
Paläontologie: Fossilien des Erdmittelalters allgemein
18.12.20 Chamäleon-ähnlicher Beutefang mit Zungenschleuder bereits in der Kreide
Die vor allem auf dem afrikanischen Kontinent und angrenzenden Gebieten beheimateten Chamäleons fangen ihre Beute mit einer sehr speziellen und komplexen Methode. Haben sie ein entsprechendes Insekt fixiert, so können sie es mit ihrer Zunge erfassen, die sie bis zum 2,5-fachen ihrer Körperlänge herausschleudern und sie in ihr Maul befördern. Bernsteininklusen aus der Kreide von Myanmar zeigen, dass solche Mechanismen bereits damals etabliert waren.
Ein faszinierender Bernsteineinschluss zeigt fossile Hinweise auf einen bei heute lebenden exotischen Tieren bekannten Jagdmechanismus. Im Rahmen einer Studie an zwölf fossilen Eidechsen in Bernstein aus der Kreide (ca. 100 Millionen radiometrische Jahre; MrJ) von Myanmar beschrieben Daza et al. (2016) auch eine kleine Eidechsen-Inkluse (10,6 mm Länge). Diese Eidechse hatte einen relativ großen Kopf, kaum erkennbare Schuppen, gut erhaltene Klauen und wies einen auffallend kurzen, gebogenen Schwanz auf. Die Autoren sahen darin ein neugeborenes (neonatal) Tier, das aufgrund von Ähnlichkeitsvergleichen als zur Stammgruppe der Chamäleons gehörig interpretiert wurde.
Kürzlich berichteten Daza et al. (2020) von einem in Bernstein eingeschlossenen Kopf eines Amphibiums aus der Familie der Albanerpetontidae ebenfalls aus der Kreide von Myanmar. Die Albanerpetontidae sind eine nur fossil bekannte Amphibienfamilie, die den Schwanzlurchen zugerechnet wird und deren Arten kleinen Salamandern ähnlich sehen (Gardener & Böhme 2008). Der dreidimensional erhaltene Schädel ist von der Schnauzenspitze bis zum Hinterkopf 12,2 mm lang. Die Autoren schätzen die Körperlänge (Schnauze bis Becken) durch Vergleich mit einem anderen fossilen Albanerpetontiden (Celtedens ibericus; McGowan & Evans 1995) auf 52 mm. Das Tier wurde Yaksha peretti1 benannt. Im Bernstein ist der enthaltene Kopf nicht wirklich gut zu sehen. Ein hochaufgelöstes computertomographisches Bild lässt allerdings erstaunliche Details erkennen. Am bezahnten Unterkiefer ist die Verwachsung (Symphyse) von linkem und rechtem Unterkieferteil (am Kinn) verzahnt, was auf eine gewisse Flexibilität schließen lässt. Sehr auffällig ist eine knorpelartige Verlängerung des Zungenbeins (hyoid entoglossal process, hep). Dieselbe Struktur war bei dem von Daza et al. (2016) beschriebenen neugeborenen Tier aufgefallen und war der Hauptgrund, warum es als zur Stammgruppe der Chamäleons gehörig eingestuft wurde.
Das Zungenbein ist bei Wirbeltieren nicht direkt mit dem übrigen Skelett verknüpft, sondern bildet Ansatzstellen für Muskeln und wird dadurch in seiner Position gehalten. Der Fortsatz des Zungenbeins spielt bei der für Chamäleons typischen Art, Beute zu fangen, eine grundlegende Rolle. Sie können ihre Zunge herausschleudern und dabei eine Beute in einer Entfernung von ein bis 2,5 Körperlängen zielsicher treffen und ins eigene Maul befördern. Der gesamte Prozess hat eine extreme Dynamik und läuft im Bereich von wenigen Millisekunden ab, so dass den Beutetieren nur sehr geringe Fluchtchancen bleiben.
Der Mechanismus der Zungenschleuder von Chamäleons ist intensiv untersucht worden, auch mit dem Ziel, diesen für technische Problemlösungen zu nutzen. Im Aufbau zeigt der Zungenapparat eines Chamäleons, dass unmittelbar auf dem Fortsatz des Zungenbeins mehrere Lagen aus Bändern von Kollagenfasern liegen, die ihrerseits von einem Muskelschlauch umgeben sind. Zunächst war nicht klar, wie das Chamäleon seine Zunge mit einer Geschwindigkeit von bis zu 500 m s-1 in Richtung des erspähten Beutetiers abschießen kann, denn allein mit der von den vorhandenen Muskeln gelieferten Energie ist eine solche Beschleunigung nicht zu erreichen. Beobachtungen und entsprechende Modellversuche haben gezeigt, dass der Muskelschlauch den hep nicht komplett bedeckt, sondern das hep zur Mundöffnung hin noch aus dem Muskelschlauch herausragt. (Weiteres, über dem Muskelschlauch liegendes Zungengewebe umschließt den hep vollständig und mündet in der Zungenspitze, dem Zungenpolster.)
Hat das Chamäleon nun ein Beutetier erspäht, so kontrahiert es den Muskelschlauch, der sich dadurch verlängert und sich dem Vorderende des hep nähert. Beim Tier beobachtet man, dass es den Mund öffnet und die Zunge mitsamt dem vorderen Teil des hep aus dem Mund hervorsteht. Durch die Muskelkontraktion und die dadurch bewirkte Verlängerung des Muskelschlauchs werden die Kollagenbänder direkt über dem hep vorgespannt und speichern damit bereits Muskelarbeit. Der Abschuss der Zungenschleuder erfolgt in dem Moment, wenn der Muskelschlauch die abgerundete Spitze des hep erreicht. Dieser wird im selben Moment noch weiter durch die Mundöffnung nach außen geschoben. Durch weitere Kontraktion trägt der Muskelschlauch zu Beschleunigung des Zungengewebes bei. Ein wesentlicher Teil der Beschleunigungsenergie kommt aus der Vorspannung der Kollagenschichten. Durch entsprechende Muskelstränge wird der Zungenapparat – im Erfolgsfall mit der Beute – dann wieder in das geöffnete Maul befördert. Der Rückholvorgang ist verglichen mit dem Herausschleudern langsamer.
Kleine, lungenlose Salamander (Plethodontidae) nutzen eine interessante und noch schnellere Variante zum Beutefang ein. Sie können ihr hep falten und schießen es beim Zungenschuss wie ein Projektil mit dem Zungenapparat so heraus, dass es den Mund komplett verlässt. Dadurch erreichen diese kleinen Salamander noch höhere Geschwindigkeiten bei ihrem Zungenschuss.
Es ist nun sehr interessant, dass die CT-Aufnahmen des Yaksha peretti-Schädels den hep und sogar noch einen Teil des Zungenpolsters an der Spitze des hep abbilden. Daza et al. (2020) interpretieren diesen Befund als Hinweis, dass Y. peretti in ähnlichen Lebensräumen gelebt und mit ähnlichen Methoden Beute gejagt haben, wie wir das von Chamäleons heute kennen. Für die Albanerpetontiden wurden auch schon unterirdische Lebensräume diskutiert; dort würden solche Lebens- und Ernährungsweisen aber keinen Sinn ergeben.
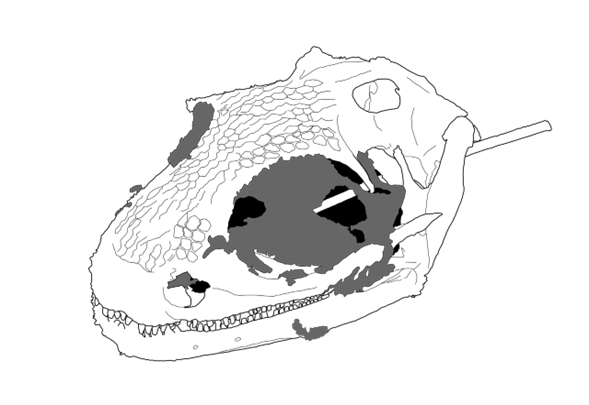
Abb. 1: Schädel des ausgestorbenen Albanerpetontiden Yaksha peretti, neu gezeichnet nach einem Bild aus der Pressemitteilung; das dunkle Grau steht für erhaltene organische Substanz. (Wikimedia: Hemiauchenia, CC BY-SA 4.0)
Es ist sehr interessant, dass wir schon aus der Zeit der Kreide Befunde haben, die eine hochspezialisierte Jagdtechnik wie die Zungenschleuder belegen und dass der zugrundeliegende Mechanismus heute die Bionik herausfordert und bereichert.
Literatur
Daza JD, Stanley EL, Wagner P, Bauer AM & Grimaldi DA (2016) Mid-Cretaceous amber fossils illuminate the past diversity of tropical lizards. Sci. Adv. 2: e1501080.
Daza JD, Stanley EL, Bolet A, Bauer AM, Arias JS, Cernansy A, Bevitt JJ, Wagner P & Evans SE (2020) Enigmatic amphibians in mid-Cretaceous amber were chameleon-like feeders. Science 370, 687-691.
Gardener JD & Böhme M (2008) Review of Albanerpetontidae (Lissamphibia) with comments on the paleoecological preferences of European Tertiary Albanerpentotids. In: Sankey JT & Baszio S (Hg.) Vertebrate Microfossil Assemblages – Their Role in Paleoecology and Paleobiogeography. University of Indiana Press, Bloomington, 2008, p. 178-218.
McGowan G & Evans SE (1995) Albanerpetontid amphibians from the Cretaceous of Spain. Nature 373, 143-145.
Anschauliche Erklärung des Zungenschuss von Chamäleons: https://www.madcham.de/de/der-zungenschuss/
Anmerkung
1 „Yaksha“ ist ein Geist, ein Wächter natürlicher Kostbarkeiten, der in der Erde oder in Baumwurzeln verborgen ist; mit „peretti“ wird Adolf Peretti für seine Verdienste im Zusammenhang mit der Erforschung von Bernstein inklusen geehrt. Dieser gründete 1996 das GEM RESEARCH SWISSLAB (GRS; gemresearch.ch). Der hier vorgestellte Schädel ist als Holotyp dort gelagert und wird als Herzstück der dortigen Ausstellung präsentiert. Eine Dokumentation der Arbeit ist online verfügbar: https://www.youtube.com/watch?v=jO2cHlujLDM&feature=youtu.be
Autor dieser News: Harald Binder
© 2020, http://www.genesisnet.info/schoepfung_evolution/n283.php
30.01.18 Paradebeispiel für Co-Evolution in Frage gestellt
Bisher galt als Lehrmeinung, dass Schmetterlinge und bedecktsamige Blütenpflanzen „Hand in Hand“ evolvierten. Doch nun zeigen fossile Reste von Schmetterlingsschuppen, dass die einen Saugrüssel tragenden Schmetterlinge sehr viel früher existiert haben. Der Fall zeigt beispielhaft: Stimmige „Geschichten“ sind keine Erklärungen und es muss mit umfangreichen geologisch nicht überlieferten Lebensräumen gerechnet werden.
Schmetterlinge (Lepidoptera) sind der Inbegriff für Symbiose, d. h. eine Vergesellschaftung von Individuen zweier verschiedener Arten, die für beide Partner vorteilhaft ist. Die nektarsaugenden Falter nehmen mit ihrem Saugrüssel beim Blütenbesuch auch Pollen auf bzw. geben ihn ab und ermöglichen auf diese Weise bei vielen Pflanzen die Bestäubung. Sie selber erhalten nahrhaften Nektar aus den Blüten. Was liegt im Rahmen einer evolutionären Weltsicht näher als die Annahme, dass die Entstehung von Schmetterlingen und nektarproduzierenden Blütenpflanzen (Angiospermen) Hand in Hand ging? Diese Geschichte war bisher Lehrmeinung und mit den bekannten Fossilfunden vereinbar. Allerdings ist der Fossilbericht dieser Insektengruppe sehr dürftig. Der bislang älteste Fund stammte aus der Unterkreide und wird auf 129 Millionen radiometrische Jahre (MrJ) datiert. Molekulare Studien sprechen unter evolutionstheoretischen Voraussetzungen jedoch dafür, dass die Lepidopteren schon sehr viel früher entstanden sind als die Angiospermen, nämlich (je nach verwendeter Methode) im Perm oder in der oberen Trias (> 200 MrJ), während eindeutige Angiospermen-(Makro-)Fossilien erst ab der Unterkreide bekannt sind.
Nun wurden in der Nähe von Braunschweig in einem Sedimentbohrkern aus dem Bereich der Grenze Trias / Jura unter anderen Mikrofossilien zarte, schuppenähnliche Gebilde entdeckt (Van Eldijk et al. 2018). Etwa 70 Schuppen konnten genauer analysiert werden. Die Untersuchung ergab, dass einige von diesen Schuppen Merkmale besaßen, die typisch sind für Flügelschuppen der Gruppe der Glossata, den Motten und Schmetterlingen mit Saugrüssel. Der Besitz solcher Schuppen, die Flügel, Beine und den Körper bedecken, ist das namengebende Merkmal der Lepidoptera – „Schuppenflügler“ (gr. lepis, Schuppe). Besonders die auf den Flügeln sitzenden Schuppen sind taxonomisch informativ, d. h. sie erlauben eine recht genaue Zuordnung zu einer bestimmten Gruppe. Damit ist nun der fossile Nachweis erbracht, dass Schmetterlinge 70 MrJ früher existiert haben als bisher bekannt und ca. 50 MrJ vor dem ersten Nachweis von Angiospermen. Die Geschichte von der Co-Evolution von zwittrigen, nektarführenden Blüten und Insekten mit Saugrüssel passt also nicht mehr zum fossilen Befund.
Wie könnte eine neue „Geschichte“ aussehen? Die Forscher mutmaßen, dass der Saugrüssel anfangs als Trinkwerkzeug evolvierte, mit welchem Tropfen von pflanzlichem Saft aufgenommen werden könnten (z. B. Bestäubungstropfen von Nadelbäumen, mit denen Pollen aufgefangen werden, oder bei Verletzungen von Pflanzen austretende Flüssigkeit). Es gibt Hinweise auf große Trockenheit in jener Zeit, weshalb der Besitz eines Saugrüssels vorteilhaft gewesen sei. Der Nutzen war zuerst einseitig nur beim Insekt. Erst später sei der Saugrüssel dann für die Aufnahme von Nektar mit gegenseitigem Nutzen (Bestäubung) verwendet worden.
Diskussion. Die Umwandlung der zuvor beißenden Mundwerkzeuge in ein Saugorgan wird von Van Eldijk et al. (2018) als evolutionäre Antwort auf Trockenheit beschrieben. Wie die Umwandlung erfolgte, wird nicht thematisiert. Es ist einfach eine neue Geschichte, die an die Stelle der alten getreten ist. Solche Geschichten sind keine Erklärungen, sie können sich jederzeit durch neue Funde als falsch erweisen. Sie sind vage, gehen nicht in Details und wirken geradezu „undarwinistisch“, weil der Eindruck erweckt wird, dass als Folge veränderter Bedürfnisse (hier Trockenheit) passende Reaktionen zielgerichtet möglich seien.1 Das erinnert sehr an Lamarcks hypothetischen Ansatz, wonach Umweltreize einen direkten Einfluss auf den optimalen und angepassten Gestaltwandel der Organismen ausgeübt haben sollen. Die neue Geschichte ist zudem wenig glaubwürdig, da viele andere Insekten zur gleichen Zeit ohne saugende Werkzeuge auskamen. Warum also sollte sich gerade in der Linie, die die Lepidopteren hervorbrachte, ein Saugrüssel bilden? Das ist völlig unklar, zumal Angebote von Flüssigkeit zum Saugen auch schon vorher vorhanden waren. Sowohl die bisherige als auch die neue Geschichte sind typische Beispiele von „adaptivem Storytelling“. Die bisherige Geschichte ist im Übrigen gar nicht widerlegt, wie behauptet wird. Würde man nämlich in Schichten der Trias Angiospermen finden, würde sie wieder passen. Dafür gibt es sogar Hinweise (Hochuli & Feist-Burkhardt 2013; vgl. Kutzelnigg 2014), auf die die Autoren selber hinweisen (van Eldijk et al. 2018, 52).
In jedem Fall zeigen die neuen Funde, dass mit umfangreichen fossil nicht überlieferten Lebensräumen gerechnet werden muss, da es eine Lücke von 70 MrJ zwischen den stratigraphisch ältesten und zweitältesten fossilen Nachweisen der Lepidopteren gibt. Auch der große zeitliche Versatz zwischen dem erstmaligen fossilen Auftreten von angiospermenartigem Pollen und dazu passenden Makrofossilien deutet auf fossil nicht überlieferte Lebensräume hin.
Die Daten laden darüber hinaus auch dazu ein, über einen prinzipiell anderen Deutungszugang bezüglich der geologischen Überlieferung nachzudenken, der u.a. ohne Evolution und lange Zeiträume als paradigmatischen Rahmen auskommt. Der Autor ist sich bewusst, dass dazu im Rahmen der Schöpfungsforschung bisher nur sehr wenig vorgelegt werden konnte.
Anmerkungen
1 „The transition to exclusively feeding on liquids was most likely an evolutionary response to widespread heat and aridity during the Norian … substitution of mandibulate mouthparts by a sucking proboscis could be seen as an adaptation to adequate maintenance of body water balance of small, short-lived moths. … short and simply composed proboscides of ancestral Glossata must have been used initially to drink from water droplets or sap from injured leaves“ (Van Eldijk 2018, 4).
„Um an den Nektar in den Blüten heranzukommen, wandelten einige Insektengruppen, darunter auch die Schmetterlinge, ihre zuvor beißenden Mundwerkzeuge in einen Saugrüssel um.“ (http://www.scinexx.de/wissen-aktuell-22292-2018-01-11.html
2 „It should be noted that the wide morphological diversity of the dispersed Triassic-Jurassic pollen record strongly suggests that, perhaps, entire families or orders of seed plants have still escaped sampling in the coeval megafossil record. Some pollen types have angiosperm-like morphological characters. Although affinity to angiosperm crowngroups remains questionable, notably, the presence of a reticulate wall structure might be functionally linked to pollination by flying insects“ (Van Eldijk et al. 2018, 5).
Literatur
Hochuli PA & Feist-Burkhardt S (2004) A boreal early cradle of Angiosperms. Angiosperm-like pollen from the Middle Triassic of the Barents Sea (Norway). J. Micropalaeontol. 23, 97-104.
Kutzelnigg H (2014) Sind Blütenpflanzen 100 Millionen Jahre älter als bisher angenommen? Stud. Integr. J. 21, 41-43.
Van Eldijk TJB, Wappler T et al. (2018) A Triassic-Jurassic window into the evolution of Lepidoptera. Sci. Adv. 2018;4: e1701568.
Autor dieser News: Reinhard Junker
© 2018, http://www.genesisnet.info/schoepfung_evolution/n255.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
14.04.14 Königsfarn – 180 Millionen radiometrische Jahre lang unverändert
Der Fund eines fossilen Königsfarns aus dem schwedischen Unterjura (180 Millionen radiometrische Jahre) stellt bisher bekannte Beispiele lebender Fossilien aufgrund detaillierter Erhaltung selbst von subzellulären Strukturen in den Schatten. Dass auch zelluläre Details über einen mutmaßlich so großen Zeitraum nahezu unverändert bleiben, wirft einige Fragen auf.

Abb. 1: Königsfarn (Osmunda regalis). (Wikimedia: Christian Fischer, CC BY-SA 3.0)
Lebewesen, die als Fossilien bekannt sind, aber auch heute in sehr ähnlicher Form existieren, üben eine besondere Faszination aus. Der Zahn der Zeit scheint ihnen nichts anhaben zu können. Solche „lebenden Fossilien“ werden in evolutionärer Perspektive eher als Ausnahmen angesehen – widerstanden sie doch viele Millionen Jahre lang dem scheinbar unausweichlichen Drang der Natur zur Veränderung, der – so die Ansicht der Evolutionsbiologie – zur immensen Vielfalt an Formen geführt haben soll. Wenn also Evolution das Normale ist, wären unverändert lebende Fossilien als Ausnahme zu kennzeichnen. Was aber, wenn die Ausnahme zur Regel wird? Denn lebende Fossilien sind keineswegs so selten, mindestens wenn man den Vergleich zwischen heute lebend und fossil nicht zu eng auf der Art-Ebene ansetzt, sondern auf der Ebene von Grundtypen. Wie kommt es, dass der mutmaßlich lang anhaltende Prozess der Evolution bei so vielen Organismen irgendwann zum weitgehenden Dauerstillstand gekommen ist? Ist eine innovative Evolution wirklich eine Grundeigenschaft des Lebens? Oder bestätigen lebende Fossilien nicht gerade das Gegenteil, dass den Veränderungen der Organismen Grenzen gesetzt sind?
Aus der Sicht der Schöpfungslehre ist das häufige Vorkommen lebender Fossilien nicht überraschend, ja dies kann sogar als eine Voraussage verstanden werden, die durch Forschung belegt oder auch widerlegt werden kann.
Auch wenn lebende Fossilen also keineswegs selten sind, ist der Fund eines „lebenden Fossils“ eines Königfarns, über den Bomfleur et al. (2014) berichten, bisher einzigartig und höchst erstaunlich. Dass Königsfarne (Familie Osmundaceae) sich seit dem unteren Mesozoikum (220 Millionen radiometrische Jahre) in ihrem Bau fast nicht verändert haben, ist schon länger bekannt. Der jüngst beschriebene Fund aus dem schwedischen Unterjura (180 Millionen radiometrische Jahre) stellt bisher bekannte Beispiele lebender Fossilien aufgrund detaillierter Erhaltung selbst von subzellulären Strukturen jedoch in den Schatten. Ein etwa 6 cm langes Rhizom (Erdspross) von ca. 7 mm Durchmesser und zahlreichen Ansätzen von Blattwedeln lässt aufgrund besonderer Erhaltung nicht nur die Zellwände sehr gut erkennen, sondern auch Zellkerne und Zellorganellen. Und es kommt noch besser: Einige der Zellen waren gerade in Teilung begriffen, so dass sogar spezifische Chromosomen in verschiedenen Teilungsstadien identifiziert werden können. Die Größe der fossilen Zellkerne während der Interphase entspricht der Größe heutiger Osmundaceae. Die Autoren schließen daraus, dass sich die Genomgröße 180 Millionen Jahre lang nicht geändert hat, ein „hervorragendes Beispiel eines evolutionären Stillstands“ (Bomfleur et al. 2014, 1376). Weder Vervielfachung des Erbguts noch nennenswerte Genverluste seien vorgekommen – der evolutionäre Stillstand ist nahezu total. „Wenn wir die feinen Zellstrukturen dieses 180 Millionen Jahre alten Fossils mit denen heute lebender Königsfarne vergleichen, sind sie eigentlich identisch“, wird Bomfleur im Deutschlandfunk zitiert (https://www.deutschlandfunk.de/lebende-fossilien-koenigsfarne-haben-sich-ueber-100.html).
Möglich wurde die hervorragende Erhaltung dadurch, dass durch eine vulkanische Schlammlawine vermutlich 70-120oC Grad Celsius heiße, mit Mineralen beladene vulkanische Wässer Kontakt mit den Pflanzen bekamen und dadurch Kalzit auskristallisiert wurde. Das ermöglichte die Konservierung des Inhalts der Zellen bis in feinste Details. Die Fossilisierung muss ungewöhnlich schnell innerhalb weniger Minuten abgelaufen sein.
Lebende Fossilien wie die Königfarne sind eine Herausforderung für den Geltungsanspruch einer evolutionsbiologischen Interpretation des Lebens. Darüber hinaus ergeben sich aber auch Anfragen dahingehend, ob die ermittelten Zeiträume wirklich reale Zeitabschnitte repräsentieren. Diese Anfrage wiegt umso schwerer, je vehementer man auf die Evolution als eine Grundeigenschaft des Lebens verweist.
Literatur
Bomfleur B, McLoughlin S & Vajda V (2014) Fossilized nuclei and chromosomes reveal 180 million years of genomic stasis in royal ferns. Science 343, 1376-1377.
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n209.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
25.11.13 Sind Blütenpflanzen 100 Millionen Jahre älter als bisher angenommen?
Gastbeitrag von Herfried Kutzelnigg
„Blütenpflanzen sind 100 Millionen Jahre älter als bisher angenommen“ – so oder ähnlich lauten verschiedene Pressemitteilungen über Funde von 240 Millionen Jahre altem Pollen, den Peter A. Hochuli (Universität Zürich) und Susanne Feist-Burkhardt (Ober-Ramstadt) in Bohrkernen der Nordschweiz gefunden haben. Die sehr bemerkenswerten Ergebnisse wurden vor kurzem in der Zeitschrift „Frontiers in Plant Science“ vorgestellt (Hochuli & Feist-Burkhardt 2013). Sie sind gut dokumentiert, und ihre mögliche Bedeutung für die Abstammung der Bedecktsamigen Blütenpflanzen wird sorgfältig diskutiert. Der immense zeitliche Unterschied zwischen der fossilen Überlieferung von Pollen und dazu gehörenden Makrofossilien legt nahe, dass es in erheblichem Umfang geologisch nicht überlieferte Lebensräume gibt.
Hintergrund: Bedecktsamige Blütenpflanzen (Angiospermen) sind im Fossilbericht erst seit der unteren Kreide nachgewiesen, also seit ca. 140 Millionen rJ (= Jahren gemäß radiometrischer Datierung). Dabei stellen Pollenkörner die ältesten fossilen Angiospermen-Reste dar. Das lässt sich leicht dadurch erklären, dass Pollen sehr umweltbeständig ist, in großer Menge produziert wird, und leicht an Standorte gelangen kann, wo die Pflanzen selber nicht wachsen. Die bislang ältesten Angiospermen-Pollenkörner sind aber nur ca. 5 Mio. rJ älter als die ältesten Angiospermen-Makrofossilien. (Makrofossilien sind Überreste kompletter Pflanzen bzw. Pflanzenteile wie Blätter, Stängel oder Wurzeln.) Der zeitliche Versatz ist also relativ gering, ganz anders als bei den jüngst beschriebenen Funden (s. u.).
Während verschiedene Gruppen von Nacktsamigen Blütenpflanzen (Gymnospermen) schon viel länger fossil bekannt sind (z. T. schon seit dem Karbon), tauchen die Bedecktsamer – wie gesagt – fossil erstmals in der Unterkreide auf. Dort aber treten sie ziemlich von Beginn ihrer Überlieferung in großer Formenfülle auf. Gleichzeitig fehlt ein überzeugender morphologischer Anschluss an irgendeine andere Pflanzengruppe. Schon Darwin hatte erkannt, dass dieses unvermittelte Auftreten der Angiospermen, immerhin die artenreichste Pflanzengruppe überhaupt, ein großes Problem für seine Abstammungslehre bedeuten würde. Er nannte das ein „abomenable mistery“, also ein „abscheuliches Geheimnis“. Bis heute ist man trotz intensiven Bemühens der Lösung des Problems nicht wirklich nähergekommen. Man vergleiche dazu etwa den Übersichtsartikel in Studium Integrale Journal (Kutzelnigg 2000, 2001), den Kurzbeitrag in genesisnet.info im Darwinjahr 2009 (Das „abscheuliche Geheimnis“ im Darwinjahr, https://www.genesisnet.info/index.php?News=131) oder Abschnitt VI.14.10 der soeben erschienenen Neuauflage von „Evolution – Ein kritisches Lehrbuch“ (https://www.wort-und-wissen.org/publikationen/lehrbuch/). Interessanterweise wird der Begriff „abomenable mistery“ noch heute gerne von Evolutionsvertretern benutzt.
Natürlich hat man intensiv nach Angiospermen-Resten in älteren geologischen Systemen als der Kreide gesucht, vor allem im Jura als dem nächst älteren. Es gab auch entsprechenden Meldungen, aber sie konnten allesamt nicht bestätigt werden.
Da man keine Fossilien fand, bekam eine auf Axelrod (1952) zurückgehende, nicht gerade alltägliche Hypothese Auftrieb. Danach sollen die Angiospermen wesentlich älter sein, als sie fossil belegt sind, weil sie zunächst an Standorten lebten, die fossil nicht erhalten wurden (vgl. Stephan 2002).
Solche Überlegungen bekamen Unterstützung durch molekulare Untersuchungen. Hier wurde versucht, nach dem Prinzip der molekularen Uhren das „tatsächliche“ Alter der frühesten Angiospermen zu berechnen. Hochuli & Feist-Burkhardt referieren den neuesten Stand entsprechender Bemühungen. Die Ergebnisse sind je nach Methode und verwendetem Datensatz sehr verschieden. So reicht der Beginn der Angiospermen entweder zurück bis in Jura (193.8 Mio. rJ), Trias (221.5 Mio. rJ) oder Perm (275 Mio rJ), um nur einige Beispiele zu nennen. – Am Rande sei bemerkt, dass solche gewaltigen Diskrepanzen Fragen über die Aussagekraft molekularer Datierungen aufwerfen.
Die aktuellen Funde: Die Pollenfunde von Hochuli & Feist-Burkhardt sind insofern sehr bemerkenswert, als sie aus der Trias (247-242 Mio. rJ) stammen und somit – wie gesagt – die ersten fossilen Reste unterhalb der Kreide darstellen, die wahrscheinlich Angiospermen zuzuordnen sind. Die sehr gut erhaltenen und mit modernsten Methoden optisch gut erfassten Pollenkörner zeigen im Feinbau der Pollenwand (Exine) sehr große Übereinstimmungen mit Angiospermen. Wie die ersten sonst nachgewiesenen Angiospermen-Pollenkörner besitzen sie nur eine Falte (Einfurchen-Pollen). Allerdings ist ein deutlicher Unterschied zu diesen durch die extrem dünne innerste Schicht gegeben. Sechs verschiedene Typen wurden festgestellt. Sie alle konnten keinem bisher bekannten Pollentyp zugeordnet werden. Außerdem wurde in den Trias-Schichten Pollen des Afropollis-Typs gefunden. Dieser konnte verschiedentlich auch in der Kreide nachgewiesen werden. Aber auch hierbei ist unklar, ob es sich um Gymnospermen oder Angiospermen handelt.
Die große Ähnlichkeit in der äußeren Pollenwand spricht bei den Neufunden sehr für die Zugehörigkeit zu den Angiospermen. Als Alternative bliebe eine bisher nicht bekannte Gruppe von Gymnospermen; aber das ist eher unwahrscheinlich. Entscheidend aber ist, dass die neu gefundenen Pollenkörner nicht als Vorläufer der Angiospermen betrachtet werden können, da sie vom Feinbau der Pollenwand her keinen Anschluss an frühe Angiospermen-Pollen darstellen. Im Übrigen sprechen auch einige andere Befunde gegen diese Denkmöglichkeit. Dazu gehört die beobachtete große Bandbreite der verschiedenen Typen und die aus den Begleitfunden von Gymnospermen-Pollen in den betreffenden Schichten zu schließende große Unterschiedlichkeit der zugehörigen Standorte. Außerdem hatten die Autoren schon früher in ähnlichen Trias-Abfolgen der Barentssee (Norwegen) vergleichbare Pollenkörner gefunden (Hochuli & Feist-Burkhardt 2004). Dabei ist bemerkenswert, dass die dort als Begleitelemente gefundenen Sporen ein feuchtes Klima dokumentieren, während für die Schweizer Funde ein eher trockenes Klima anzunehmen ist.
Fazit: Die neuen mit ca. 240 Mio. rJ datierten Pollenfunde aus der Mittleren Trias der Nordschweiz und der Barentssee sind sehr bemerkenswert, da sie höchstwahrscheinlich Angiospermen zuzuordnen sind. Im Vergleich dazu stammen die ältesten bis dahin bekannten Pollenfunde und Makrofossilien von Angiospermen aus der Unterkreide (ca. 140-135 Mio. rJ). Es klafft also eine enorme Überlieferungslücke von ca. 100 Mio. rJ. Wie die zugehörigen Pflanzen ausgesehen haben, ist völlig unklar. Sicher ist aber von der Feinstruktur der Pollenwand her, dass es sich nicht um Vorläufer jener Pollentypen handelt, die man bei den für ursprünglich gehaltenen Angiospermen findet. Auch bleibt die Frage offen, wieso es über einen Zeitraum, der den gesamten Jura und die Untertrias umfasst, keine fossilen Nachweise gibt. Der Gedanke an geologisch nicht überlieferte Lebensräume liegt hier nahe (vgl. Stephan 2002). Außerdem passt die ökologische Bandbreite nicht ohne weiteres zu dem oben genannten Konzept von Axelrod, wonach Angiospermen-Fossilien in älteren Systemen als Kreide nur deshalb fehlen, weil sie zu der Zeit an sehr speziellen Standorten wuchsen.
Insofern sind die Neufunde sehr bemerkenswert, weil sie darauf hinweisen, dass ganze Organismengruppen über große Zeiträume bzw. über viele geologische Schichtglieder hinweg ohne fossile Dokumentation existieren können. Zur Frage der Abstammung der Angiospermen bringen sie aber keine entscheidende Klärung.
Literatur
Axelrod DI (1952) A theory of angiosperm evolution. Evolution 6, 29-60.
Hochuli PA & Feist-Burkhardt S (2004) A boreal early cradle of Angiosperms. Angiosperm-like pollen from the Middle Triassic of the Barents Sea (Norway). J. Micropalaeontol. 23, 97-104.
Hochuli PA & Feist-Burkhardt S (2013) Angiosperm-like pollen and Afropollis from the middle triassic of the Germanic basin (Northern Switzerland). Frontiers in Plant Science. doi: 10.3389/fpls.2013.00344.
Kutzelnigg H (2000, 2001): Das „abscheuliche Geheimnis“. Woher kommen die Angiospermen? Stud. Int. J. 7, 51-58; 8, 10-15. (http://www.wort-und-wissen.de/sij/sij72/sij72-1m.html)
Stephan M (2002) Der Mensch und die geologische Zeittafel. Holzgerlingen.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2013, http://www.genesisnet.info/schoepfung_evolution/n203.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
14.10.11 Über den Ursprung der Schildkröten
Schildkröten haben einen unverwechselbaren Bauplan, der sich deutlich von den Bauplänen aller anderen Wirbeltiere absetzt. Ein neuer Genesisnet-Artikel (Artikel Entstehung der Schildkröten, https://www.genesisnet.info/index.php?Artikel=42847&Sprache=de&l=2) diskutiert Hypothesen über ihre Entstehung.
Schildkröten haben den ungewöhnlichsten Bauplan unter den Wirbeltieren. Er ist so verschieden von anderen Tiergruppen, dass über die taxonomische* Zuordnung keine eindeutige Aussage möglich ist. Nachdem morphologische** Merkmale eher Verwandtschaftsbeziehungen mit stammesgeschichtlich alten Gruppen unterstützten, wiesen neuere molekularbiologische Untersuchungen ab Ende der 1990er Jahre in eine neue Richtung, nämlich einer Verwandtschaft mit diapsiden Formen, die als abgeleitet und jünger gelten. Morphologische und molekulare Analysen passen nicht zusammen.
*taxonomisch (= die Ordnung der Lebewesen betreffend)
**morphologisch (= gestaltliche)
Über die Entstehung des hervorstechendsten Merkmals, den Panzer, werden unterschiedliche Hypothesen vertreten. Nach der Transformationshypothese entstand der Panzer allmählich aus Hautknochen durch Fusion mit den Rippen und Wirbelfortsätzen unter der Wirkung der natürlichen Selektion. Nach der Emergenzhypothese ist der Panzer dagegen zunächst unabhängig von Anpassungsprozessen durch Neuverschaltungen von Steuergenen und Entwicklungsmodulen, also durch Veränderungen in der ontogenetischen* Entwicklung entstanden. Eine besondere Rolle spielt nach dieser Hypothese eine nur in der Ontogenese* der Schildkröten vorkommende Aufwölbung aus Ektoderm und Mesoderm, die sich oberhalb der Extremitätenknospe in Längsrichtung auf der Körperflanke bildet, die Carapaxfalte. Beiden Hypothesen stehen schwerwiegende Probleme entgegen. Das Für und Wider der beiden Hypothesen wird erläutert.
*Ontogenese (= individuelle Entwicklung vom Ei zum ausgewachsenen Organismus)
In der Fossilüberlieferung erscheint der Schildkrötenbauplan ziemlich abrupt. Neuere Fossilfunde könnten die Lücken zu anderen Tiergruppen verringern. Die in den Jahren 2008 und 2009 neu beschriebenen Gattungen Odontochelys und Chinlechelys werden im Rahmen der konkurrierenden Panzer-Entstehungshypothesen diskutiert. Diese beiden Gattungen besaßen Merkmale, aus denen Hinweise auf ihre hypothetische Evolution entnommen werden können. So besaß Odontochelys keinen verknöcherten Rückenpanzer, und der Panzer von Chinlechelys war sehr dünn und nur teilweise mit den Rippen verwachsen. Es ist aber in beiden Fällen nicht sicher, ob es sich dabei um phylogenetische Primitivmerkmale handelt. Odontochelys könnte auch eine spezialisierte wasserlebende Gattung gewesen sein. Und die fossilen Reste von Chinlechelys sind möglicherweise zu spärlich, um sichere Schlussfolgerungen zu erlauben.
Anschließend wird eine Reihe von Fragen zur Evolution der Schildkröten zusammenfassend diskutiert: Ontogenetische Befunde und die Bedeutung der Carapaxfalte, die uneinheitliche systematische Stellung der ältesten Fossilfunde, molekulare Daten und der Fossilbefund, einzigartige Merkmalskombinationen und Konvergenzen*, Probleme des Umbaus zum Panzer und die Ontogenese als fragwürdiges Modell für die Stammesgeschichte.
Abschließend werden kurz einige spekulative Ideen für die Möglichkeit einer nicht-evolutionären Deutung der Fossilüberlieferung der ältesten Schildkröten diskutiert.
*Konvergenzen (= unabhängig entstandene Ähnlichkeiten)
Der Artikel liegt in zwei unterschiedlichen Versionen vor und erschien in ähnlicher Form in zwei Folgen in der Zeitschrift Studium Integrale Journal (http://www.si-journal.de/index2.php?artikel=jg17/heft1/sij171-3.html).
„Experten“-Version: Entstehung der Schildkröten
PDF: Entstehung der Schildkröten
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2011, http://www.genesisnet.info/schoepfung_evolution/n177.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
09.06.09 Das „abscheuliche Geheimnis“ im Darwinjahr
Aktuell im Darwinjahr: Die Frage nach der Abstammung der Blütenpflanzen ist nach wie vor ungeklärt
Gastbeitrag von Herfried Kutzelnigg
Eines der großen Probleme der Darwinschen Abstammungslehre ist die Herkunft der Bedecktsamigen Blütenpflanzen (Angiospermen), die heute mit ihren fast 300.000 Arten den weitaus größten Teil der grünen Pflanzen ausmachen. Sie treten schlagartig und ohne erkennbare Zwischenstufen zu Beginn der Kreideformation vor ca. 100 Millionen Jahren herkömmlicher Zeitrechnung auf und haben in vergleichsweise kurzer Zeit die bekannte Formenfülle erreicht. Darwin erkannte in selbstkritischer Weise, dass dieses Problem seine Theorie erheblich in Frage stellen würde und bezeichnete daher einem Kollegen gegenüber in einem Brief aus dem Jahre 1879 die Situation als „abominable mistery“, also als abscheuliches Geheimnis. Vor allem störte ihn das sprunghafte Auftreten dieser Pflanzen, das so gar nicht in das Konzept einer Evolution passte, die lange Zeiträume zu ihrer Realisierung erfordert.
Nach nunmehr 130 Jahren intensiver Forschung ist das Geheimnis nach wie vor ungelüftet. Umso erstaunlicher ist es, dass der Ausdruck „abominable mistery“ auch von Evolutionsbefürwortern gerne benutzt wird. Offenbar ist man so unerschütterlich fest von der Theorie überzeugt, dass man es nur für eine Frage der Zeit hält, wann das Geheimnis gelüftet sein wird.
Entsprechend ist auch im Darwinjahr mehrfach über das Thema geschrieben worden, z. B. in einem Artikel von Elizabeth Pennisi in der führenden Wissenschaftszeitschrift Science in der Ausgabe vom 3. April 2009 mit dem Titel „On the Origin of Flowering Plants“. Es ist in solchen Artikeln immer wieder eindrucksvoll, wie ausführlich darüber berichtet wird, dass man trotz moderner Forschung in all den verschiedenen Teildisziplinen noch nicht weiter ist, aber gleichzeitig mit beachtlichem Selbstbewusstsein darauf hingewiesen wird, dass man kurz vor dem Ziel sei.
Nun kurz zu einigen Teilaspekten der Problematik: Woher die Bedecktsamer kommen, ist weiterhin völlig offen. Hoffnungen, die Vorfahren bei irgendwelchen Gruppen der Nacktsamer (Gymnospermen) zu finden, wurden sämtlich zerschlagen. Dies gilt sowohl für die Suche nach Fossilfunden, die man als Zwischenstufen interpretieren könnte, als auch für theoretische Ableitungen der Blütenherkunft und auch für Überlegungen aufgrund des Vergleichs von molekularen Daten. Als unbefriedigende Lösung blieb nur übrig, das Problem zeitlich zurück zu verschieben und jetzt von einem gemeinsamen Ahnen der Angiospermen und Gymnospermen auszugehen.
Wie die ursprüngliche Angiospermenblüte ausgesehen haben könnte, ist wegen der explosionsartigen Ausbreitung dieser Pflanzengruppe nach wie vor unklar. Es scheint jetzt nur klar zu sein, dass die jahrzehntelang gelehrte Lehrbuchmeinung, wonach die primitiven Blüten in der Verwandtschaft der Magnolien zu suchen sind, aufgegeben werden muss.
Neuere Arbeiten über die Abstammung der Angiospermen nennen regelmäßig als großen Wissenschaftsfortschritt die in den letzten Jahren aufgestellten „Stammbäume“, die aufgrund morphologischer und vor allem molekularer Analysen konstruiert wurden. Dabei wird oft übersehen, dass es sich lediglich um die graphische Darstellung von Merkmalsvergleichen handelt und nicht um den Nachweis von Abstammung. Ohne Zweifel ist die gebotene Datenfülle eindrucksvoll, und es gibt beachtliche Erkenntnisse über abweichende Pflanzengruppen, die in solchen „Stammbäumen“ an die Basis gestellt werden. Hierzu zählen z. B. die Seerosen und ihre Verwandten und der viel zitierte, nur in Neukaledonien vorkommende Strauch Amborella. Was aber diese Abweichungen wirklich bedeuten, ist eine offene Frage.
Eine Zeitlang galt eine fossile Pflanze mit Namen Archaefructus als möglicher Kandidat einer frühen Angiosperme. Dann aber stellte sich heraus, dass nicht nur das angenommene Alter von ca. 144 Millionen Jahren zu hoch gegriffen war, sondern auch die Blütenreste falsch interpretiert worden waren, so dass jetzt auch dieser Hoffnungsträger ausscheidet.
So bleibt die Hoffnung, dass entwicklungsbiologische Untersuchungen aus der neuen Forschungsrichtung „Evo-Devo“ die Wissenschaft vielleicht der Problemlösung näher bringen. Allerdings ergaben erste molekulargenetische Untersuchungen zur Frage der Blütenentwicklung das interessante Ergebnis, dass die wesentlichen Aspekte der Blütenentwicklung seit den frühesten Linien konserviert sind. Daher führt auch dieser Befund bei der Suche nach ursprünglichen Formen nicht weiter.
Somit ist also Darwins „abscheuliches Geheimnis“ einer Lösung immer noch nicht näher gekommen, und es ist daher umso erstaunlicher, dass namhafte Wissenschaftler davon sprechen, dass sie eine Lösung des „abominable mistery“ in Reichweite sehen.
Literaturhinweise
Wer sich für nähere Einzelheiten interessiert, sei auf folgende Publikationen hingewiesen:
Kutzelnigg H (2000, 2001): Das „abscheuliche Geheimnis“. Woher kommen die Angiospermen? Stud. Int. J. 7, 51-58; 8, 10-15.
Kutzelnigg H (2008) Neues zum explosionsartigen Auftreten der Bedecktsamer. Stud. Int. J. 15, 88-92.
Pennisi E (2009) On the origin of flowering plants. Science 324, 28-31.
Autor dieser News: Herfried Kutzelnigg
© 2009, http://www.genesisnet.info/schoepfung_evolution/n131.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.03.09 Von Schildkröten, Makroevolution und Kreationismus
Im Newsbeitrag „Schildkröten-Übergangsform?“, https://www.genesisnet.info/index.php?News=121 wurde über einen Schildkröten-Fossilfund berichtet, der als evolutionäre Übergangsform diskutiert wird. In einem Beitrag zum Thema „Kreationismus und ‘Intelligent Design’“ im jüngst erschienenen Band „Evolution. Der Fluss des Lebens“ des Staatlichen Museums für Naturkunde Stuttgart geht Hansjörg Hemminger auf diesen Fund ein. Seine Ausführungen sollen hier kurz kommentiert werden.
Hemminger thematisiert nur die Deutung des neuen Fundes Odontochelys semitestacea als evolutive Übergangsform (vgl. Li et al. 2008). In derselben Nature-Ausgabe erschien jedoch auch eine Kritik dieser Deutung von Reisz & Head (2008, 451), wonach es sich bei Odontochelys nicht um eine primitive Form, sondern im Gegenteil um eine an das Leben im Wasser spezialisierte Anpassung handeln könnte, bei der sekundär der Rückenpanzer verloren ging. Da es sich bei Odontochelys um das älteste bekannte Schildkrötenfossil handelt, würde sich bei dieser Deutung die Frage nach der Entstehung der Schildkröten verschärfen. Im oben genannten Newsbeitrag wird diese Frage offen gelassen, da sie anhand der vorliegenden Daten offenbar nicht entschieden werden kann. Von der Kritik der Deutung als Übergangsform ist bei Hemminger nichts zu lesen, er präsentiert die wissenschaftliche Diskussion also selektiv und begeht damit eine Unterlassung, die er selber oft den Kreationisten vorwirft. Er schreibt dann ausgerechnet, dass der Fall „bezeichnend“ sei „für die Art, wie der Kreationismus argumentiert“, und behauptet: „Lücken im Fossilbericht werden dazu genutzt, riesige und durch die Evolutionstheorie scheinbar nicht erklärbare Entwicklungssprünge festzustellen.“ Tatsächlich ist von „nicht erklärbar“ im Text der Studiengemeinschaft Wort und Wissen, auf den sich Hemminger hier ausdrücklich bezieht, nirgends die Rede, sondern nur von nicht erklärt – ein großer Unterschied! Das gilt auch für andere Texte, insbesondere auch für „Evolution – ein kritisches Lehrbuch“. Und die Kennzeichnung „riesige“ ist eine Übertreibung der Aussagen von Wort und Wissen. Die Lücken und die Erklärungsprobleme werden im Falle der Schildkröten auch von Evolutionsbiologen thematisiert. Wenn Funde gelingen, die diese Lücken teilweise schließen oder schließen könnten, wurde das bei Wort und Wissen häufig thematisiert, z. B. auf dem Gebiet des Übergangs von Fischen zu Vierbeinern (s. z. B. „Entstehung der Vierbeiner“, https://www.genesisnet.info/index.php?Artikel=42843&Sprache=de&l=1 und „Tiktaalik – ein erstklassiges Bindeglied?“, https://www.genesisnet.info/index.php?News=63 oder die Monographien der Reihe STUDIUM INTEGRALE, https://www.wort-und-wissen.org/shop/, und viele Beiträge in Studium Integrale Journal, https://www.si-journal.de/). Was ist hier also „bezeichnend“? Nur durch eine selektive Auswahl von Detailaussagen und ihre verzerrte Wiedergabe kann Hemminger seine Argumentation plausibel durchhalten und Wort und Wissen auch noch „Propaganda gegen die Naturwissenschaft“ vorwerfen (S. 177).
Weiter ist für Hemminger „unerfindlich“, warum es sich beim Übergang zu den Schildkröten um Makroevolution handeln solle. Er zitiert in diesem Zusammenhang aus dem Genesisnet-Artikel „Evo-Devo“, https://www.genesisnet.info/pdfs/Evo-Devo.pdf, leider ohne Quellenangabe, so dass es dem Leser nicht möglich ist, das Zitat im Zusammenhang zu lesen. Dort findet sich auch eine Begründung, warum die Entstehung der Schildkröten einen makroevolutiven Schritt fordert und ebenso, warum es für diesen Schritt keine evolutionstheoretische Erklärung gibt. Hemminger teilt seinen Lesern diese Begründung nicht mit, obwohl sie im selben Artikel zu finden ist wie das Zitat, das er anführt. So schreiben Gilbert et al. (2001, 47): „Insgesamt enthält der Panzer über 50 Hautknochen, die bei keiner anderen Wirbeltier-Ordnung bekannt sind, und die Anwesenheit dieser Knochenhülle erforderte aufwändige Veränderungen des Vierfüßerbauplans. … Der Hals, der Schädel und der Kopulationsapparat sind ebenfalls in hohem Maße verändert.“ Die Neuorganisation des Bauplans betrifft auch Atmung und Fortbewegung „radikal“, und sie ist von allen anderen Wirbeltieren verschieden (Burke 1989, 364). Kein Wunder also, dass Rieppel (2001, 987) feststellt: „Die Evolution der stark abgeleiteten adulten (= erwachsenen) Anatomie der Schildkröten ist ein Paradebeispiel eines makroevolutiven Ereignisses, das durch Veränderungen in der frühen Embryonalentwicklung ausgelöst wurde.“ Ob Änderungen in der Ontogenese (= Individualentwicklung) makroevolutive Erklärungen wirklich ermöglichen, wird im Evo-Devo-Artikel (https://www.genesisnet.info/pdfs/Evo-Devo.pdf) diskutiert; das soll hier nicht wiederholt werden.
Angesichts dieser Tatsachen ist die Behauptung Hemmingers, dass Funde wie Odontochelys die Lücken schließen und die Evolutionstheorie bestätigen, nicht gerechtfertigt. Unabhängig von der Tatsache, dass es „die Evolutionstheorie“ nicht gibt, beruhen seine Darstellungen hier nachweislich auf einem selektiven, oberflächlichen und unkritischen Umgang mit den vorliegenden Befunden und Argumenten. Leider ist das kein Einzelfall; vielmehr scheint diese Art der Behandlung von Fachproblemen bei Hemminger System zu haben, wenn es ihm darum geht, „kreationistische“ Positionen z. B. als „Calvinball“ zu diskreditieren (z. B. EZW-Text 195, „Mit der Bibel gegen die Evolution“; siehe dazu dessen Besprechung unter https://www.wort-und-wissen.org/disk/d08-2/).
Literatur
Burke AC (1989) Development of the Turtle Carapace: Implications for the Evolution of a Novel Bauplan. J. Morphol. 199, 363-378.
Gilbert SF, Loredo GA, Brukmann A & Burke AC (2001) Morphogenesis of the turtle shell: the development of a novel structure in tetrapod evolution. Evol. Dev. 3, 47-58.
Hemminger H (2009) Kreationismus und „Intelligent Design“. In: Schmid U & Bechkly G (Hg) Evolution. Der Fluss des Lebens. Stuttgarter Beiträge zur Naturkunde _Serie C 66/67, S. 173-179.
Li C, Wu XC, Rieppel O, Wang LT & Zhao LJ (2008) An ancestral turtle from the Late Triassic of southwestern China. Nature 456, 497-501.
Reisz RR & Head JJ (2008) Palaeontology: Turtle origins out to sea. Nature 456, 450-451.
Rieppel O (2001) Turtles as hopeful monsters. BioEssays 23, 987-991.
Originalzitate
„Altogether, the shell contains over 50 dermal bones found in no other vertebrate order, and the presence of this bony casing has necessitated extensive modifications of the tetrapod body plan. … The neck, skull, and copulatory apparatus are also greatly modified“ (Gilbert et al.).
„The evolution of the highly derived adult anatomy of turtles is a prime example of a macroevolutionary event triggered by changes in early embryonic development“ (Rieppel).
Autor dieser News: Reinhard Junker
© 2009, http://www.genesisnet.info/schoepfung_evolution/n126.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.12.08 Schildkröten-Übergangsform?
Schildkröten (Testudines) sind faszinierende Tiere. Ihr ungewöhnlicher Bauplan ist sehr verschieden von den Bauplänen anderer Echsen. Der einzigartige Panzer ist aus zahlreichen Knochenplatten zusammengesetzt, die von ca. 50 Hornschilden oder lederartiger Haut bedeckt sind. Er entsteht durch Verschmelzung von Wirbelsäule, Rippen, Teilen des Schultergürtels und spezialisierten Hautknochen. Rückenpanzer (Carapax) und Brustpanzer (Plastron) sind durch eine Knochen- oder Knorpelbrücke miteinander verbunden. Der eigenartige Bau erfordert entsprechende bauliche und funktionale Abstimmungen mit anderen Organen, zum Beispiel bei der Atmung und Fortbewegung. Einzigartig ist auch die Lage des Schultergürtels, der sich innerhalb der Rippen entwickelt. Der Körperbau ist so ungewöhnlich, dass nur wenige Merkmale benutzt werden können, um die Schildkröten mit irgendwelchen anderen Tiergruppen zu verbinden und daraus Hinweise auf mögliche stammesgeschichtliche Beziehungen zu entnehmen. Da auch die bislang ältesten fossilen Schildkröten der Gattung Proganochelys aus der Obertrias Deutschlands (auf 204-206 Millionen Jahre datiert) den typischen Schildkröten-Bauplan bereits besaßen (Li et al. 2008), gaben Fossilien bis vor kurzem keine Hinweise auf den Ursprung der Schildkröten.
Neuer Fund. Li et al. (2008) berichten nun von Fossilfunden einer Schildkröte, die einen Zwischenschritt in der Evolution des Panzers dokumentieren soll. Die neue Art Odontochelys semitestacea wurde in Schichten in der südwestchinesischen Provinz Guizhou in Küsten-Ablagerungen entdeckt, die mit datierten 220 Millionen Jahren noch älter sind als Proganochelys. Der Artnahme bedeutet sinngemäß „Halbpanzerschildkröte mit Zähnen“, womit zwei wesentliche Unterschiede zu heutigen Schildkröten angedeutet sind. Zum einen waren Ober- und Unterkiefer des Tieres bezahnt und hatten keine schnabelartigen Kieferleisten heutiger Schildkröten. Zum anderen war zwar der Bauchpanzer des etwa 40 cm großen Tieres voll entwickelt; der Rückenpanzer bestand aber nur aus Neuralplatten; die rückenseitigen Rippen waren lediglich verbreitert. Hautknochen („osteoderms“) waren nicht ausgebildet. Li et al. (2008) schließen daraus, dass sich der bauchseitige Panzer vor dem Rückenpanzer entwickelt hat und dass der erste Schritt der Entstehung des Rückenpanzers in der Verknöcherung von Neuralplatten und Verbreiterung der Rippen bestand. Diese Abfolge stimmt mit dem Verlauf der frühen Embryonalentwicklung der heutigen Schildkröten überein. Bisher war ein Zusammenwachsen verknöcherter Hautschuppen als erster Schritt favorisiert worden. Gegen die Deutung, es handle sich um ein jugendliches Exemplar spricht nach Li et al. (2008, 499) die Verschmelzung von Sprungbein (Astragalus) und Fersenbein (Calcaneum).
Alternative Interpretation. Der Interpretation von Li et al. stellen Reisz & Head (2008) in einem Kommentar jedoch eine andere entgegen: Lange, verbreiterte Rippen sind Bestandteile des Rückenpanzers aller Schildkröten. Deren Vorkommen bei Odontochelys sei ein Hinweis darauf, dass das embryonale Gewebe, welches die Bildung des Carapax kontrolliere, ebenfalls vorhanden gewesen sei. Ebenso sei die Verbindungsbrücke zwischen Carapax und Plastron ausgebildet, was ebenfalls für die Anwesenheit eines Carapax spreche. Zusammengenommen spreche das für die alternative Interpretation, dass ein Carapax doch ausgebildet gewesen sei, jedoch einige seiner Teile nicht verknöchert gewesen seien. Diese Interpretation von Odontochelys führe zur Möglichkeit, dass dessen Panzer nicht primitiv, sondern stattdessen eine spezialisierte Anpassung durch einen sekundären Verlust gewesen sei (Reisz & Head 2008, 451), also keine Ausprägung, die einem frühen Ontogenesestadium entspricht. Eine Reduktion der Hautknochen ist bei wasserlebenden Schildkröten verbreitet; die Deutung einer Spezialisierung passt also zum vermutlichen Lebensraum von Odontochelys im küstennahen Wasser. Diese Ausprägung könne durch ein Stehenbleiben der Entwicklung auf einem jugendlichen Stadium (sog. Paedomorphose) zustandegekommen sein.
Allerdings passt die Bezahnung, die evolutionstheoretisch als ursprüngliches Merkmal gelten muss, nicht ohne Weiteres zu dieser Deutung. Insgesamt handelt es sich also um ein Merkmalsmosaik, das nicht leicht in ein stammesgeschichtliches Schema eingeordnet werden kann. Wie in vielen anderen Fällen auch zeigt sich, dass Mosaikformen nicht mit evolutiven Übergangsformen gleichzusetzen sind.
Ursprungsfrage verschärft? Die alternative Interpretation der Befunde durch Reisz & Head würde die Frage nach dem Ursprung der Schildkröten unverändert offen lassen, wenn nicht sogar verschärfen: Denn demnach wäre ausgerechnet eine spezialisierte Form die bislang älteste. Die Funde von Odontochelys machen auch beispielhaft deutlich, wie schwierig die Interpretation fossiler Funde sein kann.
Literatur
Li C, Wu XC, Rieppel O, Wang LT & Zhao LJ (2008) An ancestral turtle from the Late Triassic of southwestern China. Nature 456, 497-501.
Reisz RR & Head JJ (2008) Palaeontology: Turtle origins out to sea. Nature 456, 450-451.
Autor dieser News: Reinhard Junker
© 2008, http://www.genesisnet.info/schoepfung_evolution/n121.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
21.04.05 Konvergenz der Gehörknöchelchen der Säugetiere
Ein neuer Fossilfund eines Kloakentiers (Untergruppe der Säugetiere) zeigt deutliche Hinweise darauf, dass die säugertypischen Gehörknöchelchen Hammer, Amboss und Steigbügel bei diesem Säuger nicht ausgebildet waren. Daraus muss gefolgert werden, dass diese Konfiguration, die als eines der wichtigsten Schlüsselmerkmale der Säugetiere betrachtet wird, mindestens zweimal unabhängig (konvergent) entstanden ist. Befunde wie diese bleiben nicht ohne Folgen für Phylogenetik (= Stammbaumforschung), wie weiter unten kurz erläutert werden soll.
Die drei Gehörknöchelchen leiten im Mittelohr der Säugetiere die Schwingungen des Trommelfells zum ovalen Fenster des Innenohrs. Nach evolutionstheoretischen Vorstellungen leiten sie sich von Knochen her, die bei Echsen das Kiefergelenk bilden. Der Bau des Säuger-Mittelohrs ist so komplex und einzigartig, dass einige Forscher die Ablösung der Mittelohrknochen vom Kiefer sogar als wichtigste Synapomorphie (abgeleitetes Merkmal) der heutigen Säugetiere betrachten (Martin & Luo 2005, 861).
Das neuerdings von Rich und Mitarbeitern (2005) beschriebene Fossil erzwingt nun aber eine neue Sicht. Die Konfiguration des Unterkieferknochens des jüngst entdeckten, bisher ältesten bekannten Kloakentiers Teinolophus trusleri (auf 115 Millionen Jahre datiert) zeigt klare Hinweise auf das Vorhandensein eines Angulare, Artikulare und Quadratum und ihre Verbindung mit dem Unterkiefer. Da gleichzeitig lebende Säugetiere den typischen Bau des Kiefergelenks und des Mittelohrs aufweisen und die heutigen Kloakentiere (mit drei Gehörknöchelchen) aus dem beschriebenen Fossil hervorgegangen sein sollen, muss aufgrund dieses Fundes geschlossen werden, dass die Mittelohrknochen zweimal unabhängig voneinander bei den Säugetieren entstanden sind – in der Gruppe der Theria (Beuteltiere und Plazentatiere) vor ca. 215-225 Millionen Jahren und etwa 100 Millionen Jahre später in der Gruppe der Kloakentiere. Bisher war man der Auffassung, dass die Auswanderung der Gehörknöchelchen aus dem Kiefergelenk vor der Aufspaltung der Kloaken-, Beutel- und Säugetiere erfolgte. Die Mittelohrknochen Hammer, Amboss und Steigbügel verlieren damit nach Rich et al. (2005) ihre Bedeutung als charakteristisches Kennzeichen der Säugetiere. Konvergenz eines Schlüsselmerkmals: das ist ein Vorgang, den Biologen bisher höchstwahrscheinlich ausgeschlossen hätten. Ist eine solche Situation evolutionstheoretisch noch plausibel deutbar?
Die Konvergenz der Gehörknöchelchen demonstriert beispielhaft die Problematik des evolutionären Homologie-Arguments. Wenn selbst einzigartige Schlüsselmerkmale mehrfach unabhängig entstehen können, ist das Homologie-Argument am Ende. Denn offenbar kann aufgrund von Bauplanähnlichkeiten nicht auf gemeinsame Abstammung geschlossen werden. Daher gibt es auch keinen Homologiebeweis der Evolution (vgl. „Ähnlichkeiten in der Morphologie und Anatomie“, https://www.genesisnet.info/index.php?Artikel=41301&Sprache=de&l=1; für eine ausführliche Diskussion dieses Sachverhalts siehe Junker 2002, 28-45). Ein verschachteltes, hierachisches System von Schlüsselmerkmalen (das enkaptische System) wurde häufig als einer der stärksten Belege für Makroevolution genannt. Dieser Beleg wird mit zunehmender Kenntnis der Formenvielfalt in Frage gestellt.
Literatur
Junker R (2002) Ähnlichkeiten, Rudimente, Atavismen. Holzgerlingen.
Martin T & Luo ZX (2005) Homoplasy in the Mammalian Ear. Science 307, 861-862.
Rich TH, Hopson JA, Musser AM, Flannery TF & Vickers-Rich P (2005) Independent Origins of Middle Ear Bones in Monotremes and Therians. Science 307, 910-914.
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n35.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/