Among Neanderthals. PLoS ONE 4, e5151, doi: 10.1371/journal.pone.0005151.
Lacruz RS et al. (2019) The evolutionary history of the human face. Nat. Ecol. Evol. 3, 726–736, https://doi.org/10.1038/s41559-019-0865-7.
Sawyer GJ & Deak V (2008) Der lange Weg zum Menschen. Lebensbilder aus 7 Millionen Jahren Evolution. Spektrum Akademischer Verlag.
Zaidi AA et al. (2017) Investigating the case of human nose shape and climate adaptation, PLoS Genet. 13, e1006616, https://doi.org/10.1371/journal.pgen.1006616.
Aktualisierung am 23.01.2023.
Autor dieser News: Benjamin Scholl
© 2022, http://www.genesisnet.info/schoepfung_evolution/n305.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.07.22 Sind die Schädel von Dmanisi Übergangsformen zum echten Menschen?
In Form eines Überblicks werden die wichtigsten Argumente zu Bewertung der Homininen-Funde aus Dmanisi (Georgien) zusammengestellt. Er zeigt, dass man diese Funde als echte Menschen statt als evolutionäre Übergangsformen deuten kann. Eine ausführliche Begründung wird in einem Special Paper mit dem Titel „Homininen-Schädel: ‚Stolpersteine‘ des Grundtypmodells?“ geboten: http://www.wort-und-wissen.de/artikel/homininen-schaedel/
- Die Schädel von Dmanisi als Übergangsformen?
Schädel früher Homininen1 werden immer wieder als Belege einer mutmaßlichen Abstammung des Menschen aus affenähnlichen Vorfahren angeführt. Zu den Schädeln, die häufig als Übergangsformen gewertet werden, gehören die Schädel von den sogenannten „Homo“-Arten habilis und rudolfensis aus Afrika und die Schädel von Homo erectus ergaster georgicus aus Dmanisi in Georgien. Letztere datiert man auf ca. 1,8 MrJ (Millionen radiometrische Jahre). Über sie heißt es in der deutschsprachigen Wikipedia: „Die homininen Fossilien von Dmanisi gelten als mögliches Bindeglied zwischen den frühesten Vertretern der Gattung Homo aus Afrika und den späteren, aus Asien bekannten Fossilien des Homo erectus“.2
Lordkipanidze und Kollegen (2013) behaupteten gar, dass die großaffenartigen „Homo“-Arten habilis und rudolfensis mit den Homo–erectus-Funden aus Dmanisi sowie aus Afrika und Asien in die gleiche Art gestellt werden sollten, weil sich die Schädel ausreichend ähneln würden.
Dies widerspricht dem Grundtypenmodell nach Brandt (2017), bei dem „Homo“ habilis und rudolfensis als großaffenähnliche Australomorphen und alle Formen von Homo erectus als echte Menschen betrachtet werden. Eine Studie zum mathematischen Schädelvergleich der Homininen von Lauren Schroeder et al. (2017) sowie andere Erkenntnisse aus der Fachliteratur über die Funde aus Dmanisi liefern jedoch unterstützende Hinweise für die Deutung als getrennt erschaffene Grundtypen (vgl. den ausführlichen Fachartikel von Scholl 2022b).
- Die Schädelvergleiche von Schroeder et al. (2017) unterstützen das Grundtypenmodell prinzipiell
Schroeder et al. (2017) verglichen in ihrer Studie anhand von 19 Messpunkten fünf verschiedene Flächen auf den Schädeln von Dmanisi und von über 30 anderen Homininen.
Insgesamt gibt es bei den Daten ihrer Studie eine große Übereinstimmung mit den Ähnlichkeitsbeziehungen, die man aufgrund des Grundtypmodells erwarten würde. Elf der dreizehn untersuchten Homininen-Spezies haben die größte Ähnlichkeit mit solchen Arten, die zum selben mutmaßlichen Grundtyp gehören. Besonders hervorzuheben ist, dass alle Australomorphen – inklusive der sogenannten „Homo“-Arten rudolfensis, habilis und naledi – ihre jeweils ähnlichsten Schädelformen innerhalb der Australomorphen besitzen, welche einen oder mehrere Grundtypen von Großaffen darstellen. Nicht nur das Körperskelett (vgl. Brandt 2017, 51–92; Collard & Wood 2015), sondern auch die Schädelmerkmale legen damit einen Ausschluss dieser drei Arten aus der Gattung Homo (Mensch) nahe. Die Schädel von Homo erectus aus Dmanisi sind dagegen von denen der Australomorphen verschieden und ähneln anderen echten Menschen (wie Heidelberger Mensch und Neandertaler) am stärksten. Sie eignen sich daher nicht gut als Übergangsform von Australomorphen hin zu Menschen.
Abbildung: Die Dmanisi-Schädel: Die Dmanisi Schädel 1 bis 5 hatten zwar ein kleines Gehirnvolumen von 546 bis 775 cm³, sind aber doch der Menschenart Homo erectus zuzuordnen. (Nach Wikimedia: Gerbil, CC BY-SA 4.0; Emőke Dénes, CC BY-SA 4.0)
Doch was ist mit den beiden Ausnahmen, bei denen die größten Ähnlichkeiten der Schädelflächen zu Arten aus einem anderen Grundtyp bestehen?
Die erste Ausnahme ist der Mensch Homo floresiensis, der zu keiner Art eine größere Ähnlichkeit aufweist, aber entfernt dem großaffenartigen Australopithecus sediba (vgl. Scholl 2022a) in den Schädelflächen ähnelt. Allerdings könnte man Homo floresiensis aufgrund seines extremen Zwergwuchses als einzigartigen Menschen interpretieren, der wegen dieser Einzigartigkeit zu keinem der untersuchten Homininen eine besonders große Ähnlichkeit der Schädelflächen aufweist. Eventuell spielt bei einer groben Ähnlichkeit der Schädelflächen die vergleichbare Gehirngröße von jeweils ca. 420 cm³ von Australopithecus sediba und Homo floresiensis eine Rolle (vgl. Berger et al. 2010; Kubo et al. 2013; Lacruz et al. 2019); die aber nichts mit einer gemeinsamen Abstammung zu tun haben muss.
Die zweite Ausnahme ist Homo erectus aus Afrika und Asien. Dieser weist zu „Homo“ habilis eine minimal größere Nähe (3,7 Prozentpunkte) auf als zu Homo erectus aus Georgien. Die Nähe zu „Homo“ habilis sollte aber nicht überbetont werden, weil von „Homo“ habilis nur in zwei von fünf Flächenvergleichen fossiles Vergleichsmaterial vorliegt. Außerdem entsteht die größere Nähe von Homo erectus aus Afrika und Asien zu „Homo“ habilis als zu Homo erectus aus Dmanisi im Wesentlichen durch einen einzigen Unterkiefer (SK 15), dessen Zuordnung zu „Homo“ habilis möglicherweise falsch ist. Dem Paläoanthropologen John Hawks (2016) zufolge ist die Zuordnung des Unterkiefers SK 15 unklar, und mittlerweile stellt „eine Reihe Wissenschaftler“ SK 15 zu „Homo“ habilis (Hawks 2016; vgl. Curnoe 2006: größte Ähnlichkeit bei Messwerten zu „Homo“ habilis ), oder betont gar eine Ähnlichkeit mit den Australopithecinen (vgl. die Zahnanalyse von Zanolli et al. 2022).
Insgesamt lassen sich also auch die Ähnlichkeitsbeziehungen der Schädelflächen von Homo floresiensis und Homo erectus aus der Grundtypenperspektive befriedigend interpretieren. Außerdem muss noch erwähnt werden, dass sich die enorme innerartliche Variabilität (Vielfalt) der Homininen-Schädel ganz allgemein einer widerspruchsfreien Stammbaumrekonstruktion bei Schroeder et al. (2017) widersetzt. Dies wird auch dadurch deutlich, dass die einzelnen fossilen Schädel in drei Viertel der Flächenvergleiche Schädeln aus anderen Arten mehr ähneln als den Schädeln ihrer eigenen Artgenossen. Das wiederum zeigt, dass Schädel sehr variabel sind und daher für Fragestellungen der Zuordnung (Taxonomie) und der Abstammung nur begrenzte Informationen liefern können.
- Weitere Argumente für die Menschlichkeit von Homo erectus aus Dmanisi
Trotz der hohen Variabilität der Homininen-Schädel hatten Lordkipanidze et al. (2013) ihre These, dass die großaffenähnlichen „Homo“-Arten habilis und rudolfensis gemeinsam mit Menschen Homo erectus aus Afrika, Asien und Georgien eine Art bilden sollen, ausschließlich auf Schädelmerkmale gegründet. Dieses Vorgehen wurde dementsprechend von Wood (2014) sowie von Collard & Wood (2015, 2130) scharf zurückgewiesen. Sie kritisieren explizit an der Vorgehensweise von Lordkipanidze und Kollegen, dass diese ausschließlich Schädelmerkmale verwendeten und viele weitere Unterscheidungsmerkmale ignoriert haben. Dazu zählt eine Detailanalyse des Basicraniums (des unteren Schädels), des knöchernen Innenohrs, des Fußes und der Hand, der Form und Stärke der Langknochen, der relativen Zahngröße und auch der Individualentwicklung. Zusammengefasst bewerten Collard & Wood (2015) die Gründe von Lordkipanidze et al. (2013) für die Zusammenlegung von „Homo“ habilis und Homo erectus zu einer einzigen gemeinsamen Art als nicht gerechtfertigt und wenig tragfähig. Insgesamt hält Wood (2014) „Homo“ habilis nicht einmal für ähnlich genug zu Homo erectus, um als dessen direkter Vorfahre betrachtet werden zu können.
Es gibt zwar eine ganze Reihe von Detailmerkmalen, bei welchen die vielfältigen Formen des frühen Menschen Homo erectus gewisse Ähnlichkeiten mit der Vielfalt ausgestorbener Großaffen als auch mit „Homo“ habilis aufweisen. Dennoch sprechen ganz wesentliche Argumente gegen eine gemeinsame Abstammung bzw. Übergangsstellung von „Homo“ habilis und rudolfensis zu Homo erectus.
Eines dieser Argumente betrifft die Schädel aus Dmanisi: So ist die Ähnlichkeit der fünf Schädel aus Dmanisi mit Homo erectus aus Asien und Afrika ziemlich eindeutig (vgl. Gabunia et al. 2000; Lordkipanidze et al. 2006; Hawks 2013; und z.T. auch Lordkipanidze et al. 2013). Homo erectus aus Afrika lässt sich wiederum insbesondere am Gesichtsschädel gut von „Homo“ habilis und rudolfensis unterscheiden (Lacruz et al. 2019). Die Vielfältigkeit der Schädel aus Dmanisi übersteigt die der Schädel von „Homo“ habilis und rudolfensis – entgegen der Behauptung von Lordkipanidze et al. (2013) – gar nicht, und macht es daher auch nicht notwendig, diese beiden Gruppen einer gemeinsamen Sammel-Art zuzuordnen (vgl. Hawks 2013; Lordkipanidze et al. 2013, Fig. S7).
Der Schädel Nr. 5 aus Dmanisi ist zwar äußerst klein (546 cm³) und besitzt eine deutliche Prognathie (vorstehender Oberkiefer); diese beiden Merkmale sind aber bei heutigen bzw. fossilen Menschen nicht so außergewöhnlich. Prognathie tritt auch bei heutigen Menschen mit dem Möbius-Syndrom auf (vgl. Pedersen et al. 2017). Eine kleine Gehirngröße findet sich ähnlich auch bei Homo floresiensis sowie bei heutigen Menschen mit Mikroencephalie (vgl. Falk et al. 2007), während das Gehirn bei manchen Mitgliedern heutigen Naturvölkern und anderen Homo–erectus-Funden nur wenig größer ist (vgl. Wells 1942; Brandt 2000, 13–15).
Insbesondere der mit 598 cm³ Gehirnvolumen ebenfalls winzigen Schädel DAN5 aus Afrika weist deutliche Parallelen auf, die entscheidende Hinweise für die Deutung des Schädels 5 aus Dmanisi liefern (vgl. Semaw et al. 2020). Beide Schädel sind sehr klein und weisen doch genügend Ähnlichkeiten mit anderen Homo–erectus-Funden auf, so dass beide der Art Homo erectus zugeordnet werden (vgl. Semaw et al. 2020).
Bei dem Schädel DAN5 wurden zudem sowohl einfache Geröllwerkzeuge (Oldowan, Mode I) als auch komplexe Faustkeile (Acheuléen, Mode II) gefunden. Dies bedeutet, dass einfachere Werkzeuge oder ein kleines Gehirn nicht unbedingt für eine geringe Intelligenz sprechen müssen, da komplexere Werkzeuge ebenfalls von Menschen mit einem so kleinen Gehirn wie bei DAN5 gefertigt werden können. Die Abwesenheit von komplexeren Werkzeugen könnte daher auch mit den jeweiligen Lebensumständen statt mit mangelnder Intelligenz erklärt werden (vgl. Semaw et al. 2020). Des Weiteren ist eine kleine Gehirngröße (auch relativ zur Körpergröße) allgemein kein guter Indikator für Intelligenz bei Homininen; und sie kann außerdem auch durch ungünstige Lebensbedingungen verursacht sein (vgl. Brandt 2000, 22–25; Roth 2010, 340f; Antón & Snodgrass 2012; Brandt 2018).
Dazu kommt, dass die Steinwerkzeugfunde aus Dmanisi gar nicht so „einfach“ sind, wie ursprünglich vermutet und publiziert worden war (vgl. Hartwig-Scherer 2000; 2002). Dies zeigen die Untersuchungen von Baena et al. (2010) an den Steinwerkzeugen aus Dmanisi, die auf 1,77 bis 1,85 MrJ datiert werden (vgl. Ferring et al. 2011): Die Menschen aus Dmanisi verwendeten 24 verschiedene Rohmaterialien, schätzten das Rohmaterial hinsichtlich Arbeitseffizienz ein, rotierten die Steine je nach Zweck, nutzen natürliche Vorteile des Ausgangsgesteins, besaßen verschiedene Fertigungstechniken, bearbeiteten die Steine nach (Retusche) und fertigten sogar eine Klinge, die der Levallois-Technik des Neandertalers ähnelte. Dies alles weist auf eine „bedeutende Erfahrung“ bei der Steinwerkzeugfertigung (Baena et al. 2010) hin – und somit auf ausgeprägte Kultur und Intelligenz bei den Menschen aus Dmanisi.
Auch die lange Überlebenszeit des zahnlosen Menschen Nr. 4 aus Dmanisi spricht wahrscheinlich für eine typisch menschliche Fürsorge für Gruppenmitglieder (vgl. Hartwig-Scherer 2006; Lordkipanidze et al. 2006; Roberts 2018, 111; Scholl 2018).
Damit sind die Schädel aus Dmanisi zwar deutlich kleiner als die allermeisten heutigen Menschen, ihre Besitzer können aber trotzdem eine normal menschliche Intelligenz besessen haben, da selbst moderne Naturvölker zum Teil nur wenig komplexe Steinwerkzeuge verwenden (vgl. Brandt 2000, 13–15; Brandt 2019, 398).
Das Körperskelett von Homo erectus aus Dmanisi ist insgesamt deutlich menschlich konfiguriert. Somit besaßen die Homo–erectus-Formen einen gewohnheitsmäßigen (habituellen) menschlichen Gang (vgl. Lordkipanidze et al. 2007; Lordkipanidze et al. 2013; Domínguez-Rodrigo et al. 2015; Collard & Wood 2015, 2110; Brandt 2017, 93–108). So schreiben Tappen et al. (2022): “Wichtig ist, dass H. erectus in Dmanisi als obligater terrestrischer Zweibeiner anerkannt ist“. Außerdem waren sie vor ca. 2 MrJ über längere Zeit Zeitgenossen von „Homo“ habilis und rudolfensis (vgl. Wood & Boyle 2016), obwohl letztere häufig als deren Nachfahre vorgeschlagen wurden.
- „Homo“ habilis war kein Mensch
Abbildung: Die Schädel von „Homo“ habilis: Die nachfolgend genannten Schädel werden „Homo“ habilis zugeschrieben, sind aber bis auf OH 7 (dem Holotyp der Spezies) letztlich alle umstritten: OH 7 (kaum erhalten), OH 24, KNM-ER 1805, KNM-ER 1813, SK 847 und STW 53. (Wikimedia: Nachosan, CC BY-SA 3.0; Guérin Nicolas, CC BY-SA 3.0; Ryan Somma, CC BY-SA 2.0; Tiia Monto, CC BY-SA 3.0; Rhuangagabi, CC BY-SA 4.0)
Die oft „Homo“ habilis und rudolfensis zugeordneten Fossilienweisen im Gegensatz zu Homo erectus aus Dmanisi viele großaffenähnliche Merkmale am Schädel und Körperskelett auf (vgl. Clarke 2008; Wood 1996; Wood 2014; Collard & Wood 2015; Grabowski et al. 2015; Brandt 2017, 86–88; Lacruz et al. 2019; Prahabat et al. 2021). Darüber hinaus ist die Rekonstruktion ihrer größeren Hirnschädelvolumina (über 700 cm³) umstritten – ebenso wie die Frage, welche Skelettfragmente man überhaupt diesen beiden Homininenarten zuordnen soll (vgl. Scholl 2022, Anhang Tab. 14).
- Fazit
Alles in allem kann man daher Homo erectus aus Dmanisi mit guten Argumenten hinsichtlich seines Schädelbaus, seines Körperskeletts und seiner Steinwerkzeuge aus Grundtyp-Perspektive als echten Menschen deuten. Die Parallelen zu dem kleinen Schädels DAN5 aus Afrika machen deutlich, dass kleine Schädel bei Homo erectus nichts Ungewöhnliches waren, und dass auch solche Menschen hohe kognitive Fähigkeiten bei der Herstellung von Steinwerkzeugen bewiesen haben. Entsprechend eindeutige Hinweise konnten in Bezug auf vergleichbar anspruchsvolle Fähigkeiten der Werkzeugherstellung bei „Homo“ habilis und rudolfensis bisher nicht erbracht werden (vgl. Roberts 2018, 102f). Die nicht wesentlich älteren „Homo“-Arten habilis und rudolfensis lassen sich hingegen aus der Perspektive des Grundtypmodells hinsichtlich Schädelbau und Körperskelett gut den – getrennt vom Menschen erschaffenen – australomorphen Großaffen zuordnen.
Anmerkungen:
1 Homininen sind vermutete Vorfahren des Menschen im Rahmen des Evolutionsmodells seit Aufspaltung von den Schimpansenvorfahren vor ca. 5 bis 7 MrJ.
2 Wikipedia.de, aufgerufen am 25.02.2022, https://de.wikipedia.org/w/index.php?title=Hominine_Fossilien_von_Dmanissi&oldid=218433192.
Literatur:
Antón SC & Snodgrass JJ (2012) Origins and Evolution of Genus Homo: New Perspectives. Curr. Anthropol. 53, 479–496, doi: 10.1086/667692.
Baena J et al. (2010) Technical and technological complexity in the beginning: The study of Dmanisi lithic assemblage. Quat. Int. 223–224, 45–53, https://doi.org/10.1016/j.quaint.2010.01.019.
Berger LR et al. (2010) Australopithecus sediba: A New Species of Homo-Like Australopith from South Africa. Science 328, 195–204.
Brandt M (1995) Der Ursprung des aufrechten Ganges. Zur Fortbewegung plio-pleistozäner Hominiden. Studium Integrale Paläontologie. Hänssler-Verlag.
Brandt M (2000) Gehirn, Sprache, Artefakt. Fossile und archäologische Zeugnisse zum Ursprung des Menschen. Studium Integrale Paläontologie. Hänssler-Verlag.
Brandt M (2017) Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special. SCM Hänssler.
Brandt M (2018) Merkmalsnetz statt Stammbaum. Neues Vernetzungsmodell in der Paläanthropologie ähnelt Verwandtschaftsverhältnissen im Grundtyp Mensch. Stud. Integr. J. 25, 47–51.
Brandt M (2019) Vergessene Archäologie. Steinwerkzeuge fast so alt wie die Dinosaurier. 2. erw. Aufl. SCM Hänssler.
Clarke R (2008) Latest information on Sterkfontein’s Australopithecus skeleton and a new look at Australopithecus. S. Afr. J. Sci. 104, 443–449, http://www.scielo.org.za/pdf/sajs/v104n11-12/a1510412.pdf.
Collard M & Wood B (2015) Defining the Genus Homo. In: Henke W, Hardt T & Tattersall I (Ed.) Handbook of Paleoanthropology. Springer‐Verlag, 2107–2144, doi: 10.1007/978-3-642-39979-4_51.
Curnoe D (2006) Odontometric systematic assessment of the Swartkrans SK 15 mandible: Homo 57, 263–294, doi: 10.1016/j.jchb.2006.02.001.
Domínguez-Rodrigo M et al. (2015) Earliest modern human-like hand bone from a new 1.84-million-year-old site at Olduvai in Tanzania. Nat. Commun. 6, 7987, doi: 10.1038/ncomms8987.
Falk D et al. (2007) Brain shape in human microcephalics and Homo floresiensis. PNAS 104, 2513–25188, doi: 10.1073/pnas.0609185104.
Ferring R et al. (2011) Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85–1.78 Ma. PNAS 108, 10432–10436, https://doi.org/10.1073/pnas.1106638108.
Gabunia L (2000) Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: taxonomy, geological setting, and age. Science 288, 1019–1025, doi: 10.1126/science.288.5468.1019.
Grabowski M, Hatala KG, Jungers WL & Richmond BG (2015), Body mass estimates of hominin fossils and the evolution of human body size. J. Hum. Evol. 85, 75–93, http://dx.doi.org/10.1016/j.jhevol.2015.05.005.
Hartwig-Scherer S (2000) Der erste Europäer? Streiflichter. Stud. Integr. J. 7, 92–100.
Hartwig-Scherer S (2002) Wurde Europa doch früher besiedelt? Überraschungen aus Ost und West. Stud. Integr. J. 9, 67–73.
Hartwig-Scherer S (2006) Mensch von Anfang an: Altruismus so alt wie die Gattung Homo. Stud. Integr. J. 13, 52, https://www.si-journal.de/jg13/heft1/sij131-s.html#idx13.
Hawks J (2013) The new skull from Dmanisi. john hawks weblog, veröffentlicht am 18.10.2013, https://johnhawks.net/weblog/fossils/lower/dmanisi/d4500-lordkipanidze-2013.html.
Hawks J (2016) Photo: SK 15. john hawks weblog, veröffentlicht am 05.08.2016, https://johnhawks.net/weblog/topics/photos/sk-15-photo-2016.html.
Kubo D, Kono RT & Kaifu Y (2013) Brain size of Homo floresiensis and its evolutionary implications, Proc. R. Soc. B 280, 20130338, https://doi.org/10.1098/rspb.2013.0338.
Lacruz RS et al. (2019) The evolutionary history of the human face. Nat. Ecol. Evol. 3, 726–736, https://doi.org/10.1038/s41559-019-0865-7.
Lordkipanidze D et al. (2006) A fourth hominin skull from Dmanisi, Georgia. Anat. Rec. 288A, 1146–1157, https://doi.org/10.1002/ar.a.20379.
Lordkipanidze D et al. (2007) Postcranial Evidence from Early Homo from Dmanisi, Georgia. Nature 449, 305–310, doi: 10.1038/nature06134.
Lordkipanidze D et al. (2013) A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo. Science 342, 326–31, doi:10.1126/science.1238484.
Pedersen LK et al. (2017) Moebius sequence – a multidisciplinary clinical approach. Orphanet. J. Rare. Dis. 12, 4, https://doi.org/10.1186/s13023-016-0559-z.
Prabhat AM et al. (2021) Homoplasy in the evolution of modern human-like joint proportions in Australopithecus afarensis. eLife 10, e65897, https://doi.org/10.7554/eLife.65897.
Roberts A (2018) Die Anfänge der Menschheit. Vom aufrechten Gang bis zu den frühen Hochkulturen. Dorling Kindersley Verlag.
Roth G (2010) Wie einzigartig ist der Mensch? Die lange Evolution der Gehirne und des Geistes. Springer-Verlag.
Scholl B (2018) Affe = Mensch? Ein Überblick über verhaltensbiologische Unterschiede zwischen Affen und Menschen. W+W Special Paper B-18-1, https://www.wort-und-wissen.org/artikel/verhalten-affe-mensch/.
Scholl B (2022a) Paläontologie: Woher kommt der Mensch? factum 4, 42–43.
Scholl B (2022b) Homininen-Schädel: „Stolpersteine“ des Grundtypmodells? Eine schöpfungstheoretische Deutung der Funde von Dmanisi. W+W Special Paper, https://www.wort-und-wissen.org/artikel/homininen-schaedel/.
Semaw S et al. (2020) Co-occurrence of Acheulian and Oldowan artifacts with Homo erectus cranial fossils from Gona, Afar, Ethiopia. Sci. Adv. 6, eaaw4694, doi: 10.1126/sciadv.aaw4694.
Tappen M et al. (2022) Life and death at Dmanisi, Georgia: Taphonomic signals from the fossil mammals. J. Hum. Evol. 171, 103249, https://doi.org/10.1016/j.jhevol.2022.103249.
Wells RJ (1942) A Diminiutive Skull from Peru. Am. J. Phys. Anthropol. 29, 425–427.
Wood B & Boyle EK (2016) Hominin Taxic Diversity: Fact or Fantasy? Am. J. Phys. Anthropol. 159, 37–78, doi: 10.1002/ajpa.22902.
Wood B (1996) Homo habilis und Homo rudolfensis. In: Aiello L & Bennike P (1996) 4 Millionen Jahre Mensch. Das einzigartige Schauspiel der Menschheitsgeschichte. United Exhibits Group.
Wood B (2014) Human evolution: Fifty years after Homo habilis. Nature 508, 31–33, https://doi.org/10.1038/508031a.
Zanolli C, Davies T, Joannes-Boyau R & Beaudet A (2022) Dental data challenge the ubiquitous presence of Homo in the Cradle of Humankind. PNAS 119, e2111212119, doi: 10.1073/pnas.2111212119.
Aktualisiert am 02.02.2023
Autor dieser News: Benjamin Scholl
© 2022, http://www.genesisnet.info/schoepfung_evolution/n303.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.02.22 Eine modern menschliche Wirbelsäule bei Australopithecus sediba?
Auf der Basis neu gefundener Lendenwirbel des Homininen Australopithecus sediba schließen Forscher um Lee Berger, dass dieser mutmaßliche Vorfahr des Menschen menschlich aufrecht gehen konnte. Eine detaillierte Analyse der zugrundeliegenden Befunde kann diese Einschätzung jedoch keineswegs bestätigen. Es zeigt sich, dass die ermittelten Daten an den fossilen Lendenwirbeln und die paläoanthropologischen Deutungen sowie die populäre Darstellung in den Medien nicht zusammenpassen. Dies liegt daran, dass die untere Wirbelsäule von Australopithecus sediba trotz krasser Krümmung insgesamt hinsichtlich Winkel der Wirbel, Gelenkflächen und Wirbelform nicht menschenähnlicher konfiguriert ist als bei Australopithecus africanus, sondern eindeutig australopithecusartig. Somit liegen keine belastbaren Hinweise auf eine Menschenähnlichkeit der unteren Wirbelsäule von Australopithecus sediba vor.
- Einleitung
„,Er lief wie ein Mensch und kletterte wie ein Affe‘, [so] fasst Lee Berger, Paläontologe […] in Südafrika, die Ergebnisse einer neuen Studie über Australopithecus sediba zusammen“ (Dönges 2021). Diese neue Studie von Williams et al. (2021) handelte über neu entdeckte Lendenwirbel L1 bis L4 von Australopithecus sediba.
Es sind insgesamt ca. 10 fossile Arten von Australopithecus bekannt, von denen manche robuster und manche graziler gebaut waren. Nach evolutionärer Deutung der Herkunft des Menschen wird Australopithecus zu den „Homininen“ gerechnet. Dieser Begriff fasst alle fossilen Formen zusammen, die man nach der vermuteten Abspaltung der Vorfahren des Schimpansen von den Vorfahren des Menschen vor 5 bis 7 Millionen radiometrischen Jahren (MrJ) einordnet. Verschiedene Australopithecus-Arten werden insbesondere aufgrund von potenziellen Skelettanpassungen an einen aufrechten Gang als direkte Vorfahren des Menschen diskutiert. Dazu gehört auch der 2 MrJ alte Australopithecus sediba aus Südafrika, der erst ab 2010 wissenschaftlich beschrieben und als Übergangsform zwischen älteren Australopithecus-Arten und der Gattung Homo (Mensch) vorgeschlagen wurde. Im Folgenden wird untersucht, ob das Eingangszitat von Lee Berger durch die vorgestellten Daten unterstützt wird.

Abb. 1: Links: Die beiden Skelette MH1 und MH2 von Australopithecus sediba. Rechts: Die neu entdeckten Lendenwirbel von MH2 (im roten Kasten), wobei nur L2 bis L3 relativ vollständig sind. (eigene Darstellung nach: Wikimedia, Profberger, CC BY-SA 3.0, sowie Lendenwirbel nach: Williams et al. 2021, Figure 3, elife, CC BY 4.0)
- Die Lendenwirbelsäule von Australopithecus sediba
Die Wirbelsäule beim Menschen ist typisch doppelt-S-förmig gekrümmt, um die Gewichtsbelastung und Kraftübertragung beim aufrechten Gang zu kompensieren, während sie bei heutigen Menschenaffen nur einfach S-förmig gekrümmt ist. Williams et al. (2021) wollen anhand der Winkel der fossilen Lendenwirbel zueinander auf die Art der Wirbelsäulenkrümmung schließen. Die Winkel der Wirbel L2 bis L4* haben beim modernen Menschen einen riesigen Variationsbereich, der alle fossilen Homininen sowie die allermeisten heutigen Menschenaffen (Schimpanse, Gorilla und Orang-Utan) einschließt. Damit ermöglicht dieses Merkmal keine sicheren Aussagen über Abstammung oder Fortbewegung.
*Bzw. der entsprechenden Wirbel L1 bis L3 bei heutigen Menschenaffen, die oft einen Lendenwirbel weniger haben.
Williams et al. (2021) bezeichnen den Übergang vom positiven L3-Winkel zum negativen L4-Winkel bei Australopithecus sediba als „menschenähnliche“ Wirbelsäulenkrümmung und Anpassung an Zweibeinigkeit. Diese Schlussfolgerung ist aber methodisch nicht möglich, weil ca. 18 Prozent von Schimpansen, Gorillas und Orang-Utans ebenfalls einen solchen Übergang von L3 zu L4 besitzen, der aber 60 Prozent der heutigen Menschen fehlt (ebd. Tab. 2+3 sowie Fig. 5–source data 1).
Der Winkel des Wirbels L5 hat einen sehr großen Variationsbereich beim Menschen (-12,3° bis 2,2°), der beinahe komplett den Variationsbereich des vorzugsweise am Boden lebenden Gorillas (-6,2° bis 3,5°) abdeckt. Schimpansen und Orang-Utans haben aber zum Teil positivere Winkelwerte (bis 7 bzw. 8°). Die drei Winkelwerte von L5 beim Neandertaler (4,9° bis -10,6°) bewegen sich in deutlich menschenähnliche Richtung, was auch für Australopithecus africanus gilt (-6,9° bis -4,2°). Der sehr stark negative L5-Winkel von A. sediba (-11,2°) ist im Bereich des unteren Endes des Variationsbereichs des modernen Menschen und nahe Homo erectus (-11,8°) gelegen. Andererseits besitzen zwei untersuchte nichtpathologische moderne Menschen sogar positive und drei andere Menschen fast neutrale L5-Winkelwerte. Außerdem haben ca. 54 Prozent aller heutigen Menschenaffen einen negativen L5-Winkelwert und vier untersuchte Menschenaffen liegen sogar näher am Mittelwert des Menschen als A. sediba. Aus einem negativen L5-Winkel kann somit nicht direkt auf einen aufrechten Gang geschlossen werden.
Auch die Aufsummierung der Winkelwerte der betreffenden Wirbel ergibt kein klares Signal in Richtung menschenartigere Lendenwirbelsäule von Australopithecus sediba (Williams et al. 2021, Fig. 5 + source data 1, Tab. 2+3), weil sowohl heutige als auch fossile echte Menschen (Homo erectus und Neandertaler) jeweils einen riesigen Variationsbereich abdecken. Insgesamt liegen bei den Gesamtwinkeln ca. 74 Prozent aller Schimpansen, Orang-Utans und Gorillas im Variationsbereich des modernen Menschen, von denen ein untersuchtes Gorilla-Exemplar sogar einen noch negativeren Wert als Australopithecus sediba hat. Sieht man einmal vom Mangel an Vergleichsmaterial von anderen Affen ab (vgl. Almécija et al. 2021), liegen selbst zwei Fossilien von Australopithecus africanus (-3,8° bzw. 1,0°) näher an dem Mittelwert heutiger Menschen (-1,6°) als A. sediba (-7,3°), weswegen letzterer nicht „menschenähnlicher“ sein kann.
Williams et al. (2021) gehen davon aus, dass beim Menschen die Gelenkflächen zwischen den Wirbeln der Lendenwirbelsäule (bzw. des Kreuzbeins) nach unten hin „pyramidenförmig“ (L2 zu L3 im Verhältnis zu L5 zu S1) größer werden, um als Anpassungen an die Kräftewirkungen beim aufrechten Gang zu fungieren. Die Gelenkflächenverhältnisse von Menschen und heutigen Menschenaffen überlappen tatsächlich ganz knapp nicht. Dagegen fallen alle Homininen-Fossilien außer Lucy (Australopithecus afarensis, mit unklarer Lendenwirbelzuordnung) eindeutig in den Variationsbereich des Menschen. Williams et al. (2021) leiten aus den Messdaten „eine menschenähnliche Konfiguration des Neuralbogens“ bei Australopithecus sediba ab. Es muss aber beachtet werden, dass Sts 14 von Australopithecus africanus hinsichtlich der Gelenkflächenverhältnisse (1,54) sogar noch näher am Mittelwert moderner Menschen (1,68) als MH2 von A. sediba (1,43) liegt. A. sediba befindet genau zwischen den beiden Werten von A. africanus und ist somit typisch australopithecusartig konfiguriert.
Auf Grundlage der Rekonstruktion des Lendenwirbels L3 ergeben sich mittels morphometrischer Prokrustusanalyse (PC*) anhand von 48 Messpunkten folgende Ergebnisse: Obwohl einige Gemeinsamkeiten zu Menschenaffen und auch Homininen bestehen, überraschen zwei Analysen (PC4 und 7-Fig. Suppl.), da Ausrichtung und Verjüngung der Rippenfortsätze (Processus costales) sowie Ausrichtung der Gelenkflächen von Australopithecus africanus und „insbesondere“ A. sediba nicht zu denen bei modernen Menschen und Neandertalern passen. Zudem sind die beiden Australopithecus-Arten in Bezug auf diese Merkmale einzigartig. Insgesamt vier Mal wurde von Williams et al. (2021, Fig. 7 A-D) ein gemeinsamer multimetrischer Raum aus je zwei PC-Analysen erstellt: Demnach lag die Wirbelform des Neandertalers immer im Variationsbereich des modernen Menschen. Die L3-Wirbel von Australopithecus sediba und africanus lagen einmal im Variationsbereich des Menschen, einmal knapp an dessen Rand und zweimal völlig isoliert. Alle PC-Analysen ergaben eine sehr große Nähe von Australopithecus sediba und africanus. Auch die Prokustusdistanzen (Fig. 7-Suppl. 1) zeigen klar, dass die Form des L3-Wirbels von A. sediba dem von A. africanus am stärksten ähnelt und weit vom Schimpansen, aber auch vom modernen Menschen und Neandertaler entfernt ist.
*PC ist die Abkürzung für Prokrustus-Analyse. Die Messpunkte der Fossilien werden aus verschiedenen Perspektiven in ihren Abständen gemessen und anschließend auf ihre Ähnlichkeit hin verglichen.
Außerdem ermittelten Williams et al. (2021, Fig. 7 Suppl. 3) noch den Einfluss der Größe auf die Form des L3-Wirbels. Während sich das Neandertalerfossil noch im Variationsbereich des Menschen befindet und dies ebenfalls für einen Teil der drei Gattungen heutiger Großaffen gilt, sind die L3-Wirbel von Australopithecus sediba und africanus im Verhältnis von Größe und Form weit vom Menschen entfernt und typisch schimpansenartig konfiguriert.
- Diskussion
Williams et al. (2021) versuchen bei Australopithecus sediba eine starke Krümmung der fragmentarisch vorliegenden Lendenwirbelsäule (Lendenlordorse) anhand der gemessenen Winkel abzuleiten. Deutlich negative Winkelwerte gibt es aber auch bei A. africanus, der Mehrheit heutiger Menschenaffen und bei einigen heutigen Menschen. Das verbietet wiederum Schlussfolgerungen über den aufrechten Gang. Über alle Merkmale der Lendenwirbelflächenverhältnisse und PC-Analysen hinweg sind die Konstellationen bei Australopithecus sediba nicht menschlicher als beim ca. 1 MrJ älteren A. africanus. Die untere Wirbelsäule von A. sediba ist damit insgesamt entgegen der Schlussfolgerung von Williams et al. (2021) eindeutig australopithecusartig und kann auch nicht als Weiterentwicklung der Zweibeinigkeit in Richtung Mensch interpretiert werden.
All das bedeutet, dass Lee Bergers spekulative Aussage, dass Australopithecus sediba „wie ein Mensch lief und wie ein Affe kletterte“, überhaupt nicht von der Datenlage gedeckt ist. Wenn er recht hätte, wäre es möglich, dass A. sediba menschlich (also ausdauernd und energiesparend) aufrecht gehen konnte und trotzdem extrem an das Klettern angepasste Oberkörper und Arme besaß. Dies ist in sich nicht gut begründet. Erstens gibt es auch im unteren Körperskelett genügend Merkmale, die gegen eine menschliche Fortbewegung sprechen, so dass „Australopithecus sediba wahrscheinlich von allen Australopithecinen am stärksten an ein Leben in Bäumen angepasst war“ (DeSilva et al. 2012 bei Brandt 2017a, S. 30). Und zweitens ist dies aufgrund des ans Klettern angepassten oberen Körperskeletts kaum sinnvoll vorstellbar. Brandt (2017a) zufolge ist der obere Brustkorb von Australopithecus sediba großaffenähnlich eng konfiguriert und das Schulterblatt großaffenähnlich hoch gelegen. A. sediba konnte „deshalb beim Gehen und schnellen Laufen die Arme nicht so effektiv schwingen wie der Mensch“, und der enge obere Brustkorb und der Schultergürtel haben zudem wohl die beim schnellen oder ausdauernden Gehen erforderliche Hebung des oberen Rippenbogens beim tiefen Einatmen eingeschränkt (ebd.).
Auch neuere Studien über A. sediba zeigen „vorwiegend primitive“ Morphologie beim oberen Körperskelett, die auf „beträchtliche Kletter- und Hangelfähigkeit hindeuten“ (Churchill et al. 2018). Ebenso betonen Holliday et al. (2018) nicht nur die geringe Körpergröße und das niedrige Körpergewicht von A. sediba, sondern auch Kletteranpassungen an Fuß und Armen und den Mangel an typisch menschlicher „Zweibeinigkeit über weite Strecken“ aufgrund kürzerer Beinknochen. Auch die Orang-Utan-ähnliche Trabekel-Knochenstruktur der Mittelhandknochen spiegelt nach Dunmore et al. (2020) ihre starke Rolle in einer zugreifenden Fortbewegung wider (auch trotz menschen- bzw. gorillaähnlicher Trabekelstruktur des Mittelhandknochens des Daumens). Prahabat et al. (2021) beschreiben ebenfalls die affenartige Gliedmaßenproportionen bei Australopithecus sediba sowie bei anderen Australomorphen (einschließlich „Homo“ habilis) im Gegensatz zu Homo erectus und auch zu dem fast doppelt so alten A. afarensis. Deshalb bezeichnen sie bei A. afarensis die zweibeinigen Anpassungen sogar als „plausiblere“ „unabhängige Entwicklung“ (also Konvergenz). Diese Sichtweise findet grundsätzlich bei Almécija et al. (2021) Unterstützung und stellt daher eine Ableitung einer gemeinsamen Evolutionsgeschichte mit den Menschen aufgrund „menschenähnlicher“ Merkmale bei Australomorphen generell in Frage. Auch die Zahnsteinanalysen von A. sediba weisen nach Henry et al. (2012) auf eine schimpansenartige Ernährung und Lebensweise bei A. sediba und Ardipithecus ramidus hin. Dafür sprechen auch das schimpansenähnlich kleine Gehirn (ca. 420 cm³) und die großaffenartige Hirnfurchenmusterung des Stirnlappens (vgl. Tobias 1987, Berger et al. 2010, Brandt 2021).
Zudem erwies sich Homo erectus mit 1,95 bis 2,04 MrJ entgegen der ursprünglichen Erwartung als etwas älter als Australopithecus sediba mit 1,98 MrJ (Herries et al. 2020; Pickering et al. 2011). Somit ist nach Prahabat et al. (2021) noch kein bisschen klar, wann und wie überhaupt ein Übergang „vom Baumleben zur dauerhaften Zweibeinigkeit“ stattgefunden haben könnte. Außerdem war H. erectus auch als ältester Hominine eindeutig an einen gewohnheitsmäßigen menschlichen Gang angepasst (vgl. Ruff 2009; Brandt 2017a). Der menschliche Gang von Homo erectus erscheint also plötzlich im Fossilbericht – und nicht schon bei Australopithecus sediba, wie Lee Berger behauptet hatte. Dies kann man auch als Hinweis auf eine getrennte Erschaffung deuten. Denn nach dem Grundtypmodell erschuf Gott den Grundtypen der echten Menschen (mit einem Formenspektrum im Bereich von Homo erectus, Neandertaler und dem modernen Menschen) mit menschlich-aufrechtem Gang einerseits, und ein oder mehrere Grundtypen australomorpher Großaffen einschließlich Australopithecus sediba mit jeweils ganz eigenem Fortbewegungsrepertoire andererseits.
- Literatur
Almécija S et al. (2021) Fossil apes and human evolution. Science 372, eabb4363, doi: 10.1126/science.abb4363.
Berger LR et al. (2010) Australopithecus sediba: A New Species of Homo-Like Australopith from South Africa. Science 328, 195–204, doi:10.1126/science.1184944.
Brandt M (2015) Flores-Mensch: Moderner Homo sapiens mit Down-Syndrom? Stud. Int. J. 22, S. 30–34.
Brandt M (2017a) Wie sicher sind Deutungen in der Paläanthropologie? Australopithecus sediba und sein merkwürdiges Merkmalsmosaik. In: Brandt M (Hg) Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special, S. 9–49.
Brandt M (2017b) Homo naledi – neuer Hominine mit vielen Fragezeichen. In: Brandt M (Hg) Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special, S. 51–79.
Brandt M (2021) Gab es Vormenschen? Irrungen und Wirrungen in der Paläoneurologie. W+W Special Paper B-21-4.
Churchill SE et al. (2018) The shoulder, arm, and forearm of Australopithecus sediba. PaleoAnthropology 2018:234–281, doi:10.4207/PA.2018.ART114.
Daegling DJ et al. (2016) Comparative biomechanics of Australopithecus sediba mandibles. J. Hum. Evo. 100, 73 –86, https://doi.org/10.1016/j.jhevol.2016.08.006.
Dönges J (2021) Früher Menschenverwandter: Fossile Wirbel von Australopithecus sediba gefunden. spektrum.de. Veröffentlicht am 24.11.2021, https://www.spektrum.de/news/fossile-wirbel-von-australopithecus-sediba-gefunden/1952386.
Dunmore CJ et al. (2020) The position of Australopithecus sediba within fossil hominin hand use diversity. Nature Ecology & Evolution 4, 911–918, doi: https://doi.org/10.1038/s41559-020-1207-5.
Hartwig-Scherer S (2005) Die Zwergenmenschen – Genetische Isolation führt zur Bildung von „Hobbits“ auf indonesischer Insel. Stud. Int. J. 12, S. 3–10.
Henry AG et al. (2012) The diet of Australopithecus sediba. Nature 487:90–93, https://doi.org/10.1038/nature11185.
Herries AIR et al. (2020) Drimolen crania indicate contemporaneity of Australopithecus, Paranthropus and early Homo erectus in South Africa. Science 368, eaaw7293, doi: 10.1126/science.aaw7293.
Holliday TW et al. (2018) Body size and proportions of Australopithecus sediba. PaleoAnthropology 2018:406–422, doi:10.4207/PA.2018.ART118.
Pickering RJ et al. (2011) Australopithecus sediba at 1.977 Ma and Implications for the Origins of the Genus Homo. Science 333, 1421–1423, doi:10.1126/science.1203697.
Prabhat AM et al. (2021) Homoplasy in the evolution of modern human-like joint proportions in Australopithecus afarensis. eLife 10:e65897, https://doi.org/10.7554/eLife.65897.
Ruff C (2009) Relative Limb Strength and Locomotion in Homo habilis. Am. J. Phys. Anthropol. 138:90–100, doi: 10.1002/ajpa.20907.
Tobias VP (1987) The brain of Homo habilis: A new level of organization in cerebral evolution. J. Hum. Evol. 16, 7–8, 741-761, https://www.sciencedirect.com/science/article/abs/pii/0047248487900224.
Williams SA et al. (2021) New fossils of Australopithecus sediba reveal a nearly complete lower back. Elife. 10, e70447, doi: 10.7554/eLife.70447.
Autor dieser News: Benjamin Scholl
© 2022, http://www.genesisnet.info/schoepfung_evolution/n298.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
29.09.20 Mann und Frau – auch im Gehirn unterschiedlich
Das menschliche Gehirn wird in verschiedenster Hinsicht intensiv diskutiert. Dabei geht es nicht nur um den Zusammenhang zwischen Geist und Gehirn, sondern z. B. auch um geschlechtsspezifische Ausprägungen des Gehirn. Letztere wurden in der Forschung bereits vielfach belegt und gelten entgegen mancher populären Diskussion als geklärt. Hier wird eine aktuelle Studie vorgestellt, die geschlechtsspezifische Merkmale im menschlichen Gehirn auch auf molekularer Ebene aufzeigt.
Das menschliche Gehirn wird intensiv untersucht mit dem Ziel, dieses einzigartige Organ in seinem Aufbau, seiner Struktur, seiner Funktion und bezüglich seiner Bedeutung für den Menschen besser zu verstehen. Konkrete Projekte sind z. B. das 2010 gestartete „Human Connectome Projekt“, in dem die Verknüpfungen aller Neuronen beim Menschen untersucht und dokumentiert werden. Das „Human Brain Project“ ist eine entsprechende europäische Initiative, die seit 2013 eine Plattform zur Bündelung der Erforschung des menschlichen Gehirns bietet. Mit all diesen Anstrengungen sind ähnlich wie bei der Erforschung des menschlichen Erbguts große Erwartungen verknüpft, dass mit zunehmendem Verständnis des menschlichen Gehirns etwas Wesentliches vom Menschen zugänglich würde. Ein geschlechtsspezifischer Unterschied in Aufbau und Funktion des humanen Gehirns (Sexualdimorphismus) wurde in der Geschichte seiner Erforschung nie in Frage gestellt. Das Ausmaß der Unterschiede, ihre Ursache und ihre Bedeutung werden nach wie vor kontrovers diskutiert (Sacher et al. 2013, de Lacy et al. 2019, Pallayova et al. 2019). Im Rahmen der erwähnten Projekte werden auch Gehirne von Tieren als Modell für das humane Gehirn untersucht, da diese experimentellen Methoden leichter zugänglich sind. Herausforderungen bestehen dann darin, die Übertragbarkeit der Erkenntnisse aus Tiermodellen auf das menschliche Gehirn zu prüfen. Am Gehirn von Mäusen sind geschlechtsspezifische Unterschiede gut untersucht und etabliert; dazu gehören (1) die Volumenverteilung der grauen Substanz in bestimmten Bereichen der Großhirnrinde (Kortex) und in subkortikalen Gehirnregionen; (2) die unterschiedlichen Volumina an grauer Substanz vor allem in Bereichen für soziales Verhalten und für die Fortpflanzung und (3) der Zusammenhang zwischen der räumlichen Verteilung der genannten Bereiche grauer Substanz und der Expression geschlechtschromosomaler Gene – also deren Konzentration – im Gehirn erwachsener Tiere.
Liu et al. (2020) haben nun eine Arbeit vorgelegt, in der sie die Übertragbarkeit dieser Befunde aus dem Tiermodell auf den Menschen geprüft haben. Dazu haben sie zunächst zwei unabhängige und umfangreiche Datensätze von Schichtaufnahmen (Scans) genutzt, die durch bildgebende Verfahren von menschlichen Gehirnen gewonnen worden waren. Ein Datensatz stammt aus dem Human Connectome Projekt (HCP), der andere aus der englischen UK Biobank (UKB); insgesamt wurden mehr als 2000 Scans in diese Untersuchung einbezogen. Die geschlechtsspezifische räumliche Verteilung der Volumina der grauen Masse erwies sich in diesen Datensätzen als in hohem Maße reproduzierbar.
Weiter untersuchten Liu et al. die bisher beschriebenen im Gehirn ausgeprägten Gene (Transkriptom) und verglichen diese Daten mit der geschlechtsspezifischen Verteilung der grauen Substanz. Das Allen Institute for Brain Science liefert einen öffentlich zugänglichen Atlas mit ca. 16.000 Genen, die im menschlichen Gehirn in RNA umgeschrieben (transkribiert) werden. Dies ermöglicht zu prüfen, ob Gene der X-Chromosomen bzw. des Y-Chromosoms bevorzugt in den geschlechtsspezifischen Bereichen des Gehirns transkribiert werden, wie das im Mausmodell nachgewiesen worden ist. Darüber hinaus können die im menschlichen Gehirn exprimierten Gene danach eingeteilt werden, inwieweit ihre Expression mit den geschlechtsspezifischen anatomischen Unterschieden in den Gehirnstrukturen gekoppelt ist.
Für die geschlechtsspezifischen strukturellen, anatomischen Unterschiede im menschlichen Gehirn führen die Autoren auch Erkenntnisse aus seltenen medizinischen Störungen an, die zeigen, dass Sexualhormone und die Menge an X- bzw. Y-chromosomalen Genen die Volumina der Gehirnbereiche beeinflussen, die in einer Population geschlechtsabhängige Anatomie aufweisen. Bei der Geburt werden ebenfalls geschlechtsspezifische Unterschiede in der menschlichen Gehirnanatomie beschrieben, zu diesem Zeitpunkt werden diese Differenzen vor allem durch die Gene und die Bedingungen in der Gebärmutter beeinflusst. Die Autoren resümieren zu diesem Aspekt Untersuchungen, wonach es kaum möglich ist, die geschlechtsspezifischen Unterschiede in der menschlichen Gehirnstruktur allein auf die Umgebungsbedingungen zurückzuführen.
Weiter bestätigt die Studie von Liu et al. (2020) eine enge Verknüpfung der geschlechtsspezifischen anatomischen Unterschiede und der Expression von Genen der X-Chromosomen bzw. des Y-Chromosoms. Dies war zuvor nur in Mäusen, einem Modellsystem für den Menschen, beschrieben worden. Die Autoren sehen in diesem Befund auch einen Hinweis darauf, dass Steroide aus den Geschlechtsdrüsen nicht die einzigen Einflussgrößen bei der Musterbildung und geschlechtsspezifischen Verteilung der grauen Masse im menschlichen Gehirn sind.
Fazit. Auch wenn nach wie vor viele Fragen zum Unterschied im Gehirn von Mann und Frau offen bleiben und der experimentellen Untersuchung menschlicher Gehirne enge Grenzen gesetzt sind, so haben Liu et al. mit ihrer aktuellen Arbeit doch eindrucksvoll bestätigt, dass es eine Fülle von Befunden gibt, die für eine geschlechtsspezifische Unterscheidung des männlichen und weiblichen Gehirns sprechen. Wenn dann in der Titelformulierung einer populären Zusammenfassung – Überraschung anzeigend – formuliert wird: „Gehirn von Mann und Frau ist doch unterschiedlich“ (Podbregar 2020), dann zeugt das entweder von Ignoranz oder massiver weltanschaulicher Voreingenommenheit. Denn das ist, wie oben erwähnt, bereits lange bekannt und wird durch die hier vorgestellte Untersuchung nur einem konkreten Zusammenhang vertieft.
Dass bei der Übertragung von tierischen Modellen auf den Menschen Vorsicht geboten ist, zeigen Schaeffer et al. (2020) in einer aktuellen Untersuchung. Demnach weisen die Gehirne von Nagetieren und Primaten zwar eine vergleichbare Architektur auf, aber bei Mäusen, Krallenäffchen und Mensch sind funktionale Bereiche im Frontallappen unterschiedlich verschaltet. Das unterstreicht die Notwendigkeit, an Tiermodellen gewonnene Erkenntnisse vor der Übertragung auf den Menschen gründlich zu prüfen, wie das im Falle von Liu et al. (2020) vorbildlich gemacht worden ist.
Literatur
de Lacy N, McCauley E, Kutz JN & Calhoun VD (2019) Multilevel mapping of sexual dimorphism in intrinsic functional brain networks. Front. Neurosci. 13:332. doi: 10.3389/fnins.2019.00332
Liu S, Seidlitz J, Blumenthal JD, Clasen LS & Raznahan A (2020) Integral structural, functional, and transcriptomic analyses of sex-biased brain organization in humans. Proc. Natl. Acad. Sci. USA; www.pnas.org/cgi/doi/10.1073/pnas.1919091117
Pallayova M, Brandeburova A & Tokarova D (2019) Update on sexual dimorphism in brain structure-function interrelationships: a literature review. Appl. Psychophysiology & Biofeedback 44, 271-284.
Podbregar N (2020) Gehirn von Mann und Frau ist doch verschieden. https://www.scinexx.de/news/biowissen/gehirn-von-mann-und-frau-sind-doch-verschieden/
Sacher J, Neumann, Okon-Singer H, Gotowiec S & Villringer A (2013) Sexual dimorphism in the human brain: evidence from neuroimaging. Magnetic Resonance Imaging 31, 366-375.
Schaeffer DJ, Hori Y, Gilberrt KM, Gati JS, Menon RS & Everling S (2020) Divergence of rodent and primate medial frontal cortex functional connectivity. Proc. Natl. Acad. Sci. USA; www.pnas.org/cgi/doi/10.1073/pnas.2003181117
Autor dieser News: Harald Binder
© 2020, http://www.genesisnet.info/schoepfung_evolution/n281.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
05.06.20 Das HAR1F-Gen stellt Evolution in Frage
Unerwartete Befunde bieten eine gute Möglichkeit, etablierte Hypothesen zu testen und ggf. nach alternativen Erklärungen zu suchen. Ein Beispiel aus der vergleichenden Genetik sind sogenannte HAR-Gene („Human Accelerated Regions“). 2700 solche Gene sind beim Menschen bekannt. Eines von ihnen, das HAR1F-Gen, unterscheidet sich in 18 Nukleotiden vom Gen der Schimpansen und der anderen Menschenaffen. Eine Untersuchung anhand bekannter Mutationsraten und der Populationsgenetik macht es unplausibel, dass das Gen ein Produkt ungerichteter Evolution ist.
HAR steht für „Human Accelerated Region“. Im menschlichen Genom sind 2700 solcher HAR-Gene bekannt. Die Bezeichnung spiegelt wider, dass sich diese Gene beim Menschen stark von denjenigen anderer Primaten unterscheiden. Daher sollten sie nach der Evolutionstheorie sehr schnell mutiert und selektiert worden, also durch sehr schnelle Evolution entstanden sein (daher „accelerated“). HARs unterscheiden sich in der Tat deutlich von den homologen Genen bei Primaten.
Das HAR1F-Gen codiert nicht für ein Protein, sondern für ein sog. long-non-coding RNA-Molekül mit einer Regulationsfunktion. Die entsprechende RNA reguliert ein genetisches Programm während der embryonalen Entwicklung des Gehirns. Es wird in sogenannten Cajan-Retzius-Zellen, die für die Produktion der dicken Großhirnrinde verantwortlich sind, produziert.
Wie die anderen HAR-Gene ist auch das HAR1F-Gen durch einen sogenannten genetischen Fingerabdruck gekennzeichnet, der nur beim Menschen vorkommt. Das HAR1F-Gen besteht aus 118 Nukleotiden (DNA-Bausteinen), von denen 18 für den Menschen einzigartig sind. Die Sequenz des HAR1F-Gens ist bei Schimpanse, Gorilla und Orang-Utan identisch. Nur beim Menschen werden die 18 einzigartigen Nukleotide gefunden. Diese 18 Unterschiede sind über das gesamte Gen verteilt und können nicht auf ein einziges Mutationsereignis zurückgeführt werden. Basierend auf der Sequenz des HAR1F-Gens können wir aufgrund der großen Unterschiede mit Sicherheit Menschen von Schimpansen, Gorillas und anderen Affen unterscheiden. Wenn wir ein solches Gen in einem fossilen Knochen (z. B. eines Neandertalers) finden, dann können wir sicher sein, dass wir es mit einem Menschen zu tun haben. Das HAR1F-Gen ist also ein Indikatorgen, d.h. ein Gen, das anzeigt, dass wir es mit einem Menschen zu tun haben. Die einzigartige DNA-Sequenz des menschlichen HAR1F-Gens führt zur Faltung des entsprechenden RNA-Moleküls, so dass eine neue, winzig kleine Schleife entsteht, die die Funktion des HAR1F-Moleküls bestimmt. Diese Schleife ist eine Art Schalter für das Entwicklungsprogramm der Großhirnrinde beim Fötus.
Nach dem Neodarwinismus ist das HAR1F-Gen Schritt für Schritt, d.h. Buchstabe um Buchstabe, durch zufällige, selektierbare Mutationen entstanden. Durett et al. (2008) haben berechnet, dass für das Auftreten einer einzigen Mutation an einer bestimmten Position in einem Gen zur Bildung einer neuen funktionellen Stelle Millionen von Jahren erforderlich wären. Trifft das auch für das HAR1F-Gen zu? Wir brauchen also 18 Mutationen, um eine affenähnliche HAR1F-Sequenz in ein menschliches HAR1F-Gen zu verändern.
Dazu bedarf es einer Anhäufung von Mutationen an ganz bestimmten Stellen im Genom des hypothetischen Vorfahren. Da der mutmaßliche Vorfahr des Menschen und der Schimpansen vor 6-7 Millionen Jahren gelebt haben soll, haben wir maximal 7 Millionen Jahre Zeit, um das menschliche HAR1F-Gen zu erhalten. Neodarwinisten gehen davon aus, dass die effektive Population von Homininen (Menschen und seine unmittelbaren mutmaßlichen Affen-Vorfahren) während dieser 7 Millionen Jahre etwa 10.000 betrug (Wall 2003). Und die Mutationsrate ist experimentell ermittelt worden. Sie beträgt 100 Punkt-Mutationen pro Generation pro Genom (das sind beim Menschen etwa 3 Milliarden DNA-Buchstaben). Mit anderen Worten: Jeder Nachkomme erhält 100 Mutationen von seinen Eltern. Auch Affenjunge und ihre Vorfahren.
Mit diesen Daten können wir auf ziemlich einfache Weise berechnen, ob das menschliche HAR1F-Gen auf darwinistische Weise – durch Mutation, Selektion und Gendrift – entstehen konnte oder nicht. Wie groß ist die Chance, dass wir eine Mutation an der richtigen Stelle im HAR1F-Gen bekommen?
Pro Individuum besteht die Chance von 1 zu 30 Millionen, dass eine der oben erwähnten 100 Mutationen pro Generation an die richtige Stelle fällt (100/3 Milliarden = 1/30 Millionen). Die Chance, dass dies einmal bei einer Population von 10.000 Menschen (oder ihren mutmaßlichen Vorfahren) geschieht, liegt demnach bei 1/3000. Mit anderen Worten, alle 3000 Generationen wird es durchschnittlich einmal passieren. Wenn wir durchschnittlich 10 Jahre für eine Generation rechnen, dann dauert es 30.000 Jahre, um einmal einen Treffer zu haben. Aber ist es der richtige Treffer? Es muss auch der richtige DNA-Buchstabe sein (die DNA hat vier verschiedene Nukleotide).1 In zwei von drei Fällen ist es der falsche Buchstabe. Bevor wir also den ersten richtigen Treffer landen, sind 30.000–90.000 Jahre vergangen!
Erst jetzt beginnt es wirklich spannend zu werden. Denn wir haben zwar den ersten Treffer gelandet, aber was wird mit dieser Mutation geschehen? Wenn es sich um eine neutrale Mutation handelt, wird sie normalerweise aufgrund der zufälligen genetischen Drift verloren gehen. Nach Ansicht der Populationsgenetiker hat jede neutrale Mutation nur die minimale Chance von 1/2N (mit N = Populationsgröße), dass diese Mutation nicht wieder verloren geht! Nachdem eine Mutation endlich an der richtigen Stelle aufgetreten ist, ist die Chance, dass sie tatsächlich in der Population erhalten bleibt, also minimal (1/20.000 bei einer Populationsgröße von 10.000 Individuen). Hier braucht man daher die natürliche Auslese. Wir müssen also davon ausgehen, dass die Punktmutation (Austauschs eines Nukleotids) im HAR1F-Gen des Vorfahren (das bei den oben erwähnten Affen monomorph ist, also bei allen Individuen identisch und nicht mutiert) einen selektiven Wert hat. Das ist durchaus möglich. Geben wir dieser Mutation einen Selektionsvorteil von 0,5% gegenüber dem Gen des Vorfahren, ist das „leicht vorteilhaft“, aber dennoch sehr großzügig für eine Punktmutation in einem stabilen Gen (eher erwartet man einen Nachteil). Die Mutation hat nach populationsgenetischen Berechnungen nun eine Chance von 1%, sich in der Bevölkerung zu etablieren. Ein Treffer muss also durchschnittlich 100 Mal erfolgen, um sich dauerhaft einmal in der Population anzusiedeln. Der erste richtige Treffer tritt erst nach 30.000–90.000 Jahren ein, aber in der Population muss dies durchschnittlich 100 Mal geschehen! 30.000–90.000 x 100 ergibt 3–9 Millionen! Es dauert daher 3–9 Millionen Jahre, um eine Mutation mit 0,5% Selektionsvorteil durch Selektion dauerhaft im HAR1F-Gen zu erhalten! Es werden aber noch weitere 17 Mutationen im HAR1F-Gen benötigt.
Ein Kommentar in der Wissenschaftszeitschrift Nature bestätigt, dass das HAR1F-Gen durch Mutation/Selektion schwer zu erklären ist: „Es wird angenommen, dass die Rekombination und der damit verbundene Prozess, die verzerrte Genkonversion („biased gene conversion“), die Aufnahme von G- und C-Nukleotiden gegenüber den beiden anderen möglichen Nukleotiden, A und T, begünstigt […]. Da alle bei HAR1F beobachteten Nukleotidsubstitutionen von diesem Typ sind, könnten hohe (und verzerrte) Mutationsraten einen Teil der raschen Evolution von HAR1F erklären. Dennoch kann dieser Prozess nicht die anderen Beobachtungen der Autoren erklären, wie z. B. die Substitutionspaare, die zusammen die Struktur der HAR1F-RNA weiter stabilisieren“ (Ponting & Lunter 2006).
Zufällige Mutationen, Selektion und Gendrift können das menschliche HAR1F-Gen nicht erklären. Und es gibt 2700 weitere HAR im menschlichen Genom, wo es ähnlich große Unterschiede zum Schimpansengenom gibt wie beim HAR1F-Gen! Was hat das also mit unserer eigenen Existenz zu tun? Die Antwort ist, dass es eine wichtige Einschränkung dessen darstellt, was wir aus den Ähnlichkeiten von Menschen und Menschenaffen ableiten können: Ähnlichkeiten sind als Belege für gemeinsame Abstammung fragwürdig, wenn zugleich markante Unterschiede vorliegen (vgl. Terborg 2019). Insbesondere zeigt dieses Beispiel, dass wir aus der Sequenz in Primaten nicht ableiten können, dass die darwinistische Evolution die Sequenz in Menschen hervorgebracht hat, nur weil sie ähnlich sind. Eine schrittweise Evolution ist sicherlich nicht die richtige Erklärung unserer Existenz!
Quellen
Durett R & Schmidt D (2008) Waiting for two mutations: with applications to regulatory sequence evolution and the limits of Darwinian evolution. Genetics 180, 1501-1509.
Ponting CP & Lunter G (2006) Evolutionary biology: human brain gene wins genome race. Nature 443, 149–150.
Terborg P (2019) Das Erbgut von Mensch und Schimpanse. Wie groß ist die genetische Verwandtschaft wirklich? Stud. Integr. J. 26, 4-10.
Wall JD (2003) Estimating ancestral population sizes and divergence times. Genetics 163, 395–404.
Anmerkung
1 Man könnte argumentieren, dass es nicht genau die heutige Sequenz sein muss. Dennoch müsste man eine ähnliche Schleife erklären, die ebenfalls durch paarweise positionierte Nukleotide stabilisiert werden muss.
Autor dieser News: Peter Borger
© 2020, http://www.genesisnet.info/schoepfung_evolution/n280.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
09.03.15 Bindeglied zwischen „Affenmensch“ und Mensch?
Ein fossiles Unterkieferbruchstück wird als Beleg für die Existenz eines Zwischenwesens zwischen Menschenaffen und Menschen interpretiert und öffentlichkeitswirksam mit dieser Deutung präsentiert. Doch es gibt gute Gründe dafür, dass der neue Fund nicht zwischen Affen und Menschen anzusiedeln ist, sondern zu einer Variante der ausgestorbenen Großaffengattung Australopithecus gehört.
Ein internationales Forscherteam hat im Wissenschaftsmagazin Science ein neues fossiles Unterkieferbruchstück aus Äthiopien vorgestellt, das ein radiometrisches Alter von ca. 2,8 Millionen Jahren hat. Die Forscher stellten das Fundstück zur Gattung Homo, zu der auch der moderne Mensch gehört. Die menschliche Linie sei mit diesem neuen Fossil 400.000 Jahre älter als bisher gedacht (Villmoare et al. 2015).
Laut den Forschern besitzt der Unterkiefer eine Reihe von Eigenschaften, die ihn zwischen Australopithecus und den jüngeren Homo habilis stellt (Villmoare et al. 2015). Die Australopithecinen werden im Evolutionsmodell als „Vormenschen“ oder „Affenmenschen“, im Grundtypmodell der Schöpfungslehre dagegen als ausgestorbene Großaffen gedeutet. Im Evolutionsmodell stellt der neue Unterkiefer somit ein Bindeglied zwischen „Affenmenschen“ und echten Menschen dar. Der Fund hat zwar große publizistische Resonanz gefunden, der wichtigste Punkt im Zusammenhang mit der Einordnung des neuen Fossils wurde aber weder im Originalartikel noch in der populären Verbreitung gestellt: War Homo habilis wirklich ein Mensch?
Seit Aufstellung der Art Homo habilis im Jahre 1964 gibt es daran berechtigte Zweifel. Erst in neuerer Zeit haben Paläanthropologen wieder für eine Ausgliederung der Art habilis und auch der Art rudolfensis aus der Gattung Homo mit sehr guten Argumenten plädiert (Übersicht bei Brandt 2012). Trotz der guten Argumente halten jedoch fast alle Urgeschichtsforscher an der alten Zuordnung zur Gattung Homo fest. Was könnten Gründe dafür sein?
„Homo“ habilis/rudolfensis wird seit Jahrzehnten als eine evolutionär vermittelnde Fossilgruppe zwischen den Australopithecinen und dem frühesten unbestritten echten Menschen (Homo erectus) verstanden. Mit ihrer Rückstufung erscheint die Lücke zwischen den recht großaffenähnlichen Australopithecinen und dem echten Menschen (Homo erectus und spätere Homo-Formen) größer. Auch für den Laien erkennbar ergibt sich damit eine Unterteilung der fossilen Formen in zwei Gruppen: echte Menschen, zu denen Homo erectus, Neandertaler u.a. gehören, die in der Schöpfungslehre dem Grundtyp Mensch zugeordnet werden, und großaffenähnliche Tiere wie die Australopithecinen (Brandt 2012).
Das neu entdeckte Unterkieferbruchstück aus Äthiopien gehört somit zu keinem neuen Bindeglied zwischen „Vormenschen“ und echten Frühmenschen, sondern wahrscheinlich zu einem weiteren Vertreter der Gattung Australopithecus. Der Paläanthropologe Christoph Zollikofer aus Zürich meint sogar, dass der Unterkiefer gar nicht zu einer neuen unbekannten Art, sondern zu den bekannten Australopithecinen gehört.1
Anmerkung
1 http://www.nzz.ch/wissenschaft/medizin/bindeglied-zwischen-homo-und-australopithecus-1.18495357 (Zugriff am 5.3.2015).
Literatur
Brandt M (2012) „Homo“ habilis war kein Mensch. Studium Integrale Journal 19, 4 -11. http://www.wort-und-wissen.de/sij/sij191/sij191-1.html
Villmoare B, Kimbel WH, Seyoum C, Campisano CJ, DiMaggio E, Rowan J, Braun DR, Arrowsmith JR & Reed KE (2015) Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. http://www.sciencemag.org/content/early/2015/03/03/science.aaa1343
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2015, http://www.genesisnet.info/schoepfung_evolution/n224.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
16.05.13 „Vormensch“ verliert seinen Status
Australopithecus erweist sich als guter Kletterer
Die Australopithecinen werden trotz ihrer unbestritten recht großaffenähnlichen Schädel- und Zahnstruktur als „Vormenschen“ gedeutet, denn Körperstamm- und Extremitätenskelett seien schon deutlich zum Menschen hin evolviert. Diese behauptete Menschenähnlichkeit wird durch neue Untersuchungen in Frage gestellt. Sie unterstützen frühere Forschungsergebnisse, nach denen diese fossilen Tiere keine Ahnen des Menschen sind.
Die nur fossil bekannte Menschenaffengattung Australopithecus („Südaffe“) gilt weithin als relativ gut passende Übergangsform zwischen Menschenaffen und Menschen und wird daher auch als „Vormensch“ und in populären Darstellungen sogar (irreführenderweise) als „Urmensch“ bezeichnet. Dass eine Reihe von Merkmalen diesem Status widersprechen, wird im Artikel Australopithecinen („Südaffen“) und andere Menschenaffenartigen-Fossilien, https://www.genesisnet.info/index.php?Artikel=43622&Sprache=de&l=1 behandelt. Diese in der Fachwelt teilweise auch kritisierte Position von Australopithecus wird durch neue Befunde weiter in Frage gestellt.
Die beiden Forscher David J. Green und Zeresenay Alemseged haben 2012 in der Wissenschaftszeitschrift Science eine bemerkenswerte Untersuchung zum Schulterblatt von Australopithecus afarensis publiziert. Mit dieser Analyse wurde eine seit Jahrzehnten kontrovers diskutierte Frage zur Fortbewegung der Australopithecinen vermutlich entschieden. Es spricht vieles dafür, dass diese Tiere einen wesentlichen Teil ihres Lebens in Bäumen verbracht haben. Damit muss eines der tragenden Indizien für eine Übergangsstellung von Australopithecus fallengelassen werden.
Es war schon zuvor unbestritten, dass Australopithecus afarensis im Schulterbereich großaffenähnliche Merkmale besitzt. Manche Forscher aber interpretierten die Klettermerkmale einfach als funktionslose Überbleibsel von Vorläuferformen – eine recht willkürliche und kaum prüfbare Interpretation. Andere sahen in den großaffenähnlichen Merkmalen der oberen Extremitäten von Australopithecus eine Anpassung an eine Fortbewegung in Bäumen.
Das Schulterblatt (Scapula) ist ein Schlüsselknochen in der Beurteilung der Fortbewegungsweise von Primaten. Aber dieses Knochenelement ist fossil sehr selten und wenn nur bruchstückhaft erhalten. Mit der Entdeckung des Australopithecus afarensis-Teilskelettes von Dikika (DIK-1-1), Äthiopien, veränderte sich die Datenlage grundlegend. Man besitzt nun ein nahezu vollständig erhaltenes rechtes Schulterblatt eines juvenilen (kindlichen) Individuums von Australopithecus.
Die Untersuchung von Green & Alemseged (2012) zeigt, dass das Schulterblatt von Australopithecus eine vom Menschen Homo deutlich verschiedene großaffenähnliche Struktur besitzt (Verlauf des Knochenkamms auf der Rückseite des Schulterblattes und die Ausrichtung der Schultergelenkspfanne). Die Forscher konnten nachweisen, dass die Ausrichtung der Schultergelenkspfanne von „Lucy“ nicht wie behauptet durch die geringe Körpergröße bedingt ist.
Arias-Martorell et al. (2012) haben in einer jüngsten Analyse das proximale (körpernahe) Ende des Oberarmknochens (Humerus) der lebenden Großaffen und Menschen sowie von Australopithecus afarensis untersucht. Es wurden signifikante Unterschiede zwischen diesen Lebewesen, die bisher nicht bekannt waren, festgestellt. Die proximale Humerus-Morphologie des Menschen unterscheidet sich deutlich von der der Großaffen und von Australopithecus afarensis, wobei letztere dem Orang-Utan am ähnlichsten sind!
Interessant ist, dass bereits vor vier Jahrzehnten Forscher um Charles Oxnard anhand eines bruchstückhaft erhaltenen Schulterblatts von Australopithecus africanus und eines Teils eines Schlüsselbeines von „Homo“ (Australopithecus) habilis zu ähnlichen Ergebnissen wie Green & Alemseged (2012) und Arias-Martorell et al. (2012) gekommen sind.
Bewertung
Die großaffenähnliche Schulter von Australopithecus ist durch eine große morphologische Kluft von der menschenähnlichen Schulter des fossil frühesten nachgewiesenen echten Menschen Homo ergaster getrennt (vgl. Stufen des Menschen?, https://www.genesisnet.info/index.php?Artikel=43623&Sprache=de&l=1). Die Schultermorphologie von Australopithecus bestätigt die Hypothese (Brandt 1995), dass diese fossilen Wesen sich nicht auf einem evolutionären Entwicklungsweg hin zum Menschen befanden, sondern ausgestorbene Großaffen repräsentieren, die in keinem historisch-verwandtschaftlichen Verhältnis zum echten Menschen stehen.
Eine ausführliche Darstellung der hier kurz zusammengefassten Befunde wird in Studium Integrale Journal Jg. 20, Heft 1 (voraussichtlich Juni 2013) veröffentlicht.
Literatur
Arias-Martorell J, Potau JM, Bello-Hellegouarch G & Pérez-Pérez AM (2012) 3D geometric morphometric analysis of the proximal epiphysis of the humerus of hominoids: Functional interpretation, locomotor evolution and variability. Am. J. Phys. Anthropol. 47, Suppl. 54, 85.
Brandt M (1995) Der Ursprung des aufrechten Ganges. Neuhausen-Stuttgart.
Green DJ & Alemseged Z (2012) Australopithecus afarensis scapular ontogeny, function, and the role of climbing in human evolution. Science 338, 514-517.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2013, http://www.genesisnet.info/schoepfung_evolution/n197.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
18.04.13 Endogene Retroviren – ein Beweis für gemeinsame Abstammung?
Im Erbgut des Menschen gibt es Abschnitte, die dem Genom von Retroviren auffallend ähnlich sind und an die Nachkommen vererbt werden. Solche endogenen Retroviren findet man ebenfalls im Genom anderer Primaten und zwar häufig an derselben Position und mit denselben Änderungen durch Mutationen. Dieser Befund wird als „Beweis“ für gemeinsame Vorfahren der Primaten präsentiert. Bei dieser Interpretation wird jedoch vernachlässigt, dass es in den verschiedenen Primaten auch spezifische retrovirale Sequenzabschnitte gibt und diese nicht alle stillgelegt sind; bei einzelnen sind Funktionen nachgewiesen worden. Möglicherweise sind endogene Retroviren gar nicht das, wonach sie auf den ersten Blick aussehen.
Im menschlichen Genom gibt es viele Abschnitte, die sich nicht permanent an derselben Stelle befinden, sondern mobil im Genom* ihre Position ändern können. Diese mobilen Bereiche werden als Transposons (transposable elements) bezeichnet und wurden 1948 von McClintock erstmals in Farbvarianten von Mais beschrieben (Nobelpreis 1983). Manche Transposons bewegen sich im Genom autonom, sie enthalten alle nötigen Voraussetzungen dazu, während andere auf Komponenten aus dem gesamten Genom angewiesen sind.
*Genom (= gesamtes Erbgut einer Art)
Zu den autonomen mobilen Elementen gehören auch endogene Retroviren. Darunter versteht man Abschnitte im Genom, deren Nukleotidsequenz derjenigen von Viren auffallend ähnlich oder sogar damit identisch ist. Die entsprechenden Retroviren weisen ein Genom aus RNA auf (im Gegensatz zum typischen DNA-Genom). Daher muss bei einer solchen Virus-Infektion das RNA-Genom vor der Integration (also dem Einbau) ins Genom der infizierten Zelle durch ein virales Enzym, nämlich der reversen Transkriptase in DNA umgeschrieben werden. (Die Bezeichnung „reverse Transkriptase“ bringt genau das zum Ausdruck: Statt des üblichen Umschreibens – Transkribierens – von DNA in mRNA geht es hier genau umgekehrt – revers.) Man nimmt an, dass Retroviren in der Vergangenheit Keimbahnzellen (Ei oder Samenzellen) infiziert und sich in deren Genom integriert haben. Somit werden sie von einer Generation an die nächste vererbt; diesen Status bezeichnet man als endogen.
Bis zu 8 % des menschlichen Genoms bestehen aus endogenen Retroviren. Einige davon wurden bereits als funktional beschrieben, die meisten aber werden nach derzeitigem Kenntnisstand als stillgelegt betrachtet, es ist bisher keine Funktion bekannt (Sugimoto & Schust 2009).
Retroviren und allgemein Transposons integrieren sich an sehr vielen unterschiedlichen Positionen im Genom. Aus Studien über krebsauslösende Viren ist bekannt, dass im Anfangsstadium der Infektion Retroviren an vielen unterschiedlichen Stellen im Genom auftreten, im Krebsgewebe aber findet sich das virale Genom in allen Zellen an derselben Position. Alle Krebszellen des Tumors sind also aus einer Zelle hervorgegangen, in deren Genom das Virus sich an bestimmten Stellen integriert hatte und dort ungehemmtes Wachstum auslöste.
Genomanalysen haben gezeigt, dass im Erbgut von Menschen und anderen Primaten dieselben Retroviren an denselben Positionen vorkommen und auch weitgehend dieselben Veränderungen durch Mutationen aufweisen. Im letzteren Fall könnten z. B. Defekte, die das virale Genom stilllegen, im gemeinsamen Vorfahren entstanden und dann vererbt worden sein. Dieser Befund passt gut zu einer Vorstellung, nach der Mensch und andere Primaten in ihrer Entstehung auf gemeinsame Vorfahren zurückgehen und unterstützt somit das Evolutionskonzept.
Es gibt aber auch retrovirale Sequenzen, die an unterschiedlichen Stellen beim Menschen und Primaten liegen und die Erklärungen komplizierter machen. Entweder sind sie erst später, also nach der Artaufspaltung entstanden oder eine Spezies hat sie im Verlauf der getrennten Entwicklung verloren. Außerdem setzt die oben skizzierte Erklärung eines gemeinsamen Vorfahren von Mensch und Primaten voraus, dass die retroviralen DNA-Abschnitte zutreffend als Relikte historischer Infektionen durch Retroviren interpretiert werden. Derzeit werden diese Abschnitte des Genoms als größtenteils funktionslos betrachtet. Wenn man sich aber unser begrenztes Verständnis des Humangenoms vergegenwärtigt und berücksichtigt, dass zudem durch das ENCODE-Projekt Hinweise dafür vorgelegt wurden, dass mehr als 80 % des gesamten Erbguts funktional sind, dann gibt dies Anlass, die „Beweiskraft“ der endogenen Retroviren für gemeinsame Vorfahren zurückhaltender einzuschätzen. Es könnte sein, dass zukünftige Untersuchungen für retrovirale Abschnitte noch mehr Funktionen (Sharif et al. 2013) belegen und damit andere Erklärungen erforderlich machen.
Die weitere Forschung muss zeigen, ob sich das Argument der vererbten Defekte bestätigt oder ob es durch ein besseres Verständnis möglicher Funktionen von endogenen Retroviren geschwächt wird und möglicherweise gar keine „Defekte“ vorliegen. Daher ist die plakative Behauptung, mit den endogenen Retroviren einen Beweis für die Abstammung des Menschen von affenartigen Vorfahren zu haben, nicht gerechtfertigt. Die derzeitigen Kenntnisse über endogene Retroviren ermöglichen für den Moment tatsächlich ein gutes Argument für Evolution – nicht mehr und nicht weniger. Schließlich muss auch bedacht werden, dass die Frage nach der Plausibilität einer Abstammungsbeziehung von Affen und Menschen nicht nur anhand eines einzigen Befundkomplexes – hier die endogenen Retroviren – beurteilt werden sollte.1
Anmerkung
1 vgl. Wie ähnlich sind Mensch und Schimpanse?, https://www.genesisnet.info/index.php?News=147; Über den genetischen Unterschied zwischen Mensch und Schimpanse – der „1 %-Mythos“, https://www.genesisnet.info/index.php?News=87; Mensch und Schimpanse noch weiter auseinander, https://www.genesisnet.info/index.php?News=98.
Literatur
Cowley M & Oakey RJ (2013) Transposable elements re-wire and fine-tune the transcriptome. PLOS Genet. 9(1) e1003234. doi: 10.1371/journal.pgen.1003234.
Stoye JP (2012) Studies of endogenous retroviruses reveal a continuing evolutionary saga. Nature Rev. Microbiol. 10, 395-406.
Sharif J, Shinkay Y & Koseki H (2013) Is there a role for endogenous retroviruses to mediate long-term adaptive phenotypic response upon environmental inputs? Phil. Trans. R. Soc. B, doi.org/10.1098/rstb.2011.0340.
Sugimoto J & Schust DJ (2009) Review: Human endogenous retroviruses and the placenta. Reprod. Sci. 16, 1023-1033.
Autor dieser News: Harald Binder
© 2013, http://www.genesisnet.info/schoepfung_evolution/n195.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.04.13 Schimpanse und Mensch: Affen äffen nicht nach
In Veröffentlichungen über vergleichende Verhaltensstudien werden häufig die Ähnlichkeit zwischen Schimpansen und Menschen herausgestellt. Eine Studie des Verhaltensbiologen Claudio Tennie stellt diese Ähnlichkeiten in Bezug auf eine wesentliche Fähigkeit massiv in Frage: Schimpansen sind nicht zum Lernen durch Nachahmen in der Lage. Diese These hat weitreichende Folgen. Tennies Ergebnisse werfen auch Fragen zum Wissenschaftsbetrieb auf: Gibt es eine Selektion bei der Veröffentlichung von Forschungsergebnissen?
In den Kontroversen um Schöpfung und Evolution gehören die Ähnlichkeiten von Menschen und Menschenaffen zu den Befunden, die sich eines besonders großen Publikumsinteresses erfreuen. Das ist so seit Charles Darwin und erhielt in jüngster Zeit vor allem durch die Genomforschung Auftrieb. Die SG Wort und Wissen plant dazu in ihrer Zeitschrift Studium Integrale Journal eine Artikelserie, die mit der nächsten Ausgabe beginnen wird.
Ein weiteres Gebiet, auf dem die besondere Nähe des Menschen zum Tierreich behauptet wird, ist die Verhaltensbiologie. In der Öffentlichkeit wird meistens ein Bild von den Menschenaffen gezeichnet, wonach diese den Menschen sehr ähnlich seien; mit dem Fortschritt der Forschung zeigen sich aber markante qualitative Unterschiede. Von den Menschenaffen werde ein sehr menschliches Bild gezeichnet, schreibt ZEIT ONLINE-Autor Frederik Jötten: „[S]ie trauern, trösten, malen, artikulieren, benutzen Werkzeuge und verfügen über eine Kultur“. Doch dieses Bild stimmt wahrscheinlich nicht.
Claudio Tennie, Verhaltensforscher vom Max-Planck-Institut für evolutionäre Anthropologie in Leipzig, hat dieses Bild durch eine aufwändige Studie überprüft. Vor elf Jahren habe er als Diplomand seine Arbeiten mit dem Ziel begonnen, zu zeigen, wie ähnlich Schimpansen den Menschen sind, so Jötten – und veröffentlichte nun ein Ergebnis, das seinen Erwartungen klar widersprach.
Tennie untersuchte freilebende Schimpansen auf ihre Fähigkeit hin, Verhalten anderer Schimpansen nachzuahmen. Menschen lernen ständig durch Nachahmen – tun Schimpansen das auch? Um das herauszufinden, brachte Tennie in einem zweistufigen Versuch einem Schimpansen zunächst eine bestimmte in der Natur nicht vorkommende Geste bei, um im zweiten Schritt zu sehen, ob andere Affen diese Geste nachahmen. Er wählte als Geste eine Gebetshaltung aus und konditionierte das Tier darauf, indem er es mit einer Erdnuss belohnte, wenn es diese Haltung einnimmt. Dann wird ein weiterer Affe hinzugebracht, der den ersten eine Weile dabei beobachtet, wie er zu den Erdnüssen gelangt. Als dieser zweite Affe dann alleine gelassen wird, kommt er nie auf die Idee, mit einer nachgeahmten Gebetshaltung ebenfalls zur ersehnten Erdnuss zu kommen. Er kreischt und tritt gegen die Tür, da er offensichtlich an die Erdnuss gelangen will, aber es kommt ihm nicht in den Sinn, zu diesem Zweck den ersten Affen nachzuahmen.
Insgesamt wurden 15 Tiere auf diese Weise getestet; bis zu 170 Beobachtungen des Vorbilds halfen nicht; kein Affe ahmt den anderen nach. Eine verbreitete Überzeugung der Verhaltensforscher scheint widerlegt zu sein: Schimpansen beherrschen das sprichwörtliche „Nachäffen“ gerade nicht. Tennie war mit der Absicht angetreten, den Beweis der Nachahmefähigkeit anzutreten, und bewies stattdessen das Gegenteil. Schimpansen schauen auch nützliche Handgriffe wie eine bestimmte Art des Nüsseknackens nicht voneinander ab, sondern jedes Tier entwickelt sie von selbst, so dass jedes Individuum sozusagen das Rad neu erfindet. Der Unterschied der fehlenden Nachahmung dürfte ein wesentlicher Grund dafür sein, dass Schimpansen-Gesellschaften sich so stark von der menschlichen unterscheiden, schreibt F. Jötten in ZEIT ONLINE.
„Kratzen am Mythos der unvoreingenommenen Wissenschaft“
Über die verhaltensbiologischen Ergebnisse der Studien von Tennie hinaus thematisiert ZEIT ONLINE-Autor F. Jötten den Wissenschaftsbetrieb. Der Ausgang der Experimente war – gemessen an den Erwartungen und der bislang dominierenden Auffassung – negativ. Negative Ergebnisse seien aber sehr viel schwerer in Fachzeitschriften zu veröffentlichen als positive. Außerdem würden negative Ergebnisse der scientific community viel kritischer auf methodische Fehler abgeklopft. Jötten zitiert Eric-Jan Wagenmakers, Professor für Methodische Psychologie an der Universität Amsterdam, dass negative Ergebnisse oft nicht zur Publikation gelangten. Vielleicht seien deshalb „95 Prozent aller publizierten Studienergebnisse in der Psychologie, zu der auch Tennies Forschungsgebiet zählt, positiv“. Wenn das so ist, repräsentiere die wissenschaftliche Literatur nicht die Wirklichkeit.
Dies gelte offenbar auch für die Schimpansenforschung. „Wir lieben es, Ähnlichkeiten zwischen Menschenaffen und Menschen zu finden“ wird Brian Hare, Professor für evolutionäre Anthropologie an der Duke University in North Carolina zitiert. Ähnlichkeiten zwischen Schimpansen und Menschen würden womöglich überbetont. Bei Tennie sei es wohl deshalb besser gelaufen, weil sein Doktorvater einer der renommiertesten Forscher in der evolutionären Anthropologie ist und weil das frei zugängliche, angesehene Internetjournal PLOS ONE negativen Resultaten eine gleichberechtigte Chance auf Veröffentlichung ermögliche.
Dass Wissenschaftler interessengeleitet arbeiten und entsprechende Fragestellungen entwickeln, ist selbstverständlich. Die Orientierung auf erwartete Ergebnisse ist jedoch der Wahrheitsfindung abträglich. Sich dessen bewusst zu sein könnte helfen, Ergebnisse besser einzuordnen und nicht überzubewerten. Frederik Jötten in ZEIT ONLINE wird deutlich: Die Arbeiten von Tennie „kratzen auch am Mythos der unvoreingenommenen Wissenschaft und erwecken den Verdacht, dass sie vor allem das zur Kenntnis nimmt, was zu den herrschenden Moden passt.“ Es ist freilich nicht „die Wissenschaft“, sondern es sind die Wissenschaftler. Gut aber, dass Korrekturen möglich sind, wenn Publikationsmöglichkeiten nicht von sachfremden Interessen behindert werden.
Quellen
Tennie C, Call J & Tomasello M (2012) Untrained Chimpanzees (Pan troglodytes schweinfurthii) Fail to Imitate Novel Actions. PLoS ONE 7(8): e41548. doi:10.1371/journal.pone.0041548.
Jötten F (2013) Von wegen menschlich. ZEIT ONLINE, 4.3.2013; http://www.zeit.de/2013/09/Wissenschaft-Schimpansen-Menschen (sehr lesenswert).
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n194.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
18.12.12 100 Jahre Piltdown-Mensch
Vor 100 Jahren wurde der Piltdown-Mensch entdeckt, eine der berühmtesten Fälschungen der Wissenschaftsgeschichte, die erst 41 Jahre später entlarvt wurde. Die Hintergründe der Fälschung sind noch immer nicht vollständig aufgeklärt. Unabhängig davon sind die Erkenntnisse, die dieser Fall über die Rolle des Wissenschaftlers in Wissenschaftsbetrieb bringt, nachdenkenswert.
Am 18. Dezember 1912 wurde der wissenschaftlichen Öffentlichkeit ein spektakulärer Fossilfund eines Vormenschen vorgestellt. Arthur Smith Woodward, ein britischer Paläontologe, und Charles Dawson, ein Amateur-Altertumsforscher hatten in einer Kiesgrube bei dem Dorf Piltdown in Südostengland einen fossilen Schädel entdeckt, der ein menschliches Schädeldach und einen affenartigen Unterkiefer besaß. Damals waren noch relativ wenige Funde fossiler Menschen und fossiler Menschenaffen bekannt und der „Piltdown-Mensch“ unterstützte die damals vorherrschende Auffassung, dass sich in der Evolution des Menschen zuerst das Gehirn vergrößert habe, bevor der aufrechte Gang entstand. Der Piltdown-Mensch schien ein passendes Bindeglied zwischen Menschenaffen und Menschen zu sein. Zu Ehren des Entdeckers erhielt die neue Art den wissenschaftlichen Namen Eoanthropus dawsoni – „Morgenrötemensch“.
Über den Piltdown-Menschen wurden hunderte von Publikationen veröffentlicht und er beeinflusste die Einschätzung der Bedeutung anderer Funde, die später gemacht wurden (vor allem der Gattung Australopithecus, deren erste Art 1924 entdeckt wurde). Eine Minderheit der Experten bezweifelte allerdings, dass Schädeldach und Unterkiefer wirklich zusammengehören. Schließlich wurde der Fund im Jahr 1953 aufgrund chemischer Analysen als Fälschung entlarvt. Der Schädel war aus einem Schädeldach eines Menschen und einem Unterkiefer eines Orang-Utan zusammengesetzt und die Knochen waren so bearbeitet worden, dass nicht sofort auffallen konnte, dass sie von verschiedenen Schädeln stammen. Außerdem wurden nach der Entlarvung als Fälschung Spuren der mechanischen Bearbeitung entdeckt. Diese wären von Anfang an erkennbar gewesen (so Gould [1989] in einem seiner vielen lesenswerten Essays); man war aber offenbar nicht darauf gefasst – das ist das eigentlich Interessante an der Geschichte (s. u.).
Erstaunlicherweise sind die Hintergründe der Fälschung nach wie vor nicht aufgeklärt, berichtet Chris Stringer in der neuesten Ausgabe von Nature (Stringer 2012; vgl. Gould 1989; 1991). Er gehört zu einem Team von Forschern, die herausfinden wollen, wie die Fälschung durchgeführt wurde, wer sie durchführte und welche Motivation dahinterstand. Gould (1989) schildert einige Hypothesen über den oder die Täter. Es ist demnach durchaus möglich, dass es sich um einen Scherz und nicht um eine böswillige Fälschung handelt und dass dieser Scherz durch besondere Umstände nicht mehr aufgeklärt wurde.
Doch wer immer die Piltdown-Fälschung zu verantworten hat und was auch immer seine Motive waren, ist der Fall – so Stringer –, eine Warnung an Wissenschaftler, eine kritische Haltung zu bewahren, aber auch ein Beispiel für die Leistungsfähigkeit der naturwissenschaftlichen Methode (es waren chemische Untersuchungen, die zeigten, dass Schädel und Unterkiefer sicher nicht zusammenpassen). Wenn etwas zu gut zu sein scheint, um wahr zu sein, dann ist es vielleicht auch so, merkt Stringer (2012, 179) an.
Doch in den Jahren nach 1912 hielten sich die meisten mit der Sache befassten Wissenschaftler nicht an solche Warnungen. Gould (1989, 120) nennt vier Arten von Gründen, dass eine „so unausgegorene Sache von den größten englischen Paläontologen ohne Vorbehalt akzeptiert wurde. Alle vier widersprechen dem üblichen Glauben, dass die wissenschaftliche Praxis es mit ‚harten‘, grundlegenden Fakten zu tun habe …“ (Hervorhebung nicht im Original). Die vier Gründe, die Gould nennt, sind: 1. „Die Gründung großer Hoffnungen auf zweifelhafte Beweismittel“, 2. „Verringerung einer Abweichung von der Norm durch ihre Anpassung an kulturelle Vorurteile“ (was dazu führt, dass wahrgenommen wird, was man erwartet, und das Unerwartete übersieht), 3. „Verringerung der Abweichungen von der Norm durch die Anpassung von Tatsachen an Erwartungen“ (man sieht Dinge, die es gar nicht gibt, s. u.), 4. „Schutz vor Entdeckung von Seiten der Praktiker“. Gould (1989, 120) stellt fest, dass diese Gründe unter Beweis stellten, „daß die Wissenschaft eine durchaus menschliche Tätigkeit ist, welche durch Hoffnungen, kulturelle Vorurteile und das Streben nach Ruhm motiviert wird und dennoch auf ihren Irrwegen zu einem besseren Verständnis der Natur gelangt.“ Die naturwissenschaftlichen Daten erreichen uns „stets durch die starken Filter unserer Kultur, ihrer Hoffnungen und Erwartungen“ (Gould 1989, 122). Obwohl das Schädeldach „bemerkenswert modern“ war, entdeckten Wissenschaftler daran eine Reihe von „definitiv affenartigen Merkmalen“ (vgl. den 3. Punkt oben)!
Nicht gerechtfertigt wäre es, den Fall Piltdown als Beispiel für ein verbreitetes Vorkommen von Fälschungen in der Wissenschaft zu werten; dafür gibt es jedenfalls keine Beweise. Es sind mittlerweile sehr viele fossile Funde von Menschen und Menschenaffen gemacht worden, die als Grundlage für die Einschätzung von Pro und Contra einer Evolution des Menschen aus dem Tierreich dienen können. Gerade Funde und Untersuchungen der jüngeren Zeit lassen eine deutliche Kluft zwischen fossilen Menschenaffen und Menschen erkennen (s. z. B. Brandt 2012; Hartwig-Scherer 2011 und die in diesen Artikeln verarbeitete Originalliteratur).
Literatur
Brandt (2012) „Homo“ habilis war kein Mensch. Kluft zwischen fossilen Menschen und Menschenaffen größer geworden. Stud. Int. J. 19, 4-11.
Gould SJ (1989) Ein Wiedersehen mit Piltdown. In: Der Daumen des Panda. Frankfurt, S. 112-128. (amerikan. Original 1980)
Gould SJ (1991) Die Piltdown-Verschwörung. In: Wie das Zebra zu seinen Streifen kam. Frankfurt, S. 199-224.
Hartwig-Scherer S (2011) Ardipithecus – ein Astgänger sägt am Lehrbuchwissen. Stud. Int. J. 18, 68-77.
Stringer C (2012) The 100-year mystery of Piltdown Man. Nature 492, 177-179.
Autor dieser News: Reinhard Junker
© 2012, http://www.genesisnet.info/schoepfung_evolution/n190.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
10.05.10 Neuer Australopithecus-Fund – Bindeglied zum Menschen?
Erneut wurde ein interessantes Fossil aus der Gruppe der Australopithecinen gefunden. 2008 entdeckten Forscher um Lee Berger und Peter Schmid in Südafrika zwei Teilskelette, die neben zahlreichen Australopithecus-Merkmalen auch einige menschenähnliche Anpassungen aufweisen, die bislang von keinem anderen Australopithecinen bekannt sind. Deshalb wurde eine neue Art etabliert: Australopithecus sediba. Trotz dieser interessanten Merkmalsmischung wird Australopithecus sediba selbst nicht als ein Bindeglied zum Menschen vorgeschlagen.
Die Merkmale von Australopithecus sediba erlauben keine genaue phylogenetische (= stammesgeschichtliche) Positionsbestimmung zu den frühen Homo-Arten, und das radiometrische Alter ist für ein tatsächliches Bindeglied um einiges zu gering (1,78 bis 1,95 Millionen Jahre). Die in ihrer taxonomischen Zuordnung zur Gattung Homo weiterhin umstrittenen Formen rudolfensis und habilis treten zeitlich früher auf als der neue Fund und auch der erste unbestrittene Vertreter unserer eigenen Gattung – Homo erectus – lebte zeitgleich mit der neuen Art. Die morphologische Kluft zwischen den beiden letzteren ist beträchtlich. Der Ursprung von Homo erectus bleibt weiterhin im Dunkeln (vgl. Australopithecinen („Südaffen“) und andere Menschenaffenartigen-Fossilien, https://www.genesisnet.info/index.php?Artikel=43622&Sprache=de&l=1 und Stufen des Menschen?, https://www.genesisnet.info/index.php?Artikel=43623&Sprache=de&l=1).
Der Fund. Ein Forscher-Team um Lee Berger entdeckte 2008 zwei Teilskelette von Homininen in der Malapa-Höhle, einem Fundplatz in Südafrika 15 km nordnordöstlich so berühmter Fundplätze wie Sterkfontein, Swartkrans und Kromdraai. (Hominine sind Menschenähnliche; damit werden alle fossilen und lebenden Menschenformen einschließlich ihrer im Evolutionsmodell vermuteten Vorläufer bezeichnet.)
Berger und seine Kollegen, u. a. der Züricher Anthropologe Peter Schmid beschrieben die Funde kürzlich in Science (Berger et al. 2010). Es handelt sich um zwei Teilskelette, ein jugendliches, vermutlich männliches Individuum (MH1) und ein erwachsenes, vermutlich weibliches Individuum (MH2). Die Forscher konnten die Funde aufgrund ihrer Merkmale keinem bekannten fossilen Homininen zuordnen und gaben ihnen deshalb einen neuen Namen: Australopithecus sediba.
Australopithecus heißt „Südaffe“ und sediba in der Bantu-Sprache Sesotho „Brunnen“ oder „Quelle“. Die Australopithecinen wurden im Verlauf ihrer Entdeckungsgeschichte Anfang des letzten Jahrhunderts zunächst als besondere Affen, genauer als Großaffen angesehen. Mitte des vorigen Jahrhunderts änderte sich dies und man interpretierte sie von da an als frühe Hominiden, Wesen, die im evolutionären Stammbaum nicht mehr Großaffe und noch nicht Mensch und damit „Vormenschen“ oder „Affenmenschen“ waren.
Berger et al. (2010) diskutieren die Beziehung von Australopithecus sediba zu den ersten Menschen. Aufgrund mosaikartiger Merkmalsverteilungen kommen sie zum Schluss, dass die phylogenetische Position der neuen Australopithecus-Art zu den frühen Homo-Arten nicht genau zu bestimmen ist. Außerdem diskutieren sie chronologische Fragen einer solchen Ahnenschaft. Nur wenn man annimmt, dass Australopithecus sediba schon wesentlich früher gelebt hat als derzeit fossil nachgewiesen, dann ist solch eine Linie zeitlich möglich (Berger et al. 2010).
Doch zunächst zu den Merkmalen von Australopithecus sediba.
Nach Berger et al. (2010) besitzt Australopithecus sediba die größte Ähnlichkeit mit Australopithecus africanus. Beide weisen zahlreiche gemeinsame Merkmale am Hirnschädel, am Gesichtsschädel, am Unterkiefer und an den Zähnen auf. Das Besondere an Australopithecus sediba ist die aber Tatsache, dass diese neue Art unter allen Australopithecinen die größte Ähnlichkeit zum Menschen (Homo) aufweist.
Die Ausführungen der beiden folgenden Abschnitte müssen etwas detaillierter und fachspezifischer ausfallen. Bei Anklicken der farblich markierten Begriffe wird eine Erklärung sichtbar.
Ähnlichkeit mit den Menschen. Der Hirnschädel ist transversal* relativ breit und seine Scheitelbeinwände sind vertikal ausgerichtet. Dem Gesicht fehlen die hervortretenden ausladenden Jochbeine von Australopithecus africanus. Die Ausbildung der supraorbitalen Tori*, der nasoalveolaren Region*, der infraorbitalen Region* und der Jochbeine gibt dem Gesicht von Australopithecus sediba ein fortgeschritteneres Aussehen gegenüber Australopithecus africanus.
*Begriffe:
- transversal (= senkrecht zur Körperlängsachse)
- supraorbitalen Tori (= Knochenwülste auf dem Stirnbein oberhalb der Augenhöhle)
- nasoalveolare Region (= Gebiet von Nase und Zahnfächer)
- infraorbitale Region (= unterhalb der Augen)
Die Symphyse des Unterkiefers, also die Stelle, an der die beiden Hälften in der Mittellinie zusammengewachsen sind, ist vertikal ausgerichtet und mit einem leichten Kinn versehen. Auch an den Zähnen sind einige Merkmale vorhanden, die Australopithecus sediba von Australopithecus africanus unterscheiden und mit Homo verbinden.
Gemeinsamkeiten mit anderen Australopithecinen. Australopithecus sediba besitzt aber auch viele Merkmale gemeinsam mit Australopithecus. Dazu gehören am Schädel eine kleine Gehirnkapazität (420 cm3), eine hervortretende Glabellaregion*, ein mäßiges Eckzahnjugum* mit Fossa canina*, eine kleine Spina nasalis anterior*, ein steil geneigter zygomaticoalveolarer* Knochenkamm und ein hoher Ursprung des Musculus masseter*. Außer an den Zähnen gibt es am Körperskelett von Australopithecus sediba eine große Anzahl an Merkmalen, die auch bei anderen Australopithecinen zu finden sind. Dazu gehören neben einer geringen Körpergröße (130 cm) relativ lange Arme mit großen Gelenkflächen, Schulterblätter mit nach oberen ausgerichteter Gelenkfläche für den Oberarmknochen, ein deutlicher Knochenkamm am unteren Teil des Oberarmknochens für die brachioradialen* Muskeln, eine große und tiefe Fossa olecranii*, die eine Öffnung mit Scheidewand aufweist, lange robuste gekrümmte Phalangen (Fingerknochen) mit starken Ansätzen für die Sehnen des Musculus flexor digitorum superficialis (Fingerbeugemuskeln) und viele weitere Anpassungen an ein Leben in Bäumen.
*Begriffe:
- Glabellaregion (= Region zwischen den Augenbrauen)
- Eckzahnjugum (= Vorwölbung des Eckzahnfaches)
- Fossa canina (= paarige Knochengrube des Oberkiefers)
- Spina nasalis anterior (= unpaariger Knochenfortsatz; am weitesten vorne gelegene Knochenstruktur des Oberkiefers)
- zygomaticoalveolar (= Jochbein und Zahnfächer zugehörig)
- Musculus masseter (= Kaumuskel).
- brachioradial (= Oberarm und Speiche zugehörig)
- Fossa olecranii (= Auf der Rückseite des Oberarmknochens befindliche Vertiefung, die in der Streckstellung des Unterarms einen kräftigen Knochenvorsprung der Elle (Olecranon) aufnimmt)
Allerdings gibt es auch einige Merkmale am Beckenskelett von Australopithecus sediba, die Homo ähneln und bei allen anderen Australopithecinen nicht vorhanden sind. Dazu gehören eine stärkere Verstrebung und eine vergrößerte hintere Region des Darmbeins, ein verringerter Abstand zwischen dem Kreuzbein-Darmbeingelenk und Hüftgelenk sowie zwischen der Hüftgelenkpfanne und der Rauigkeit am Sitzbein.
Deutungen. Berger et al. deuten diese Merkmale im Zusammenhang mit einer besseren zweibeinigen Fortbewegungsweise von Australopithecus sediba. In diesem Zusammenhang ist es jedoch merkwürdig, dass der Bau des Fußes ähnlich der der anderen Australopithecinen von Homo deutlich verschieden ist. So besitzt das Sprungbein (Talus) eine flache Trochlea* auf, deren innerer und äußerer Rand einen gleichen Krümmungsradius aufweist. Der Talushals ist kurz und stämmig. Er ist medial verdreht und weist einen großen horizontalen und einen niedrigen Torsionswinkel auf. Auch das Fersenbein (Calcaneus) ist in der Gesamtmorphologie deutlich von Homo verschieden. Der Knochen in Relation zur Längsachse ist stark abgewinkelt, wobei der maximale Wendepunkt sich an der großen peronealen Trochlea* befindet. Am Fersenbein fehlt ein Knochenhöcker am äußeren unteren Rand. Die Achse des Fersenbeins ist zur transversalen Ebene im Winkel von 45 Grad ausgerichtet. Die Gelenkfläche zwischen dem Fersenbein und dem Würfelbein (Os cuboideum) ist vertikal ausgerichtet und es fehlt ein erweiterter hinterer Vorsprung für die Zuspitzung der Nase des Würfelbeins. Diese und weitere Merkmale sprechen für eine von Homo verschiedene weniger geschickte Bipedie*.
*Begriffe:
- flache Trochlea (= Knochenrolle auf der Oberseite des Sprungbeins)
- große peroneale Trochlea (= Knochenrolle auf der Außenseite des Fersenbeines)
- Bipedie (= Zweibeinigkeit)
Da der Gesamtbauplan von Australopithecus sediba dem der Australopithecinen ähnelt, ist die Zuordnung der beiden Teilskelette von Berger et al. (2010) in die Gattung Australopithecus gut begründet.
Welche Stellung nimmt Australopithecus sediba im phylogenetischen Konzept ein?
Von wem stammt Australopithecus sediba ab? Wohin führt Australopithecus sediba? Ist es ein Bindeglied zu unserer eigenen Gattung Homo?
Berger et al. (2010) plädieren wegen der Ähnlichkeit mit Australopithecus africanus dafür, dass Australopithecus sediba von dieser Australopithecus-Art abstammt. Bisher konnte man aber den Schichten, aus denen die Überreste von Australopithecus africanus stammen, kein sicheres Alter zuschreiben. Schätzungen gehen aber von ca. 2,5 bis 3,5 Millionen Jahren aus (Sawyer & Deak 2008).
Zur Beantwortung der Frage, in welcher Beziehung Australopithecus sediba zu unserer eigenen Gattung steht, muss erst geklärt werden, welche fossilen Formen zu Homo gehören.
In der paläanthropologischen Literatur werden neben Homo erectus häufig auch die Arten habilis und rudolfensis als früheste Vertreter von Homo genannt. Diese Klassifikation ist aber aus guten Gründen umstritten.
Wood und Collard (1999) haben habilis und rudolfensis wegen zahlreicher Australopithecus-ähnlicher Merkmale aus Homo ausgegliedert und zu den Australopithecinen gestellt. Danach haben M. Leakey et al. (2001) für rudolfensis eine erneute Reklassifizierung vorgenommen und wegen Ähnlichkeiten mit dem neu entdeckten Kenyanthropus platyops ihn auch in diese neue Gattung gestellt. Eine zwischenzeitliche Neurekonstruktion von rudolfensis (KNM-ER 1470) mit einem etwas geringeren Schädelvolumen und stärker vorspringenden Mittelgesicht (Bromage et al. 2008) rechtfertigt diese Reklassifizierung nachträglich. Von den frühen Homo-Formen kann gesichert nur Homo erectus als „erster echter Mensch“ klassifiziert werden.
Der auf 1,78 bis 1,95 Millionen Jahre datierte Australopithecus sediba kann nun aber kein Bindeglied zu Homo erectus sein, denn letzterer tritt praktisch zeitgleich vor 1,9 Millionen Jahren auf (Wood 1991, Gabunia & Vekua 1995). Vor allem ist die Kluft im Körperbau deutlich zu groß. Der größte Unterschied dürfte aber im kognitiven Bereich liegen.
Aber auch eine Weiterentwicklung zu habilis und rudolfensis, die von Berger et al. (2010) morphologisch näher an Homo erectus stehend eingestuft werden, ist nicht plausibel. Denn rudolfensis und habilis (H. aff.) sind mit 2,4 Millionen (Schrenk et al. 1993) bzw. 2,3 Millionen Jahren (Kimbel et al. 1997) deutlich älter als Australopithecus sediba. Auch das Merkmalsmosaik von Australopithecus sediba spricht nicht unbedingt für solch eine Ahnenschaft. Wenn aber, wie Berger et al. (2010) nicht ausschließen, Australopithecus sediba deutlich früher aus Australopithecus africanus evolvierte, dann ist eine Linie zu rudolfensis und habilis zeitlich möglich. Es stellt sich aber die Frage nach der phylogenetischen Plausibilität (Selektionsdrücke?) einer Entstehung von Australopithecus sediba aus Australopithecus africanus und anschließender langen Koexistenz beider Spezies auf engem Raum.
Der üblicherweise als Ahne von Homo erectus angenommene habilis weist neben einem großen Unterschied in der Anatomie, insbesondere des Körperskeletts (Johanson et al. 1987, Hartwig-Scherer & Martin 1991, Brandt 1995), auch eine große zeitliche Überlappung mit Homo erectus auf und ist als Ahne deshalb wenig plausibel. Der Ursprung des frühesten echten Menschen Homo erectus ist ungeklärt.
Aiello und Collard (2001, S. 527) schreiben: „Zumindest vorerst ist es wahrscheinlich am besten, die Benennung von Vorfahren zu vermeiden und es bei einer einfachen Unterteilung zwischen den Vorfahren des Menschen mit archaischen Merkmalen (Orrorin, Ardipithecus, Australopithecus – einschließlich Paranthropus – und Kenyanthropus) und den Menschen und seine Vorfahren mit modernen Merkmalen (Homo sapiens und die übrigen Arten von Homo) zu belassen.“
Und Asfaw et al. (2002, S. 319) stellen fest: „Die Ursprünge der weitverbreiteten, vielgestaltigen frühen pleistozänen Homo erectus-Linie bleibt unklar. Die deutlichen Unterschiede zwischen irgendwelchen potentiellen Ahnen (Homo habilis oder andere) und dem frühesten bekannten H. erectus könnte ein plötzliches evolutionäres Ereignis signalisieren …“ Diese Einschätzungen haben bezüglich der Abgrenzung von Homo erectus und nichtmenschlichen Formen auch unter Berücksichtigung neuerer Entdeckung wie Australopithecus sediba nichts an Aktualität eingebüßt.
Die Verwandtschaftsverhältnisse der fossilen Homininen mit ihren mosaikartigen Merkmalsverteilungen und der großen Kluft zwischen den echten Menschen und allen anderen Homininen können besser in einem Grundtypmodell als in einem phylogenetischen Modell verstanden werden. In diesem Modell repräsentieren die Australomorphen („Vormenschen“) einschließlich habilis und rudolfensis einen oder mehrere großaffenähnliche Grundtypen, während die echten Menschen einen davon unabhängigen Grundtyp bilden.
Literatur
Aiello LC & M Collard (2001) Our newest oldest ancestor? Nature 410, 526-527.
Asfaw B, Gilbert WH, Beyene Y, Hart WK, Renne PR, WoldeGabriel G, Vrba ES & TD White (2002) Remains of Homo erectus from Bouri, Middle Awash, Ethiopia. Nature 416, 317-320.
Berger LR, de Ruiter DJ, Churchill SE, Schmid P, Carlson KJ, Dirks PHGM & JM Kibii (2010) Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science 328, 195-204.
Brandt M (1995) Der Ursprung des aufrechten Ganges. Neuhausen-Stuttgart.
Bromage TG, McMahon JM, Thackeray JF, Kullmer O, Hogg R, Rosenberger AL, Schrenk F & DH Enlow (2008) Craniofacial architectural constraints and their importance for reconstructing the early Homo skull KNM-ER 1470. The Journal of Clinical Pediatric Dentistry 33, 43-54.
Gabunia L & A Vekua (1995) A Plio-Pleistocene hominid from Dmanisi, East Georgia, Caucasus. Nature 373, 509-512.
Hartwig-Scherer S & RD Martin (1991) Was “Lucy” more human than her “child”? Observations on early hominid postcranial skeletons. Journal of Human Evolution 21, 439-449.
Johanson DC, Masao FT, Eck GG, White TD, Walter RC, Kimbel WH, Asfaw B, Manega P, Ndessokia P & G Suwa (1987) New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania. Nature 327, 205-209.
Kimbel WH, Johanson DC & Y Rak (1997) Systematic assessment of a maxilla of Homo from Hadar, Ethiopia. American Journal of Physical Anthropology 103, 235-262.
Leakey MG, Spoor F, Brown FH, Gathogo PN, Kiarie C, Leakey LN & I McDougall (2001) New hominin genu from eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433-440.
Sawyer GJ & V Deak (2008) Der lange Weg zum Menschen. Heidelberg.
Spoor F, Leakey MG, Gathogo PN, Brown FH, Antón SC, McDougall I, Kiarie C, Manthi FK & LN Leakey (2007) Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya. Nature 448, 688-691.
Schrenk F, Bromage TG, Betzler C, Ring U & YM Juwayeyi (1993) Oldest Homo and Pliocene biogeography of the Malawi Rift. Nature 365, 833-836.
Wood B (1991) Koobi Fora research project. Vol. 4. Oxford.
Wood B & M Collard (1999) The human genus. Science 284, 65-71.
Autor dieser News: Michael Brandt
© 2010, http://www.genesisnet.info/schoepfung_evolution/n149.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
21.11.09 Evolution der Religion
Wenn der Mensch durch Evolution aus dem Tierreich entstanden ist, müssen seine Gottesvorstellungen ein Produkt der Evolution sein. Schon Darwin mutmaßte, dass Gottesglaube entstanden sei, weil er sozial nützlich sei. Das ist bis heute eine der Spuren, die verfolgt werden, um eine Evolution der Religion plausibel machen zu können. In einem Überblicksartikel der Zeitschrift Science werden verschiedene weitere Hypothesen vorgestellt, die jedoch kaum prüfbar und ziemlich vage sind und bestenfalls Begleiterscheinungen von Religiosität darstellen, aber nicht als nachgewiesene Ursachen gelten können.
Wie kann ein bloßer Naturprozess Vorstellungen von einer Realität hervorbringen, die es gar nicht gibt? Mit dieser Frage sind atheistische Evolutionstheoretiker herausgefordert, die den Ursprung des Gottesglaubens aus evolutionären Prozessen ableiten wollen. Wenn Evolution naturalistisch verstanden wird, sollte dieser Prozess also die Vorstellung einer Illusion hervorgebracht haben.
Die Idee ist so alt wie Darwins frühe Überlegungen zur Evolution der Lebewesen. Die beiden unübertroffenen Darwin-Biographen Adrian Desmond & James Moore schreiben über seine Gedanken zur evolutionären Entstehung des Gottesglaubens: „In der Abstammungsfrage konzentrierte sich sein Interesse jetzt auf den Menschen. … Nach seiner Auffassung erklärte die Evolution jede geistige Eigenheit, jede körperliche Haltung, nicht nur Wirbelsäule und Milz, sondern auch die Lebensweise des Menschen, seine Instinkte, seine Gedanken, Gefühle, sein Gewissen und seine Moral. Jeder Trieb, jedes Verlangen … seien … doch alle evolutionäre Erbgüter – selbst die Anbetung Gottes. ‚Liebe zur Gottheit [ist] Folge von [geistiger] Organisation, o du Materialist!’ flüsterte er sich zu“ (S. 286). „Auch die christlichen Gebote ‚Tu anderen wie dir selbst’ und ‚Liebe deinen Nächsten wie dich selbst’ hätten sich ganz natürlich aus den ‚sexuellen, elterlichen und sozialen Instinkten’ unserer Vorfahren entwickelt“ (S. 300). „Unsere ‚angeborene Kenntnis des Schöpfers’ habe sich infolge ‚seiner erhabensten Gesetze’ entwickelt. Sie sei ein großartiger Instinkt, der wegen seiner sozialen Nützlichkeit entstanden sei“ (S. 308; kursiv jeweils im Original, Darwin wörtlich wiedergebend).
Wenn man den heutigen Stand der Forschung zugrundelegt, wie ihn Elizabeth Culotta in einem Überblicksbeitrag Anfang November 2009 in der Wissenschaftszeitschrift Science darstellt, könnte Darwin einige Hellsichtigkeit bescheinigt werden. Dass die „Entstehung von Religion“ auf das Verhalten evolutionärer affenartiger Vorstufen zurückgeführt wird, ist selbstverständlich, sobald eine allgemeine Evolution der Lebewesen als Tatsache akzeptiert wird. Denn Evolution macht vor den Verhaltensweisen nicht halt, auch sie tragen zur Fitness bei. Und wenn es verbreitet Gottesglaube gibt, muss er – evolutionstheoretisch interpretiert – einen Nutzen haben, der das Überleben fördert. Für Evolutionstheoretiker stellt sich daher die Frage, worin der Überlebensvorteil besteht und wie er durch natürliche Prozesse entstehen konnte.
In ihrem Überblicksartikel stellt Culotta zunächst fest, dass es unter Wissenschaftlern zu dieser Frage keinen Konsens gebe. Einige meinen, Religion würde kooperatives Verhalten fördern, was die Fitness steigere, und sei daher entstanden – Darwin lässt grüßen (s. o.). Andere sehen die Wurzeln in unserem differenzierten sozialen Erkenntnisvermögen; wir hätten die Tendenz, Zeichen von „Agenten“ in unserer Welt zu sehen, die wie wir in der Welt wirksam sind. Die Wurzeln von Religiosität werden also in unserem Gehirn gesucht. Wir seien hypersensitiv für Zeichen von „Agenten“, die wie wir selber denken; wir neigen dazu, natürliche Dinge auf Agenten zurückzuführen („Hypotertropie sozialer Kognition“). Das habe Theismus gefördert, meint die Wahrnehmungspsychologin Deborah Kelemen von der Boston University.
Andere Forscher gehen die Frage nach dem Ursprung der Religion von archäologischer Seite her an und suchen nach Anzeichen alter religiöser Vorstellungen. Eine dabei verfolgte Spur ist soziales „symbolisches Verhalten“, Religion sei eine Komponente solchen Verhaltens. Der Gebrauch von Symbolen sei ein Schlüssel für aufkeimende Spiritualität.
Solche Hinweise sind jedoch äußerst vage und bestenfalls Begleiterscheinungen, aber keine nachweislichen Ursachen für die Entstehung von Religion. Das gilt auch für Bestattungen oder das Anfertigen von religiösen Fruchtbarkeitsobjekten oder Löwenmenschen. Alle diese Aktivitäten erklären nicht das Woher der Religion.
Besonders hingewiesen wird auf teleologisches Denken bei Kindern. Kinder neigen besonders dazu, natürlichen Phänomenen Zwecke zuzuschreiben statt sie auf bloße natürliche Ursachen zurückzuführen. Man könne bei Kindern annehmen, dass diese Tendenz angeboren sei. Es sei schwer, diese teleologischen Erklärungen zu überwinden. Solche zweckorientierten Erklärungsweisen seien ein Schritt auf dem Weg zur Religion. Man könne daran erkennen, dass ein Gott eine naheliegende Konstruktion für den menschlichen Geist sei. Es sei ein kleiner Schritt, vom Design auf einen Designer zu schließen.
Abgesehen davon, dass auch diese Erklärung für die Entstehung des Gottesglaubens hochgradig spekulativ ist, kann das teleologische Denken leicht ohne jeden religiösen Bezug erklärt werden: Es entspricht ganz einfach ständiger menschlicher Erfahrung, dass es zielgerichtete Handlungen gibt. Spaemann und Löw nennen es in ihrem Klassiker „Die Frage wozu?“ eine primäre Welterfahrung des Menschen. Teleologisches Denken ist erfahrungsgestützt. Dass Menschen auch an Agenten denken, auch wenn es sich um einen natürlichen Vorgang handeln könnte (z. B. bei einem plötzlichen Knacken im Geäst), ist ebenfalls Folge von Erfahrung: Vieles von dem, was wir erleben, wird durch handelnde Personen verursacht und im Zweifel muss diese Möglichkeit zunächst bedacht werden. Von dieser Art des Umgangs mit potentiellen Agenten auf die Entstehung eines Gottesglaubens zu schließen, überstrapaziert die Befunde bei weitem.
Culotta (2009) zitiert auch kritische Stimmen. Demnach seien die Modelle weit davon entfernt, die komplexen Systeme von Gottheiten und Ritualen zu erklären, die Religion ausmachen, selbst wenn mehr Daten vorliegen werden. Und die Erklärung, dass religiöses Verhalten durch Anpassung erklärt werden könne, weil es Solidarität innerhalb der Gruppe fördere, sei fraglich, so Pascal Boyer von der Washington University in St. Louis, da der Nachweis fehle, dass die Menschen den religiösen Forderungen auch nachkommen würden.
Quellen
Culotta E (2009) On the origin of religion. Science 326, 784-787.
Desmond S & Moore J (1991) Darwin. München.
Spaemann R & Löw R (1981) Die Frage Wozu? Geschichte und Wiederentdeckung des teleologischen Denkens. München.
Autor dieser News: Reinhard Junker
© 2009, http://www.genesisnet.info/schoepfung_evolution/n138.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.07.09 Neue Texte zum Alter der Menschheit
Nach evolutionstheoretischen Vorstellungen beginnt die Geschichte der Menschheit bereits vor mindestens zwei Millionen Jahren. Diese Ansicht basiert hauptsächlich auf radiometrischen Datierungen. Doch es gibt andere Daten, die sehr deutlich auf ein sehr viel geringes Alter hinweisen, ja sogar nur dann schlüssig verstanden werden können, wenn die Menschheit nur höchstens etwa 10.000 Jahre alt ist. In zwei neuen Texten werden diese Argumente vorgestellt und die wichtigsten Gegenargumente überblicksartig diskutiert.
Aus demographischer Sicht (Bevölkerungsentwicklung) wirft ein hohes Alter der Menschheit eine folgende ungelöste Frage auf: Wie kommt es, dass die Erdbevölkerung über fast die ganze Zeit ihrer Geschichte fast nicht gewachsen ist? Im Artikel Daten zum Bevölkerungswachstum, https://www.genesisnet.info/index.php?Artikel=43661&Sprache=de&l=1 wird gezeigt, dass die bekannten Daten zur Bevölkerungsentwicklung im Widerspruch zu einem Menschheitsalter von etwa zwei Millionen Jahren stehen. Das aus der biblischen Urgeschichte resultierende geringe Menschheitsalter von maximal 10.000 Jahren passt zu den Daten über die Bevölkerungsentwicklung viel besser. Diese Schlussfolgerung kann gegen alle Kritikpunkte erfolgreich verteidigt werden.
Im zweiten Artikel Steinwerkzeugmengen und Besiedlungsspuren, https://www.genesisnet.info/index.php?Artikel=43662&Sprache=de&l=1 geht es um die Fundmengen von Steinwerkzeugen. Auch diese stehen nicht im Einklang mit einem hohen Menschheitsalter. Das gilt auch für Erkenntnisse, die man aus anderen Besiedlungsspuren gewinnen kann. Wenn die Altsteinzeit wirklich zwei Millionen Jahre lang gedauert hätte, müssten die umherstreifenden Wildbeuter eine gewaltige Menge archäologisch auffindbarer Überreste hinterlassen haben. Nach intensiver Forschung kann die Anzahl der produzierten Steinwerkzeuge verlässlich abgeschätzt werden. Es zeigt sich, dass viel zu wenige vorhanden sind und dass ihre Anzahl viel besser zu einem Menschheitsalter von nur maximal etwa 10.000 Jahren passt.
Genauere und detaillierte Begründungen für die in diesen beiden Artikeln zusammengestellten Argumente und weiterführende Informationen bietet das Buch „Wie alt ist die Menschheit?“ von Michael Brandt (https://www.wort-und-wissen.org/produkt/vergessene-archaeologie/), das jüngst in dritter Auflage erschienen ist.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2009, http://www.genesisnet.info/schoepfung_evolution/n133.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
03.06.09 Ein Fund der „alles ändert“?
Was hat „Tante Ida“ mit dem Menschen zu tun?
Als „sensationeller Fund, nach dem die Wissenschaft von der Entstehung des Menschen neu überdacht werden muss“, als langgesuchtes Bindeglied, als „Meilenstein“, gar als „achtes Weltwunder“1 wurde ein Fossil bezeichnet, das kürzlich der Öffentlichkeit vorgestellt wurde.2 Darwinius masillae, auf 47 Millionen Jahre datiert und etwas eingängiger auf „Ida“ getauft, machte in den letzten Wochen enorm viel von sich reden. Das Fossil schmückte sogar einen Tag lang das Google-Logo. Es handelt sich um ein weibliches Jungtier, das vom Kopf bis zur Spitze des langen Schwanzes 58 Zentimeter misst und wegen seines schlanken Körperbaus und dem langen Schwanz den Lemuren ähnelt. Lemuren sind Halbaffen, die auf Madagaskar leben. Ida fehlen aber die charakteristischen vorstehenden Schneidezähne, die als Kamm der Fellpflege dienen, und die „Putzkralle“ am zweiten Zeh. Das ist ein besonders gestalteter Zeigefinger, der typisch für diese Halbaffen ist. Andere Merkmale weisen in Richtung der Entwicklung des Menschen, so der Sprunggelenksknochen. Ein großer, den übrigen Fingern gegenüberstehender Daumen weist darauf hin, dass das Tier gut greifen und klettern konnte und vermutlich auf Bäumen lebte.3

Abb. 1: Fossil von Darwinius masillae, alias „Ida“. (Wikimedia: Von Jens L. Franzen, Philip D. Gingerich, Jörg Habersetzer1, Jørn H. Hurum, Wighart von Koenigswald, B. Holly Smith – http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0005723;jsessionid=E8154D7406947B36A39470C790A4F08C, CC BY-SA 2.5)
Der Enthusiasmus ist in einer Hinsicht auf jeden Fall berechtigt: Es handelt sich um ein außerordentlich gut erhaltenes Fossil und um den am besten erhaltenen Primaten, der jemals als Fossil gefunden wurde. Neben 95 % des Skeletts sind auch Körperumrisse und der Mageninhalt erhalten. Wirklich fantastisch.4 Dabei schlummerte das kostbare Stück ein Vierteljahrhundert lang bei einem Privatsammler in der Schublade, ehe es von Wissenschaftlern einer Forschergruppe um Jörn Hurum von der Universität Oslo gekauft und zwei Jahre lang untersucht wurde. Entdeckt wurde es in der berühmten Fossillagerstätte Messel, einer Ölschiefergrube bei Darmstadt. Aber wegen der hervorragenden Erhaltung alleine hätte man die Veröffentlichung dieses Fundes sicher nicht in diesem Ausmaß in der Presse inszeniert. Viel wichtiger ist die Einschätzung als „der bei weitem älteste Vorfahre von uns Menschen“ (ZDF, Terra X).
Kritik. Doch die wissenschaftliche Diskussion über „Ida“ hat gerade erst begonnen. Jens L. Franzen, einer der Bearbeiter, gab in einem Radiobeitrag zu Protokoll, dass es noch Diskussionen und verschiedene Meinungen zur Einordnung des Fossils geben werde. Das Wissenschaftsmagazin Science zitierte in einem Internetartikel den Paläontologen Elwyn Simons: „Es ist ein außerordentlich gut erhaltenes, wundervolles Exemplar, aber es erzählt uns nicht viel mehr, als wir schon wussten.“5 Manche Forscher tendieren dazu, das Tier als einen indirekten Verwandten des Menschen zu interpretieren. Nach Franzen wäre „Ida“ zwar „nicht unsere Ur-Ur-Ur-Großmutter, aber vielleicht unsere Ur-Ur-Ur-Großtante“.6
Chris Beard vom Carnegie Museum of Natural History sieht den Fund noch kritischer7: Ida sei kein missing link, jedenfalls nicht zwischen menschenähnlichen (anthropoiden) Formen und primitiveren Primaten. Weitere Studien müssten zeigen, ob es zwischen anderen Mitgliedern der Adapiformes aus dem Eozän vermittelt, aber das würde kaum den Status als achtes Weltwunder rechtfertigen. Ähnlich kritisch äußert sich auch Callum Ross, Paläontologe der University of Chicago.8
Die Adapiformes bilden eine Linie, die nach evolutionstheoretischen Vorstellungen zu den heutigen Lemuren führt. Idas Merkmale passen gut zu dieser Gruppe.9 Um in einer Verbindung zu anthropoiden Formen stehen zu können, müsste Ida entsprechende Merkmale besitzen; hierin scheitere Ida „kläglich“. Im Stammbaum nimmt das Fossil daher eine Position ein, die deutlich von der Linie abgesetzt ist, die zum Menschen führen soll.10
Kritisch angemerkt wird auch, dass Hurum und sein Team nur 30 bis 40 statt der üblichen 200 bis 400 Merkmale des Fossils mit denen anderer Funde verglichen hätten; außerdem fehlten wichtige Arten in diesem Vergleich. Die gut ins Bild passenden Rosinen seien herausgepickt worden, Hinweise auf Fossilienanalysen der vergangenen Jahre würden fehlen.11 Außerdem seien in der Originalarbeit die letzten 15 Jahre der Forschung nicht berücksichtigt worden.
In einem neueren kritischen Science-Artikel merkt auch Gibbons an, dass viele führende Wissenschaftler, die sich mit der Evolution der Primaten befassen, Ida nicht als Vorfahren des Menschen, sondern als Verwandte der Lemuren ansehen; das Skelett liefere wenig neue Information über alte Primaten, noch weniger über den Ursprung des Menschen. Einige befürchten, dass der Pressewirbel um Ida nach hinten losgehen werde, wenn die wahre Identität von Ida offenbar würde und dass Glaubwürdigkeit verloren gehe.12
Vermarktung. Die Veröffentlichung des Fossils wurde ungewöhnlich reißerisch vermarktet. Seltsam mutet an, dass nach Angaben der New York Times in den USA die Entdeckung von Ida in TV-Clips mit dem Slogan „This changes everything“ angekündigt wurde.13 Wie kann dieser Fund „alles ändern“, wenn er angeblich die Evolution des Menschen bestätigt, die doch schon längst bewiesen sein soll? Der Bearbeiter Hurum verteidigte den Aufwand mit den Worten: „Jede Popgruppe tut dasselbe. Jeder Sportler macht das. Wir müssen beginnen, in der Wissenschaft auch so zu denken.“14 Angesichts der tatsächlichen Bedeutung des Fossils kann man sich nicht des Eindrucks erwehren, dass das breite Publikum bei jeder sich bietenden Gelegenheit auf Evolution eingeschworen werden soll. In der Originalarbeit15 ist jedenfalls von „missing link“ oder „revolutionär“ nirgends die Rede. Der Kontrast zwischen der marktschreierischen Präsentation in der Öffentlichkeit und der tatsächlichen Bedeutung des Fossils gemäß den Einschätzungen der Wissenschaftler ist nicht zu übersehen.
Anmerkungen
1 „Für mich ist dieses Fossil das achte Weltwunder, so was hat die Welt noch nicht gesehen“, sagt Dr. Jens Franzen, ein führender Paläontologe und ausgewiesener Spezialist für Fossilien vom Senckenbergmuseum Frankfurt. (ZDF, Terra X)
2 „Uraffen-Fossil löst medialen Wirbel aus!“ (http://science.orf.at/science/news/155791): „Mehrere Fernsehsender griffen zu und warfen eine gewaltige Werbemaschinerie an.“
3 Franzen JL, Gingerich PD, Habersetzer J, Hurum JH, von Koenigswald W, et al. (2009) Complete Primate Skeleton from the Middle Eocene of Messel in Germany: Morphology and Paleobiology. PLoS ONE 4(5): e5723. doi:10.1371/journal.pone.0005723
4 Viele Bilder gibt es im frei zugänglichen Originalartikel (s. Anm. 3), oder auch unter der Internetadresse von Anm. 11
6 http://www.welt.de/die-welt/article3771505/Wissenschaft.html
8 Gibbons A (2009) „Revolutionary“ Fossil Fails to Dazzle Paleontologists. ScienceNOW Daily News, 19 May 2009, http://sciencenow.sciencemag.org/cgi/content/full/2009/519/1
9 „Like all adapiforms, Ida lacked a ‚toothcomb’ at the front of her lower jaw – a structure that living lemurs use for grooming fur. Ida also lacked a ‚grooming claw’ on her second toe, another difference from living lemurs. Otherwise, Ida’s overall proportions and anatomy resemble that of a lemur, and the same is true for other adapiform primates.“
10 siehe Abbildung unter http://www.newscientist.com/data/images/ns/cms/dn17173/dn17173-1_500.jpg
{kind=link}
11 Gibbons (2009) (s. Anm. 8); vgl. http://www.br-online.de/bayern2/iq-wissenschaft-und-forschung/fossil-vorfahre-mensch-ID1242807873161.xml?_requestid=747968
12 Gibbons A (2009) Celebrity Fossil Primate: Missing Link or Weak Link? Science 324, 1124-1125.
13 http://www.stern.de/wissenschaft/mensch/:Uraffen-Fossil-Ida-Kritik-Sensationsfund/701371.html
14 s. Anm. 13; in Science (Anm. 12) wird Hurum wie folgt zitiert: „It’s hard to discuss haplorrhine and strepsirrhines in a press release. You need to link it with us.“
15 siehe Anm. 3
Autor dieser News: Reinhard Junker
© 2009, http://www.genesisnet.info/schoepfung_evolution/n130.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.05.08 Neuer Artikel „Stufen des Menschen?“
Die Fossilgeschichte des Menschen wird in „Evolution – ein kritisches Lehrbuch“ (siehe https://www.wort-und-wissen.org/publikationen/lehrbuch/) relativ ausführlich behandelt. Auf dessen Grundlage war bereits voriges Jahr eine Überblicks-Artikel über die Australopithecus-Artigen in Genesisnet veröffentlicht worden („Australopithecinen („Südaffen“) und andere Menschenaffenartigen-Fossilien“, https://www.genesisnet.info/index.php?Artikel=43622&Sprache=de&l=1). In dieser Gruppe werden evolutionstheoretisch meist die Vorfahren des Menschen gesucht. Mittlerweile liegt mit dem Artikel „Stufen des Menschen?“, https://www.genesisnet.info/index.php?Artikel=43623&Sprache=de&l=1 auch eine Zusammenfassung über die Gattung Homo (Mensch) vor. In die Gattung Homo werden nur Formen gestellt, bei denen es sich eindeutig um Menschen handelt. Evolutionstheoretisch ist zu erwarten, dass es unterschiedliche Stufen des Menschseins gibt und dass dies durch Fossilfunde dokumentiert wird. Es sind zwar tatsächlich verschiedene Menschenformen fossil bekannt, aber zweifelsfreie Nachweise für primitive Menschen-Vorstufen gibt es nicht. Unterschiede erklären sich wahrscheinlich teilweise durch verschiedene Umweltbedingungen und sind Ausdruck der Polyvalenz (Vielgestaltigkeit) des Grundtyps Mensch (zur Polyvalenz siehe Artikel „Genetisch polyvalente Stammformen von Grundtypen“, https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1). Der Mensch ist nur als geistig-kulturell voll entwickeltes Geschöpf bekannt. Die wichtigsten Argumente sind im Artikel „Stufen des Menschen?“, https://www.genesisnet.info/index.php?Artikel=43623&Sprache=de&l=1 zusammengestellt. Zur genaueren Argumentation wird auf das oben genannte kritische Lehrbuch sowie auf Monographien der Reihe „Studium Integrale“ und entsprechende Artikel in „Studium Integrale Journal“ verwiesen.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2008, http://www.genesisnet.info/schoepfung_evolution/n113.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
06.03.08 Wissenschaft oder Paläophantasie?
Unterschiedliche Ansichten zur Entwicklung des menschlichen Gehirns
Der bekannte Evolutionsbiologe Richard Lewontin (Harvard University) hat auf der Jahrestagung der AAAS (American Association for the Advancement of Science; 14.-18. 2. 2008 in Boston) das Auditorium mit seinen Ausführungen unter dem Titel „Warum wir nichts über die Entwicklung der kognitiven Fähigkeiten wissen“ herausgefordert und provoziert. Unter anderem führte Lewontin aus: Uns fehlt der Fossilbericht über die kognitiven (= Erkenntnis und Denken betreffend) Fähigkeiten des Menschen, deshalb erfinden wir Geschichten. Aus seiner Sicht erlaubt die Interpretation der Fossilien keine zuverlässige Rekonstruktion der Entwicklung des menschlichen Gehirns. Außerdem warnt er davor, von der Größe fossiler hominider (= menschenartiger) Gehirnkapseln (Cranium) auf die kognitiven Fähigkeiten der Lebewesen zu schließen.
Diese ernüchternden Einschätzungen blieben auf der Tagung nicht unwidersprochen und so führt Balter (2008) in seinem Bericht von der Tagung in der Wissenschaftszeitschrift Science weitere Sprecher an, die sich zu diesem Thema äußerten und kontrovers darüber diskutierten.
Der Anthropologe Dean Falk (Florida State University, Tallahassee) führte Ergebnisse aus der vergleichenden Psychologie, Genetik, Neuroimaging und Paläoanthropologie an, die Beiträge zur Evolution menschlichen Erkenntnisvermögens leisten.
Christopher Welsh, Genetiker (Harvard Medical School) geht davon aus, dass Einsichten in die genetische Steuerung der Gehirnentwicklung beim modernen Menschen dazu beitragen können, die Entwicklungsgeschichte des menschlichen Gehirns besser zu verstehen. So könnten Veränderungen am Gen ASPM, das bei der embryonalen Entwicklung von Nervenzellen beteiligt ist, aufgrund entsprechender Selektionsprozesse einen Mechanismus zur dramatischen Vergrößerung des Cortex im Verlauf der Evolution der Hominiden liefern. Aus der Sicht der New Yorker Anthropologin Leslie Aiello stellen Fossilbericht und archäologische Befunde gute Hinweise auf die Hauptphasen menschlicher Evolution bereit. Diese dokumentierten die Trennung der Entwicklungslinien zwischen Schimpanse und Mensch oder die Erfindung von Steinwerkzeugen vor ca. 2,5 Millionen Jahren. Außerdem machte sie darauf aufmerksam, dass Modelle über Paläoklimata evolutionäre Ereignisse mit Veränderungen in der Umwelt in einen sinnvollen Zusammenhang bringen.
Marc Hauser (Psychologe, Harvard University) wies hin auf den Unterschied in der kognitiven Leistungsfähigkeit zwischen Mensch und anderen „smart species“, womit er Tiere wie Schimpansen, Elefanten und Delphine bezeichnete. Aus seiner Sicht ist dieser Unterschied größer als derjenige zwischen diesen Tieren und Würmern. Er vergleicht die zweifellos oft erstaunlichen Fähigkeiten von Tieren z. B. beim Lösen von spezifischen Problemstellungen mit einer sog. „Laserstrahl-Intelligenz“ und stellt sie der menschlichen „Flutlicht-Intelligenz“ gegenüber. Menschen können nach Hauser Problemlösungen im Gegensatz zu Tieren auch auf viele neue Problemstellungen übertragen; der Werkzeuggebrauch bei Tieren unterscheide sich deutlich von entsprechenden Handlungen von Menschen. Hauser erhofft sich von den Untersuchungen solcher Unterschiede wichtige Hinweise auf die Evolution unserer Art.
Als Titel seines Berichts über diese Vorträge und Diskussionen bei der diesjährigen AAAS-Jahrestagung verwendet Balter (2008) eine interessante Formulierung: „How Human Intelligence evolved – Is it Science or Paleofantasy?“ Mit dem Begriff „Paläophantasie“ nimmt er einen Begriff auf, der von Leslie Aiello in einer Reaktion auf Lewontin verwendet worden ist. Letzterer ist bereits in der Vergangenheit durch unorthodoxe Äußerungen und Ideen – z.T. gemeinsam mit Stephen Jay Gould – aufgefallen. Richard Lewontin hat mit seinen provokativen Thesen den Wissenschaften einen wichtigen Dienst erwiesen, indem er auf die Gefahr hingewiesen hat, dass Interpretationen von Daten übertrieben und gesicherter dargestellt und auch übernommen werden als sie sind. Jedenfalls schafft dieser Bericht von den kontroversen Diskussionen auf der AAAS-Tagung Freiraum für verschiedene Denkansätze und weitere innovative Impulse. Darin hebt er sich wohltuend ab von Darstellungen, wie die von Morell & Musi (2008) in der jüngsten Ausgabe von National Geographic Deutschland, in welcher der Eindruck erweckt wird, dass der Unterschied in der kognitiven Leistungsfähigkeit zwischen Menschen und Tieren nach neueren wissenschaftlichen Erkenntnissen zunehmend marginalisiert (= an den Rand gedrängt, unbedeutend gemacht) sei. Gerade beim Versuch, Ursprungsfragen zu klären, wird es wichtig bleiben, der Versuchung, Paläophantasien zu etablieren, zu widerstehen und Wissenschaft zu treiben – auch indem man deren Grenzen (be-)achtet.
Literatur
Balter M (2008) How human intelligence evolved – is it science or paleofantasy? Science 319, 1028.
Morell V & Musi VJ (2008) Können Tiere denken? National Geographic Deutschland 3/2008, S. 44-69.
Autor dieser News: Harald Binder
© 2008, http://www.genesisnet.info/schoepfung_evolution/n107.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
25.10.07 Mensch und Schimpanse noch weiter auseinander
Vergleichende Genomik: zunehmend größerer Unterschied zwischen Mensch und Schimpanse
Der genetische Unterschied zwischen Mensch und Schimpanse erweist sich durch weitere Untersuchungen entgegen ursprünglichen Annahmen als zunehmend größer. Die häufig zitierte Differenz zwischen den beiden Primatengenomen von ca. 1 % wird mehr und mehr durch neue Befunde in Frage gestellt. In einem Newsbeitrag hatten wir bereits darüber berichtet (Über den genetischen Unterschied zwischen Mensch und Schimpanse – der „1 %-Mythos“, https://www.genesisnet.info/index.php?News=87). In einer Analyse der Genome von Rhesusaffe (Macaca mulatta), Hund (Canis familiaris), Ratte (Rattus norvegicus), Maus (Mus musculus), Schimpanse (Pan troglodytes) und Mensch (Homo sapiens) untersuchten Hahn et al. (2007) Datensätze mit 119.746 Genen aus 9.900 Genfamilien aus allen 6 Arten. Alle Genome sind nach dem sogenannten „Schrotschuss-Verfahren“ (nach C. Venter) mindestens 6-fach sequenziert und mindestens zu 96 % untersucht worden. Hahn und Mitarbeiter zeigen, dass (unter evolutionstheoretischen Voraussetzungen) die Geschwindigkeit von Genverlusten und -gewinnen in der Linie der Primaten zunimmt, besonders bei den Großaffen. In verschiedenen Genfamilien ist die Änderung der Kopienzahl von Genen auffällig groß, die Autoren sehen darin den Einfluss natürlicher Selektion. Die Autoren folgern aus den Untersuchungen, dass im Genom des Menschen (seit seiner vermuteten Abspaltung von der Schimpansenlinie vor 5-6 Millionen Jahren) 678 Gene dazugekommen sind, während im Schimpansen-Genom* 740 Gene verloren wurden. Dies bedeutet, dass sich für 6,4 % (1.418 von 22.000) aller menschlichen Gene keine direkt vergleichbaren (orthologen) Gegenstücke im Schimpansen-Genom* finden lassen. Es bleibt spannend zu verfolgen, ob sich dieser Trend im genetischen Unterschied mit wachsenden Kenntnissen fortsetzt. Viel bedeutsamer ist jedoch, die Zusammenhänge und Bedeutungen der im Genom niedergelegten Information zu verstehen. Die großen Unterschiede implizieren zudem ernsthafte Fragen nach den Mechanismen der Veränderungen und auch nach deren Triebfeder.
*Genom = komplettes Erbgut
Quellen:
Hahn MW, Demuth JP & Han S-G (2007) Accelerated rate of gene gain and loss in primates. Genetics (published ahead of print, published on 18. October, 2007 as 10.1534/genetics.107.080077)
Dolgin E (2007) Evolutionary sprint made us human. (23.10.2007; http://sciencenow.sciencemag.org/cgi/content/full/2007/1023/2)
Autor dieser News: Harald Binder
© 2007, http://www.genesisnet.info/schoepfung_evolution/n98.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
04.07.07 Über den genetischen Unterschied zwischen Mensch und Schimpanse – der „1 %-Mythos“
Die Flut von DNA-Sequenzdaten nimmt zu und die Vielzahl von Publikationen über genetische Analysen ist unüberschaubar. Diese Entwicklung darf jedoch nicht darüber hinwegtäuschen, dass wir uns erst im Vorfeld eines Verständnisses von der Organisation genetischer Information und deren Bedeutung für einen Organismus befinden. Das wurde nicht zuletzt durch die jüngste Veröffentlichung des ENCODE1-Project-Consortiums deutlich. Darin legen Mitarbeiter aus 35 kooperierenden Arbeitsgruppen die bisher umfangreichsten Untersuchungen von ca. 30 Mb (Megabasen, Millionen Basen) des menschlichen Genoms (= Ergbuts) vor (dies entspricht etwa 1 % des gesamten Erbguts). Diese Resultate eröffnen neue Einsichten, u. a.: Das menschliche Genom wird fast durchgängig transkribiert, d.h. die allermeisten Basen werden in RNA umgeschrieben; „nicht codierende“ Abschnitte wurden identifiziert, die entgegen bisherigen Annahmen ebenfalls transkribiert werden. Für zahlreiche Sequenzabschnitte, die bisher als funktionslos angesehen wurden, konnten regulatorische Eigenschaften aufgezeigt werden. Mit anderen Worten: Der seit Beginn der Sequenzierungsaktivitäten häufig verwendete Begriff „junk-DNA“ („DNA-Müll“, womit man einen erheblichen Anteil z. B. des menschlichen Genoms bezeichnete, der ohne Funktion zu sein schien) war voreilig und kann als Hinweis auf unsere Unwissenheit aufgefasst werden. Damit ist auch ein häufig angeführtes Argument gegen „Intelligent Design“ hinfällig geworden, wonach funktionsloses Design gegen Schöpfung spreche. Die ausgesprochen populäre Evolutionsdeutung von unverstandener DNA als „evolutionärer Müll“ erweist sich zunehmend als irreführend. Glücklicherweise hat sich die empirische biologische Forschung von dieser evolutionären Spekulation nicht beeindrucken lassen und ist dabei, den Irrtum aufzuklären.
1 % Unterschied zwischen Mensch und Schimpanse? Ähnlich wie der Begriff vom „DNA-Müll“ hat sich auch die vermeintliche Tatsache von der weitgehenden Übereinstimmung zwischen dem Genom des Menschen (Homo sapiens) und des Schimpansen (Pan troglodytes) im Allgemeinwissen eingenistet; der genetische Unterschied soll gerade mal 1 % betragen.
Die Behauptung der ca. 99 %-igen Übereinstimmung der Genome von Mensch und Schimpanse wurde erstmals von Wilson & King (1975) in einem Science-Artikel aufgestellt. Damals erregte dieser erstaunlich geringe Unterschied Aufsehen. Bei aufmerksamer Lektüre der Veröffentlichung von Wilson & King fällt jedoch auf, dass die beiden Autoren vermuten, neben den Genen (bekannte codierende Sequenzabschnitte) müssen weitergehende Differenzen vorliegen, die ursächlich für die auffälligen Unterschiede z. B. in Anatomie und Verhalten sind. Sie dachten dabei vor allem an genregulatorische Prozesse.
Nachfolgende Untersuchungen unterstützten die weitgehende genetische Übereinstimmung der beiden Primaten. Als 2005 vom Chimpanzee Sequencing and Analysis Consortium die Sequenz des Schimpansengenoms veröffentlicht wurde (King war im Autorenteam), bestätigten diese Untersuchungen die auffällige Übereinstimmung: der Vergleich von 2,4•109 Basen der beiden Arten ergab eine Differenz von 1,23 %. In der Publikation wurde jedoch ebenfalls vermerkt, dass diese Angabe ausschließlich den Austausch einzelner Basen berücksichtigt, nicht jedoch die vielen größeren Genomabschnitte, die eingefügt (Insertion) bzw. herausgeschnitten (Deletion) worden sind. Diese Bereiche werden häufig als „indels“ bezeichnet. Berücksichtigt man diese „indels“, so ergibt sich nach Berechnungen des Schimpansen-Konsortiums eine Differenz von zusätzlich 3 %. Weitere vergleichende Untersuchungen zwischen dem menschlichen Erbgut und dem Schimpansengenom relativierten die Angabe von ca. 1 % Unterschied zunehmend und veranlassten Cohen (2007), in einem Science-Artikel vom „Mythos“ dieses 1 %-igen Unterschieds zu sprechen.
Doch nicht nur „indels“ vergrößern den Unterschied zwischen den beiden Genomen. Auch ganze Gene werden dupliziert oder werden eliminiert. Diese Vorgänge verursachen nach Berechnungen von Hahn und Mitarbeitern (2006) einen Unterschied von 6,4 %. Nach Ansicht der Autoren spielten Genduplikationen (= Gen-Verdoppelungen) und Genverluste bei der Entwicklung von spezifisch menschlichen Phänotypen (= äußere Gestalt) eine größere Rolle als der Austausch von Basen und ganz sicher eine größere Rolle, als man bisher geschätzt hat.
Geschwind hat gemeinsam mit Mitarbeitern untersucht, welche von 4000 Genen in bestimmten Arealen des Gehirns gleichzeitig angeschaltet (exprimiert) werden und aus den gewonnenen Daten ein Gen-Netzwerk für verschiedene Organismen konstruiert. Die Autoren berichten, dass im Cortex beim Menschen 17,4 % der Vernetzungen spezifisch sind und beim Schimpansen nicht auftreten (Geschwind et al. 2006).
Abschließend zitiert Cohen Svante Pääbo vom Max-Planck-Institut für Evolutionäre Anthropologie in Leipzig; dieser antwortete auf die Frage, ob man unter Berücksichtigung aller bekannten Daten den Unterscheid zwischen Mensch und Schimpanse genau beziffern könne, folgendermaßen: Er glaube nicht, dass es eine Möglichkeit gebe, eine Zahl zu berechnen. Weiter wird Pääbo mit der Aussage zitiert: „In the end, it’s a political and social and cultural thing about how we see our differences.“ Dem ist eigentlich nichts hinzuzufügen außer vielleicht der Wunsch, dass sich diese Einsicht ähnlich rasch und nachhaltig in das Allgemeinwissen eingräbt wie die noch viel zu häufig und unreflektiert erwähnten 1 %-Unterschied zwischen Mensch und Schimpanse. Wir wissen inzwischen mehr!
1 ENCODE: ENCyclopedia Of DNA Elements
Literatur
Cohen J (2007) Relative differences: the myth of 1 %. Science 316, 1836.
Demuth JP, Bie TD, Stajich JE, Christianini N & Hahn MW (2006) Evolution of mammalian gene families. PLoS ONE 1(1): e85, doi:10.1371.
King MC & Wilson AC (1975) Evolution at two levels in human and chimpanzees. Science 188, 107-116.
Oldham MC, Horwath S & Geschwind DH (2006) Conservation and evolution of gene coexpression networks inhuman and chimpanzee brains. Proc. Natl. Acad. Sci. USA 103, 17973-17978.
The Chimpanzee Sequencing and Analysis Consortium (2005) Initial sequence of the chimpanzee genome and comparison with the human genome. Nature 437, 69-87.
The ENCODE Project Consortium (2007) Identification and analysis of functional elements in 1 % of the human genome by the ENCODE pilot project. Nature 447, 799-816.
Autor dieser News: Harald Binder
© 2007, http://www.genesisnet.info/schoepfung_evolution/n87.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
18.06.07 Neue Texte über „Urmenschen“ und „Urvögel“
Wichtige Indizien zur Erhellung der Herkunft des Menschen sind zweifellos Fossilien, das sind konservierte Überreste von früheren Lebewesen in Form von Versteinerungen, Abdrücken u. a. (vgl. Was ist Paläontologie?, https://www.genesisnet.info/index.php?Artikel=42821&Sprache=de&l=1). Von Menschen und Menschenaffen sind zahlreiche Fossilien bekannt. Indizien für eine Vorfahrenstellung zum Menschen können vor allem der Fortbewegung, der Gehirngröße und -struktur, dem Gebiss und der Fähigkeit zur Werkzeugherstellung bzw. ihren Entsprechungen im Körperbau entnommen werden.
Die ausgestorbene Menschenaffengattung Australopithecus und einige ähnliche Gattungen (als „Australomorphe“ zusammengefasst) werden evolutionstheoretisch als mögliche Vorfahren des Menschen diskutiert. Alle diese Formen weisen jedoch Merkmale auf, die zu einer Übergangsstellung nicht passen. Das gilt auch für die erst in jüngerer Zeit entdeckten Gattungen Orrorin, Kenyanthropus und Sahelanthropus. Die bekannten Fossilien lassen sich nicht recht in eine widerspruchsarme Linie zum Menschen einordnen; bei jeder Art kommen Merkmale vor, die den jeweiligen Stammbaumdarstellungen widersprechen. Dies wird im neuen Artikel „Australopithecinen („Südaffen“) und andere Menschenaffenartigen-Fossilien“ (https://www.genesisnet.info/index.php?Artikel=43622&Sprache=de&l=1) von Sigrid Hartwig-Scherer gezeigt. In diesem Artikel werden die wichtigsten Formen der Australomorphen und andere ausgestorbene Menschenaffen vorgestellt, die als mögliche Vorfahren des Menschen diskutiert werden.
In populären Darstellungen werden die Australomorphen häufig als „Urmenschen“ bezeichnet, doch diese Bezeichnung ist nicht gerechtfertigt. Die Bezeichnung „Urmensch“ ist nicht durch die Merkmale dieser Gruppe begründet, sondern resultiert aus der verbreiteten Auffassung, dass aus diesen Formen der Mensch evolutiv hervorgegangen sei.
Die Vielfalt der Australomorphen lässt sich alternativ im Sinne eines eigenen, sehr vielfältigen Grundtyps deuten (vgl. zur Grundtypenbiologie „Heutige Grundtypen“, https://www.genesisnet.info/index.php?Artikel=1241&Sprache=de&l=1). Der Grundtyp besitzt ein Repertoire an Merkmals-Ausprägungsmöglichkeiten, das sich durch Ausbreitung und Spezialisierungen in verschiedene Richtungen „sternförmig“ entfaltet (Mikroevolution; Radiation; vgl. „Genetisch polyvalente Stammformen von Grundtypen“, https://www.genesisnet.info/index.php?Artikel=1244&Sprache=de&l=1). Beim Versuch, Stammbäume mit diesen vielen Formen zu entwerfen, äußert sich das in Merkmalswidersprüchen. Das heißt: Je nach Gewichtung der verschiedenen Merkmale ergeben sich unterschiedliche Stammbaum-Rekonstruktionen, und wenn man alle gleich gewichtet, resultiert daraus eine busch- oder sternförmige Darstellung.
„Urvögel“. Die heutigen Vögel sind gegenüber anderen heute lebenden Wirbeltieren deutlich abgegrenzt. Lange Zeit vermittelte fast allein der berühmte fossile „Urvogel“ Archaeopteryx aus dem Oberjura ein Bild davon, über welche Station ein evolutionärer Übergang von Reptilien zu Vögeln verlaufen sein könnte, denn er weist eine ausgeprägte Kombination von reptilienartigen und vogeltypischen Merkmalen auf. Der evolutive Übergang von Reptilien zu den Vögeln gilt nach der Entdeckung zahlreicher Fossilien in den letzten Jahren mittlerweile als gut belegt. Zu Archaeopteryx gesellen sich weitere fossile Gattungen (vor allem kleine Raubdinosaurier), die in den Übergangsbereich Reptilien – Vögel gestellt werden. Dennoch bleibt unklar, wie das besondere Vogelmerkmal der Federn und wie der Flug entstanden sind. Aufgrund von Merkmalswidersprüchen ist auch der evolutive Übergang zu den Vögeln umstritten.
Im neuen Artikel „Entstehung der Vögel“ (https://www.genesisnet.info/index.php?Artikel=42844&Sprache=de&l=1) von Reinhard Junker werden der „Urvogel“ Archaeopteryx und die Theropoden (zweibeinige Raubdinosaurier) als mögliche Vorfahren der Vögel vorgestellt und kritisch diskutiert. Außerdem werden Theorien zur Entstehung der Federn und des Vogelflugs vorgestellt und das Für und Wider erläutert. Folgende Ergebnisse werden präsentiert:
- Unter den zweibeinigen theropoden Dinosauriern der Kreide gibt es zahlreiche Mosaikformen mit unterschiedlichen Vogel- und Reptilmerkmalen. Daher gelten sie als die besten Kandidaten für Vogelvorfahren.
- Die zunehmende Vogelartigkeit einiger Formen unterstützt die evolutionstheoretische Deutung. Insgesamt sind die Merkmale bei den betreffenden Gattungen jedoch so mosaikartig verteilt, dass vielfach Konvergenzen und Reversionen (Rückentwicklungen) angenommen werden müssen, auch bei manchen Schlüsselmerkmalen.
- Die ältesten fossilen Federn erscheinen in fertiger Form bei Archaeopteryx. Andere Fossilerhaltungen feder- oder haarartiger Strukturen sind deutlich jünger und in ihrer Deutung umstritten. Bei manchen kreidezeitlichen Formen wird Flugverlust als wahrscheinlich betrachtet.
- Umstritten ist, welche Selektionsdrücke den Erwerb von Federn und der Flugfähigkeit begünstigt haben könnten.
- Sowohl in der Kreide als auch zu Beginn des Tertiärs treten zahlreiche Vogelgruppen plötzlich und mit markanten Diskontinuitäten auf.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2007, http://www.genesisnet.info/schoepfung_evolution/n85.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
15.05.06 Alter der Menschheit: 2 Millionen oder 10.000 Jahre?
In der Kontroverse um die Evolutionstheorie ist die Zeitproblematik von grundlegender Bedeutung. Die Existenz großer Zeiträume ist eine notwendige Voraussetzung für Makroevolution. In dem kürzlich erschienenen Wort und Wissen-Buch „Wie alt ist die Menschheit?“ diskutiert Michael Brandt eine Reihe von Befunden, die völlig rätselhaft erscheinen, wenn die Menschheit (mindestens) zwei Millionen Jahre alt ist (was durch radiometrische Datierungen begründet wird). Anhand des Bevölkerungswachstums und der gefundenen Steinwerkzeugmengen zeigt der Autor, dass selbst ein Alter von 10.000 Jahren für die Menschheit eher noch zu hoch gegriffen ist. Kritik an dieser Schlussfolgerung wird ausführlich behandelt und entkräftet.
Die mittlerweile vorliegenden Befunde zur Geschwindigkeit des Bevölkerungswachstums und zur Menge an hinterlassenen Steinwerkzeugen werden von Brandt erstmals überblicksmäßig dargestellt. Seinen Ausführungen liegen ein umfassendes Literaturstudium und Informationen von Werkzeugsammlern zugrunde. Dass die Daten einer Millionen Jahre währenden Menschheitsgeschichte widersprechen, wird in der Fachpublikationen durchaus gesehen; es werden jedoch keine Konsequenzen daraus gezogen. Dies holt Michael Brandt in seinem Buch nach: Die zahlreichen Anomalien, die bei Zugrundelegung eines großen Alters der Menschheit auftreten, können widerspruchsfrei und schlüssig erklärt werden, wenn die Menschheitsgeschichte auf maximal 10.000 Jahre verkürzt wird.
Die Brisanz dieses Buches kann kaum überschätzt werden. Denn die vorgelegten Daten widersprechen der evolutiven Deutung der Menschheitsgeschichte massiv: Sie entziehen nämlich einem wesentlichen Teil der Evolutionsgeschichte des Menschen eine notwendige Basis: die dafür benötigen großen Zeiträume. Gleichzeitig passen die Befunde zu einem Zeitrahmen, der durch die Überlieferung der biblischen Urgeschichte nahegelegt wird.
Der Autor beweist mit diesem Buch nebenbei, dass das Hinterfragen etablierter Lehrmeinungen zu Wissenszuwachs und neuen Erkenntnissen führen kann. Wer dagegen die Evolution des Menschen als nicht mehr hinterfragbare Tatsache wertet, hätte Studien dieser Art niemals durchgeführt und wäre daher auch nicht auf die in diesem Buch vorgelegten Ergebnisse gestoßen. Einmal mehr erweist sich Deutungsvielfalt – namentlich in historischen Fragen – als Motor für die Gewinnung neuer Erkenntnisse.
Das Buch ist trotz seiner wissenschaftlichen Tiefe auch für Laien gut lesbar. Es richtet sich daher an alle an der Vorgeschichte der Menschheit Interessierten, Fachleute wie Laien. Der Leser erhält über das eigentliche Thema hinaus auch viele interessante Einblicke in die Lebensumstände der Menschen der Steinzeit.
Weitere Infos zum Buch und Link zum Bestellen: https://www.wort-und-wissen.org/produkt-kategorie/si/. Die Einleitung und das komplette Inhaltsverzeichnis finden sich unter https://www.wort-und-wissen.org/produkt/wie-alt-ist-die-menschheit/.
Autor dieser News: Reinhard Junker
© 2006, http://www.genesisnet.info/schoepfung_evolution/n65.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
14.02.05 Filmbesprechung: „Der Fall des Affenmenschen“
Der Filmemacher Fritz Poppenberg hat einen neuen evolutionskritischen Film, diesmal zur Evolution des Menschen, vorgelegt. Das ist ungewöhnlich in einer Medienlandschaft, welche Evolutionskritiker – sofern diese überhaupt beachtet werden – auf bemerkenswert unsachliche, kenntnisarme und professionelle Weise mit Spott und Häme überzieht. Im Bonus-Material der DVD wird dieser traurige Sachverhalt anhand eines entsprechenden ZEIT-Artikels belegt. Es ist mutig und anerkennenswert, dass sich Fritz Poppenberg mit seinem Film derart pointiert außerhalb der politisch korrekten Meinung unserer Medienwelt stellt. Als Selbständiger weiß er genau, dass damit auch ein Risiko verbunden ist – schon deshalb wünsche ich dem Werk auch wirtschaftlichen Erfolg.
Aus dem Film geht das persönliche Engagement des Regisseurs hervor – er nimmt den Zuschauer mit hinein in seine eigene Entdeckungsreise. Der Film beginnt mit einem spannenden Bericht über die nahezu vergessenen Experimente sowjetischer Wissenschaftler, die unter dem Eindruck der Darwinschen Evolutionslehre Kreuzungen zwischen Schimpansen und Menschen erzeugen wollten. Ausführlich wird berichtet, mit welchen Schwierigkeiten das Filmteam bei seinen Recherchen in Afrika konfrontiert wurde und es wird deutlich, dass solche Versuche aus biologischen Gründen zum Scheitern verurteilt sind: Menschen und afrikanische Großaffen sind in ihren Grundbauplänen zu unterschiedlich.
Im zweiten Teil stellt der Film wichtige paläontologische Funde zur Fossilgeschichte des Menschen vor. Es handelt sich um einen Hauptteil des Werkes, er ist insgesamt hervorragend gelungen. Man darf Poppenberg gratulieren, er hat komplizierte Sachverhalte durch sehr gut geschnittene Interviews, Schädelaufnahmen und Filmsequenzen spannend und klar präsentiert. Der programmatische Untertitel des Werkes „Die Evolutionstheorie kann die Herkunft des Menschen nicht erklären“ wird hier überzeugend anhand paläontologischer Daten erläutert. Dieser Teil hätte m.E. etwas ausführlicher ausfallen dürfen. Die Grafik der Hirnvolumina von Affen, Menschen und vermuteten Übergangsgliedern greift beispielsweise zu kurz, es ist wichtig, dazu die Abhängigkeit des Hirnvolumens vom Körpergewicht darzustellen, auch wenn der Unterschied zwischen Affe und Mensch dann nicht mehr so prägnant ist.
Dann wechselt der Film die Thematik in seinen dritten Teil. Es geht um die genetische Ähnlichkeit von Menschen und Menschenaffen sowie um die umstrittene Wirkung von Mutationen. Die moderne Biologie hat dazu eine gewaltige Fülle von Daten erarbeitet. Nach meiner Meinung wäre neben den evolutionskritischen Ausführungen aber auch eine kurze Würdigung der Argumente für Evolution angemessen gewesen, die man aus den verfügbaren Daten ableiten kann. Der Film endet mit einem kurzen Überblick über die „Intelligent Design“-Idee, die das Leben – im Gegensatz zur Evolutionsanschauung – auf dem Hintergrund des Planungs-Aspektes einer überlegenen Intelligenz versteht.
Als ich die DVD abspielte, gewann – neben vielen positiven Eindrücken – eine Befürchtung jedoch immer mehr an Gestalt. Würde der Zuschauer nicht einen zu einseitigen Eindruck gewinnen, wenn grausame Kreuzungsversuche, eine plumpe Fossilfälschung (Piltdown) und Fehlinterpretationen (Ramapithecus, Neandertaler) zu stark im Mittelpunkt stehen? Würde beim Laien am Ende der diffuse Eindruck bleiben, Evolutionsbiologen seien allesamt Fälscher, Wissenschaftszensoren oder gar inhumane Gestalten, die verwerfliche Experimente mit Afrikanerinnen und Schimpansen auf dem Gewissen hätten? Es wäre schade, sehr schade, wenn der Film so wirken würde. Ethische Entgleisungen und Fehlinterpretationen sollen weder verschwiegen noch beschönigt werden, und die im Film genannten Beispiele sind im übrigen keineswegs erschöpfend. Solche traurigen Geschehnisse werden zurecht ihren Platz in der Geschichte der Biologie behalten (als Christ sehe ich mich leider veranlasst, in diesem Zusammenhang auch an die notvolle Geschichte der christlichen Kirchen zu erinnern). Doch hätte man dem Zuschauer nach meiner Meinung sagen müssen, dass die Mehrheit heutiger, experimentell arbeitender Evolutionsbiologen methodisch saubere und ethisch einwandfreie Arbeit leistet, durch die bedeutende wissenschaftliche Daten erhoben wurden und werden. Auch der Evolutionskritiker darf solche Leistungen anerkennen. Das schmälert die wissenschaftlichen Gegenargumente keineswegs, könnte aber zur dringend notwendigen Versachlichung der Diskussion beitragen. Ob die nachgeordnete evolutionäre Deutung der erhobenen Daten letztlich stichhaltig ist oder nicht, von welchen persönlichen und gesellschaftlichen Rahmenbedingungen sie beeinflusst wird, und welche Daten auf welche Weise gewichtet werden, das steht dann zur Diskussion (wer unter uns Biologen wäre nicht von seinen subjektiven weltanschaulichen Grundüberzeugungen beeinflusst, wenn er wissenschaftliche Daten in den Rahmen von Ursprungsmodellen stellt?).
Poppenbergs Film ist professionell produziert, technisch und didaktisch hervorragend aufgebaut und einfach spannend gemacht. Auch für den Laien ist er gut verständlich. So sehr ich also einerseits diesen Film begrüße und die Initiative von Fritz Poppenberg schätze, bleibt doch andererseits auch meine Befürchtung, es sei nicht alles gesagt worden, was notwendig und hilfreich gewesen wäre.
Der Film kann als VHS-Video oder als DVD bei der Studiengemeinschaft Wort und Wissen online bestellt werden:
https://www.wort-und-wissen.org/produkt/der-fall-des-affenmenschen/
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2005, http://www.genesisnet.info/schoepfung_evolution/n32.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.12.04 Film-Neuerscheinung „Der Fall des Affenmenschen“
Fritz Poppenberg, der durch seine Filme „Hat die Bibel doch recht?“ und „Gott würfelt nicht“ bekannter Filmproduzent, hat einen neuen Film zur Evolutionsthematik gedreht. Diesmal steht die Abstammung des Menschen im Mittelpunkt. Der Titel „Der Fall des Affenmenschen“ ist doppeldeutig, und diese beiden Bedeutungen greift der Film auch tatsächlich auf. Der „Fall Affenmensch“ wird aufgerollt: Aufhänger sind Berichte, wonach sowjetische Wissenschaftler versucht haben sollen, Kreuzungen zwischen Menschen und Affen durchzuführen. Poppenberg schildert seine Recherchen in dieser Sache, die dann aber zu keinem eindeutigen Ergebnis führten.
Doch das ist nur ein Aufhänger. Der Film ergeht sich nicht in Sensationsmache, sondern befasst sich mit den wichtigsten Indizien, die evolutionstheoretisch für eine Abstammung des Menschen aus dem Tierreich vorgebracht werden: Fossilfunde und Ähnlichkeiten zwischen heute lebenden Menschen und Affen. Schwerpunkt ist dabei die kritische Sichtung der fossilen Belege. Hier kommen Dr. Michael Brandt und Dr. Sigrid Hartwig-Scherer, Mitarbeiter der Studiengemeinschaft Wort und Wissen und Autoren von Fachbüchern zum Ursprung des Menschen, zu Wort. Quintessenz ihrer Darlegungen, die sie anhand von Fossilabgüssen anschaulich demonstrieren:
Die zunehmende Anzahl von Fossilfunden hat den Graben zwischen Menschenaffen und Menschen eher deutlicher hervortreten lassen als geschlossen. Auch in diesem Sinne gibt es einen „Fall des Affenmenschen“. Dieser Hauptteil des Films gibt die Diskussionslage auf verständlichem Niveau auf realistische Weise wieder.
Zum Thema „Ähnlichkeit zwischen heutigen Menschen und Affen“ kommt vor allem der Kölner Genetiker Dr. Wolf-Ekkehard Lönnig zu Wort. Er weist darauf hin, dass neben manchen verblüffenden Ähnlichkeiten zwischen Menschen und Schimpansen auch deutliche Unterschiede festzustellen seien, von denen man jedoch allgemein wenig erfahre. In diesem Teil des Films klingen manche Kommentare des Produzenten so, als würden in der Wissenschaft relevante Fakten unterdrückt; das dürfte übertrieben sein; richtig ist aber, dass Daten, die auf eine große Nähe zwischen Menschen und Affen hinweisen, mehr betont werden als andere, die deutliche Unterschiede erkennen lassen, und die es eben auch gibt.
Abschließend wird kurz das Thema „Intelligent Design“ als Alternative zu evolutionären Entstehungsmechanismen angeschnitten.
Der in weiten Teilen spannende Film ist für ein Laienpublikum und Jugendliche ab 14-15 Jahren gut geeignet und empfehlenswert. Kritische Zuschauer können darauf aufmerksam gemacht werden, dass die Behauptungen dieses Films, die natürlich nur beispielhaft belegt werden können, durch entsprechende Fachliteratur abgesichert sind.
(Dazu: http://www.wort-und-wissen.de/si/bio/gehirn.html
http://www.wort-und-wissen.de/si/bio/gang.html
http://www.wort-und-wissen.de/si/bio/ramapi.html)
Der Film kann als VHS-Video oder als DVD bei der Studiengemeinschaft Wort und Wissen online bestellt werden:
https://www.wort-und-wissen.org/produkt/der-fall-des-affenmenschen/
Autor dieser News: Reinhard Junker
© 2004, http://www.genesisnet.info/schoepfung_evolution/n30.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/