
Paläontologie: Fossilien des Erdaltertums (z. B. Tetrapoden und Kambrische Explosion)
02.05.23 Das Monster, das in kein Schema passt: Das Tully-Monster
Es sieht aus wie ein Phantasiewesen und seine Merkmalkombination hat es in sich: Das Tully-Monster (Tullimonstrum gregarium) lässt sich bis heute nicht schlüssig in das System der Lebewesen einfügen – nach einer neuen Studie gilt dies mehr denn je. Es ist ein Paradebeispiel für eine freie Kombinierbarkeit von tierklassen- und sogar tierstammübergreifenden Merkmalen. Eine solche Freiheit besitzt allerdings nur ein Schöpfer.
Zu den vielzitierten Belegen für Evolution gehört das eingeschachtelte System der Lebewesen. Verschiedene Arten lassen sich auf der Basis von Merkmalsübereinstimmungen zu Gattungen zusammenfassen (z. B. Katzen), Gattungen zu Familien (z. B. Katzenartige), Familien zu Ordnungen (z. B. Raubtiere), Ordnungen zu Klassen (z. B. Säugetiere) und Klassen zu Stämmen (z. B. Chordatiere; vgl. Abb. 1).

Abb. 1: Enkaptisches System: Ordnung der Lebewesen in einem hierarchischen, enkaptischen (eingeschachtelten) System, am Beispiel von Riesengleithörnchen (Petaurista petaurista), Eichhörnchen (Sciurus vulgaris) und Wolf (Canis lupus). (Nach Peters & Gutmann 1971)
Ein eingeschachteltes System lässt sich problemlos in eine Baumdarstellung übersetzen, die sich evolutionstheoretisch wiederum im Sinne der gemeinsamen Abstammung aller Lebewesen interpretieren lässt. Bei manchen Tiergruppen funktioniert das tatsächlich recht gut. Aber mit zunehmenden Kenntnissen über die Verteilung von Merkmalen bei verschiedenen Arten und höheren Taxa (umfassenderen Gruppen) wuchs die Anzahl von Merkmalswidersprüchen, insbesondere unter fossil bekannten Formen, aber auch bei heute lebenden Gruppen. In solchen Fällen erweisen sich netzartige Darstellungen von Merkmalsbeziehungen als deutlich widerspruchsärmer als die allgemein üblichen Baumdiagramme. Das aber widerspricht dem genannten Evolutionsbeweis und evolutionstheoretischen Vorhersagen. Die Folge ist aber in der Regel nicht, dass deshalb Evolution als Rahmentheorie in Frage gestellt wird. Vielmehr sucht man nach neuen, unbekannten Mechanismen, die zur unerwarteten Merkmalsverteilung geführt haben sollen. Evolution als Rahmentheorie steht nicht zur Disposition, eine Falsifizierungsmöglichkeit wird gar nicht erst in Betracht gezogen. Nicht zu vermeiden ist aber die Schlussfolgerung, dass damit ein wissenschaftlicher Beleg für Evolution – das eingeschachtelte System – geschwächt wird.
Tullimonstrum gregarium – das „Tully-Monster“
Ein extremes Beispiel dafür, dass die Merkmalskombination einer Art in kein evolutionäres Schema passt, ist das Tully-Monster (Tullimonstrum gregarium). Seinen Namen hat dieses nur fossil erhaltene Tier nicht umsonst. Man könnte es leicht für ein Phantasiewesen halten, wenn man eine der zahlreichen Rekonstruktionen betrachtet (s. Abb. 2).

Abb. 2: Rekonstruktion von Tullimonstrum. (Nobu Tamura, CC BY-SA 4,0,
https://creativecommons.org/licenses/by-sa/4.0/deed.en)
Tatsächlich aber wurden Tausende von Exemplaren entdeckt – ausschließlich in der oberkarbonischen Mazon-Creek-Formation (Illinois/USA; auf ca. 290 Millionen radiometrische Jahre datiert). Das Tier hatte einen spindelförmigen Körper und war ca. 30–35 cm lang, manche Individuen waren aber deutlich kleiner. Es besaß ein Paar vertikaler Bauchflossen am Schwanzende seines Körpers, einen langen Rüssel (Proboscis), der in einer Art Maul endete, das bis zu je acht kleine scharfe Zähne an jedem „Kiefer“ trug, was auf eine räuberische Lebensweise hinweist. Sehr ungewöhnlich sind auch die dünn gestielten Augen. Entlang des Körpers ist eine Segmentierung zu erkennen – auch in der Kopfregion vor den Augen, was ebenfalls ungewöhnlich ist. Hartteile wurden nicht nachgewiesen.
Es wundert nicht, dass die Einordnung des im Jahr 1966 erstmals beschriebenen Tieres ins System der Tiere Probleme bereitet und bis heute kontrovers diskutiert wird. Sein Körperbau unterscheidet sich von allen anderen bekannten Tierbauplänen. Tullimonstrum wurde mit Schnecken (Gastropoda), mit der ebenfalls ungewöhnlichen kambrischen Gattung Opabinia mit unklarer systematischer Stellung, mit fossilen Conodonten („Kegelzähner“), Vielborstern (Polychaeta-Würmer), Schnurwürmern (Nemertea) und Nektocariden (Gruppe der Kopffüßer) verglichen (Mikami et al. 2023, 1).
Ein Wissenschaftlerteam glaubte, eine Chorda (langer, dünner und biegsamer Stab im Rückenbereich), ein dreiteiliges Gehirn, Kiementaschen, Muskelsegmente wie bei Neunaugen und anderen Chordatieren sowie Zähne, die denen von Neunaugen und Schleimaalen ähneln, nachgewiesen zu haben (McCoy et al. 2016). Dieses Team schlug vor, Tullimonstrum als ursprüngliches Wirbeltier zu klassifizieren. Daran wurde postwendend Kritik geäußert (Sallan et al. 2017). Wichtige wirbeltiertypische Merkmale wie Ohrkapseln und Körperpigmentierung würden fehlen; es sei bekannt, dass viele Merkmale konvergent (evolutionär unabhängig) auftreten und daher phylogenetisch (in Bezug auf die Stammesgeschichte) nicht aussagekräftig seien. Durch eine chemische Analyse der Fossilien wurden dann allerdings Überreste von Proteinen gefunden, aus denen das Keratin und Kollagen von Wirbeltieren besteht (McCoy et al. 2020), was die Forscher wiederum als Hinweis auf eine Wirbeltierverwandtschaft werteten.
Eine aktuelle Untersuchung von Mikami et al. (2023) erbrachte nun neue Befunde, die eine Entscheidung in dieser Kontroverse herbeiführen könnten. Mikami und sein Team analysierten 153 fossile Exemplare des Tully-Monsters mithilfe eines hochauflösenden 3D-Laserscanners und mittels Mikro-Röntgentomografie. Dabei stellte sich heraus, dass sich die von McCoy et al. (2016) beschriebenen, vermeintlich wirbeltierähnlichen Merkmale (dreigeteiltes Gehirn, segmentierte Muskeln und Strahlenflossen) deutlich von Wirbeltieren unterscheiden. „Unsere Analysen […] deuten auf das Vorhandensein von Segmenten in der präoptischen Region und das Fehlen von Tektalknorpeln [Knorpel im Bereich des Gehirns], wirbeltierähnlichen Gehirnen, Kiemendeckeln und Flossenstrahlen hin, was Zweifel an der Wirbeltierzugehörigkeit von Tullimonstrum aufkommen lässt“ (Mikami et al. 2023, 7). Die vordere Körperregion zeigte auch im Kopf und im langen Mundfortsatz (Proboscis) eine Segmentierung, was untypisch für Wirbeltiere, dagegen typisch für Wirbellose ist. Zudem unterscheiden sich die schmalen, scharfen Zähne des Rüssels von den Hornzähnen von Neunaugen und Schleimaalen, zu deren Verwandtschaft Tullimonstrum von McCoy et al. (2016) gestellt worden waren. Die Forscher kommen zum Schluss, dass Tullimonstrum weder ein Wirbeltier war noch mit ihnen enger verwandt ist. Stattdessen schlagen sie vor, Tullimonstrum entweder zu den nicht-wirbeltierartigen Chordatieren oder zu den Urmundtieren (Protostomier) zu stellen. In letzterem Fall müsste man allerdings einen radikal veränderten Bauplan mit einzigartigen Merkmalen annehmen (Mikami et al. 2023, 10) und damit eine isolierte Stellung im System der Lebewesen.
Die Autoren diskutieren die Merkmalsbeziehungen von Tullimonstrum zu unterschiedlichen Tiergruppen. Der Einordnung als Stammchordat mit Segmentierung auch im Kopfbereich vor den Augen steht allerdings das Fehlen eines Nachweises einer mit den Pharyngealbögen (Schlundbögen) verbundenen Struktur in Tullimonstrum entgegen. Tullimonstrum könnte diese Struktur allenfalls sekundär verloren haben (Mikami et al. 2023, 10).
Die Autoren halten auch eine Nähe von Tullimonstrum zum weiteren Verwandtschaftsbereich der Gliederfüßer (Panarthropoden) für unwahrscheinlich, denn die Körpersegmentierung ist anders als die sklerotisierten (ausgehärteten) Segmente der Gliederfüßer-Fossilien aus Mazon Creek. Die Körpersegmentierung von Tullimonstrum unterscheidet sich auch von der eines untersuchten Stummelfüßers (Onychophora) aus Mazon Creek. Eine nähere Verwandtschaft von Tullimonstrum komme daher nur außerhalb der Panarthropoda in Frage. Doch auch eine Verwandtschaft mit Ringelwürmern (Anneliden), Weichtieren (Mollusken) und Schnurwürmern (Nemertea) sei unwahrscheinlich, da ihnen eine vergleichbare Körpersegmentierung wie Tullimonstrum fehlt. Zwar besitzen Anneliden und Nemerteen sich wiederholende Strukturen im Rumpf, sie erstecken sich aber nicht wie bei Tullimonstrum auf die Kopfregion. Außerdem unterscheiden sich die Rüssel von Anneliden und Nemerteen von denen von Tullimonstrum dadurch, dass sie einklappbar oder einziehbar sind. Darüber hinaus haben Mollusken im Allgemeinen keine segmentale Wiederholung in ihren Körpern – und soweit das doch der Fall ist (bei den Polyplacophora und Monoplacophora), sind sie nicht mit denen von Tullimonstrum vergleichbar, da ihnen klare Grenzen fehlen (Mikami et al. 2023, 11).
Fazit
Es bleibt nach Mikami et al. (2023, 12) dabei: Der einzigartige Körperbau von Tullimonstrum ist kaum mit dem eines anderen bekannten Tieres vergleichbar. Eine eindeutige systematische bzw. phylogenetische Zuordnung des Tully-Monsters ist weiterhin unklar. Die neuen Untersuchungen, die bisher nicht bekannte Details sichtbar gemacht haben, unterstreichen die Einzigartigkeit von Tullimonstrum. Die Merkmalskombination dieses merkwürdigen Tieres ist ein Paradebeispiel für eine freie Kombinierbarkeit von tierklassen- und sogar tierstammübergreifenden Merkmalen. Eine solche Freiheit besitzt aber nur ein Schöpfer.
Quellen
Mikami T, Ikeda T et al. (2023) Three-dimensional anatomy of the Tully monster casts doubt on its presumed vertebrate affinities. Palaeontology 62, e12646; doi:10.1111/pala.12646.
McCoy VE, Saupe EE et a. (2016) The ‘Tully monster’ is a vertebrate. Nature 532, 496–499.
Sallan L, Giles S et al. (2017) The ‘Tully Monster’ is not a vertebrate: characters, convergence and taphonomy in Palaeozoic problematic animals. Palaeontology 60, 149–157.
McCoy VE, Wiemann J et al. (2020) Chemical signatures of soft tissues distinguish between vertebrates and invertebrates from the Carboniferous Mazon Creek Lagerstätte of Illinois. Gebiology 18, 560–565.
Autor dieser News: Reinhard Junker
© 2023, http://www.genesisnet.info/schoepfung_evolution/n313.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
16.03.21 Lungenfische – Genome und ihre Interpretation
Lungenfische sind – darauf weist bereits ihr Name hin – Fische mit ungewöhnlichen Eigenschaften, z. B. der Fähigkeit, mit lungenähnlichen Organen Luft zu atmen. Ihr bisher unzugängliches Erbgut wurde jetzt von zwei verschiedenen Arbeitsgruppen an zwei verschiedenen Arten mit Hilfe modernster Sequenzierungsmethoden analysiert. Die Autoren interpretieren ihre Resultate in auffälliger Weise als Bestätigung etablierter Evolutionsvorstellungen für den Übergang vom Leben im Wasser zum Festland. Hier werden die Ergebnisse vorgestellt und ihre Interpretationen kritisch gewürdigt.
Lungenfische und ihre vermutete Bedeutung in der Stammesgeschichte (Phylogenese)
Die Lungenfische (Dipnoi)1 weisen neben den für Fische typischen Kiemen, die ihnen die Sauerstoffaufnahme aus dem Wasser ermöglichen, vergleichsweise einfach gebaute Lungen auf. Diese erlauben ihnen z. B. bei extremer Sauerstoffarmut im Wasser, auch an der Wasseroberfläche zu atmen. Fossil sind Lungenfische in erstaunlicher Vielfalt aus Süß- und Meerwasser seit dem Devon (Paläozoikum) überliefert. Derzeit sind verschiedene heute lebende Lungenfischarten aus Südamerika (Lepidosiren paradoxa), Afrika (Protopterus aethiopicus, P. amphibius, P. annectens und P. dolloi) und Australien (Neoceratodus forsteri) bekannt. Sie werden als träge Tiere beschrieben, die in stehenden oder langsam fließenden Gewässern andere Wasserlebewesen erbeuten und als Nahrung nutzen. Der Südamerikanische und die Afrikanischen Lungenfische können in Trockenzeiten, wenn der Wasserspiegel ihrer Gewässer sinkt und diese evtl. sogar ganz austrocknen, röhrenförmige Löcher in den Untergrund graben. Mit dem Kopf zur Öffnung begeben sie sich dort hinein, und wenn der Wasserspiegel weiter sinkt, scheiden sie große Mengen an Schleim aus, der fest wird und sie wie ein Kokon in der Röhre umgibt; so können sie sich vor dem kompletten Austrocknen schützen. Diese Lungenfische können durch kleine Öffnungen in ihrem Schleimkokon Luft atmen und so Wochen und Monate, ja sogar bis zu einigen Jahren überleben.
Lungenfische werden taxonomisch der Klasse der Fleischflosser (Sarcopterygii) zugeordnet (wie auch die Quastenflosser, Coelacanthiformes). Das für diese Zuordnung maßgebliche Merkmal sind die fleischigen Flossen, die an den paarigen Flossen (bei den Quastenflossern auch bei der 2. Rücken- und Afterflosse) Skelettstrukturen aufweisen, die bei Landwirbeltieren dem Oberarm- (Humerus) bzw. dem Oberschenkelknochen (Femur) entsprechen und durch diese Beziehung als homolog bezeichnet werden. In der Systematik und vergleichenden Anatomie wird Homologie bereits mit evolutionären Vorstellungen verknüpft. Das heißt: Die Homologie ähnlicher Strukturen wird auf vermutete evolutionäre Zusammenhänge zurückgeführt. So wird die mutmaßliche Homologie der Skelettstrukturen in den Flossen der Lungenfische mit derjenigen der Extremitäten von Tetrapoden (= Vierbeiner) in evolutionären Betrachtungsweisen als bedeutungsvoller Hinweis für einen Übergang vom Wasserlebewesen zur vierbeinigen Fortbewegung an Land bewertet. Da nun Lungenfische darüber hinaus auch noch einfach gebaute Lungen besitzen, ist leicht nachzuvollziehen, dass diesen Organismen in evolutionärer Perspektive eine bedeutende Schlüsselstellung bei der Eroberung des Festlandes zugesprochen wird.
Das Erbgut der Lungenfische
Der molekularbiologischen Erforschung dieses evolutionären Übergangs stand bisher die schiere Größe des Genoms (= gesamten Erbgutes) der Lungenfische entgegen. In bisherigen Untersuchungen war gezeigt worden, dass das Erbgut der Lungenfische einen riesigen Umfang hat und mehrfach so groß ist wie das Humangenom. Mit bisherig verfügbaren Methoden war es deshalb nicht analysierbar. Jetzt haben ein internationales und ein chinesisches Forscherteam das Genom des Australischen Lungenfisches (Neoceratodus forsteri) bzw. das eines afrikanischen Lungenfisches (Protopterus annectens) veröffentlicht. Sowohl Meyer et al. (2021) als auch Wang et al. (2021) heben bereits in den Titelformulierungen ihrer Veröffentlichung die Bedeutung des von ihnen ermittelten Genoms für den Übergang der Wirbeltiere vom Wasser ans Land hervor. In beiden Projekten wurde zur Analyse des Erbguts von Lungenfischen modernste Sequenzierungstechnologie eingesetzt wie z. B. die Nanopore-Sequenzierung. Dabei wird an einer technischen Membran mit einer minimalen Öffnung (Nanopore) ein Potenzial angelegt, das einen konstanten Ionenstrom durch die Membran bewirkt. Vor der Pore wird der zu analysierende DNA-Doppelstrang durch eine dort fixierte Helicase aufgetrennt und ein Einzelstrang in die Pore eingeführt. Jede der vier DNA-Basen verändert den Ionenstrom spezifisch und so kann mit dieser Technologie während der DNA-Strang durch die Pore gefädelt wird, dessen Sequenz aufgrund der sich ändernden Stromsignale bestimmt werden. Mit dieser Technik können sehr lange DNA-Stränge sequenziert werden. Beide Autorenteams präsentieren ein Genom der Lungenfische von bisher nicht sequenziertem Umfang von 43 Gb (Gigabasen) (Meyer et al.) und 40 Gb (Wang et al.), was etwa dem 13-Fachen der Menge an Basen im Humangenom (ca. 3,2 x 109) entspricht. Das größte bisher sequenzierte Genom war das des Axolotls (Ambystoma mexicanum) mit 32 Gb.
Das Genom von Neoceratodus forsteri
In der Arbeit über das Genom des Australischen Lungenfischs (N. forsteri) beschreiben Meyer et al. zunächst die Sequenzierungsarbeit und die Zusammenstellung (assembly) des Genoms. Mit den Sequenzbereichen, die 30 x gelesen worden waren (30 x coverage), konnten sie 37 Gb und damit 88,2 % des Genoms aufklären. Der Verteilung des Genoms auf die 17 Makro- und 10 Mikrochromosomen widmeten Meyer et al. viel Aufmerksamkeit und verglichen die Resultate mit denjenigen von Knochenhecht (Lepisosteus oculatus), Küken (Gallus gallus) und einer Eidechse (Rotkehlanolis, Anolis carolinensis). Die Resultate werden dahingehend interpretiert, dass die Mikrochromosomen bereits auf die ersten Wirbeltiere zurückgehen, wohingegen die Makrochromosomen sich im Verlauf des unterstellten Evolutionsgeschehens stark vergrößert haben. Im Genom finden sich Hinweise darauf, dass dieser Größenzuwachs vor allem auf dem Einbau von großen DNA-Abschnitten basiert, sogenannten LINEs (Long Interspersed Nuclear Elements) zurückgeht. Diese enthalten Gene, die es den Abschnitten erlauben, sich im Genom selbstständig an verschiedenen Stellen einzubauen (Transposons). Die Autoren vermuten eine ausgedehnte Wirksamkeit eines Kopier- und Einfüge- (copy-and-paste) Mechanismus im Verlauf der Stammesgeschichte.
Nach diesen eher allgemeinen und grundsätzlichen Untersuchungen widmen sich Meyer et al. in ihrer Studie intensiv speziellen Aspekten der mutmaßlichen Evolutionsverlaufs, nämlich der Lungenatmung, dem Geruchssinn und der Entwicklung von Gliedmaßen. Aus den Daten geht hervor, dass die Zahl der Gene, die für lungenspezifische Oberflächenproteine kodieren, in der typischen Größenordnung für Tetrapoden (vierbeinige Landwirbeltiere) und im Vergleich zu Knorpel- und Knochenfischen zwei- bis dreimal höher sind. Ein Gen zur Steuerung der Lungenentwicklung (shh) wird während der Embryonalentwicklung von N. forsteri im Gewebe der sich entwickelnden, einflügeligen Lunge sehr stark ausgeprägt. Die Autoren bewerten dies als notwendige Entwicklung zur Eroberung des Festlandes. Gene für Rezeptoren von in der Luft enthaltenen Geruchsstoffen gibt es in Fischen nur wenige, in Tetrapoden dagegen sind sie vielfältig. Auch die Anzahl dieser Genklasse ist im Vergleich zu anderen Fischen beträchtlich erhöht, was aus Sicht der Autoren wieder den Übergang vom Wasser ans Land anbahnt und begünstigt.
Hinsichtlich der für Tetrapoden typischen Gliedmaßen weisen Meyer et al. auf das erstmalige Auftreten von 31 dafür typischen Verstärker-Genen (Enhancer) in Fleischflossern hin. Einige davon werden in Embryonen von Lungenfisch in einem Muster ausgeprägt, das mit dem in Tetrapoden vergleichbar ist. Auch von den vier in N. forsteri vorkommenden Hox-Gencluster (Steuer- und Regulationsgene) werden einzelne (z. B. hoxc13) in auffälliger Weise am äußeren Ende der paarigen Flossen ausgeprägt, was dann in den Worten von Meyer et al. von den Tetrapoden für die Ausbildung von Nägeln, Hufen und Klauen übernommen wurde (man spricht in solchen Fällen von Kooption). Die Autoren verwenden interessanterweise bei der Interpretation ihrer Resultate die Formulierung, dass es sich bei den jeweils genannten genetischen Besonderheiten um „genomische Vor-Anpassungen im Lungenfisch für den Übergang der Wirbeltiere vom Wasser ans Land“2 handle. Die Autoren bemerken abschließend, dass diese Neuheiten die Fleischflosser auf die Eroberung des Festlandes vorbereitet hätten. Dies zeige, wie die Kenntnis des Genoms des Lungenfischs dazu beitragen kann, diesen Hauptübergang in der Evolution der Wirbeltiere besser zu verstehen.
Das Genom von Protopterus annectens
Wang et al. haben das Erbgut eines der Afrikanischen Lungenfischen (Protopterus annectens) sequenziert und auf chromosomaler Ebene ein Genom mit ca. 40 Gb in hoher Qualität (50 x coverage) und mit nur wenigen Lücken veröffentlicht (Wang et al. 2021). Sie konnten DNA-Stränge von 862 Megabasen (Mb) bis 5,28 Gb auf 17 Chromosomen bestimmen und 12 640 Fragmente, die nicht lokalisiert werden konnten, mit 126 Mb. Auch die Studie an diesem Lungenfischgenom ergab, dass die enorme Größe des Genoms wohl vor allem auf Transposons zurückzuführen ist. Die Autoren bezeichnen 15 Gb des Genoms als funktionslos (aufgrund von mutmaßlichen Degenerationsphasen in der Evolution; Wang et al. zitieren hier eine andere Arbeit, in der diese Befunde als „Friedhof der transponierbaren Elemente“3 bezeichnet worden war). Als auffällig wird in der Veröffentlichung auch vermerkt, dass das Erbgut des Lungenfischs sehr lange Gene (18 Mb) einhält. Während das Erbgut des Menschen nur 91 Gene enthält, die umfangreicher als 1 Mb sind enthält das Erbgut des Lungenfisches davon mehr als 5 000. Da auch die langen Gene mit vergleichbarer Effektivität wie die kleinen Gene ausgeprägt werden, muss dieser Prozess im Lungenfisch sehr fein austariert sein; da der Aufwand, umfangreiche Gene korrekt und zuverlässig zu nutzen, deutlich höher ist.

Abb. 1: Afrikanischer Lungenfisch (Protopterus annectens). (Wikimedia: Vassil, CC0)
Auch Wang et al. legen bei ihrer Studie über das Erbgut des Afrikanischen Lungenfisches (P. annectens) das Hauptaugenmerk auf stammesgeschichtliche Aspekte. Im Blick auf die namengebende Lunge finden die Autoren vier Gene, die im Lungengewebe von P. annectens wie auch im Krallenfrosch und der Maus stark ausgeprägt werden, dagegen nicht in der Schwimmblase von Flössel- und Alligatorhecht. Diese vier Gene stehen im Zusammenhang mit Oberflächenkomponenten der Lunge; sie sind an der Steuerung und Regulation der Eigenschaften der Lungenoberfläche beteiligt. Wang et al. stellen sich die Evolution der Lungenatmung aufgrund ihrer Ergebnisse wie folgt vor: Die ersten Ansätze zur Luftatmung sind bereits beim Vorläufer der Knochenfische angelegt, denn bei allen ist ein spezielles Gen (Sftpc) bereits vorhanden. Der zweite Schritt erfolgt durch weitere in der Lunge wirksame Gene, die bei den Fleischflossern auftreten, und im dritten Schritt erlangen dann die Tetrapoden durch weitere Gene eine leistungsfähige Lunge. Auch die Entwicklung zur Fünffingrigkeit der Gliedmaßen und der Übergang von Flossen zu den Gliedmaßen der Tetrapoden bringen Wang et al. mit dem Expressionsmuster von Hox-Genen in Verbindung. Verschiedene Hoxa– und Hoxd-Gene werden bei Tetrapoden ausschließlich in den Gliedmaßen ausgeprägt, während sie bei der Entwicklung der Flossen nach einem zeitlichen Muster an unterschiedlichen Orten ausgeprägt werden. Auch anhand anderer Genabschnitte, die nicht für Proteine kodieren und in verschiedenen Organismen dieselbe Sequenz aufweisen, was in evolutionärer Perspektive als „konserviert“ interpretiert wird, lassen sich ähnliche Zusammenhänge aufzeigen. Die Autoren werten das als weiteren Beleg für ihre Sicht der großen Zusammenhänge; sie sind aufgrund ihrer genetischen Analysen davon überzeugt, dass der Übergang von Flossen zu Gliedmaßen in drei Schritten verlaufen sei, diese sollen von proximal nach distal verlaufen sein, d. h. zunächst sollen die Gliedmaßenabschnitte in unmittelbarer Körpernähe verändert worden sein und danach die weiter vom Körperstamm entfernten Abschnitte.
Auch in Bezug auf die Innervierung der Gliedmaßen führen Wang et al. Hox-Gene an, die vor allem für die Motoneuronen bedeutend erscheinen, also die Nerven, durch die die Muskeln angeregt werden können. Diese Hox-Gene waren bei den Fleischflossern nachweisbar, nicht jedoch bei anderen Fischen. Zwei neue Gene, die für Neurotransmitterproteine kodieren, treten bei Fleischflossern auf und werden in deren Gehirn und Rückenmark ausgeprägt. Bei Tetrapoden werden diese Neuroproteine im Zusammenhang mit Stressmanagement und Angstauflösung diskutiert; und so passt dies aus Sicht der Autoren zur Stellung, die sie den Lungenfischen bei der Eroberung des Festlandes zugedacht haben.
Zusammenfassend skizzieren Wang et al. ihre Vorstellung vom Übergang der Lebewesen vom Wasser ans Land in drei Stufen: der Vorläufer der Knochenfische entwickelte die ersten Anlagen zur Luftatmung, in einem zweiten Schritt konnten Lungenfische und Tetrapoden durch weitere Anpassungen das Wasser zeitweise verlassen, um dann in einem dritten Schritt Atmung und Fortbewegung für ein Leben an Land zu verbessern.
Dasselbe chinesische Autorenteam hat in derselben Ausgabe des Journals Cell „die genetischen Spuren des Landgangs“4 noch weiter bis zu den Strahlenfischen zurückverfolgt (Bi et al. 2021). Sie konzentrieren sich dabei auf den Geruchssinn, die Brustflossen, die Lunge und das Herz. In dieser Studie werden vier neue Genome von Strahlenflossern beschrieben, nämlich von Flösselhecht (Polypterus senegalus), Löffelstör (Polyodon spathula), Kahlhecht (Amia calva) und Alligatorhecht (Atractosteus spatula). Die Resultate der vergleichenden und bioinformatischen Genanalysen werden nach demselben Muster wie oben präsentiert und die Argumentation verläuft analog. Die Autoren behaupten aufgrund von vergleichenden Analysen des Transkriptoms verschiedener Organgewebe von Vertretern von Fleischflossern, Strahlenflossern und Knochenfischen, dass dadurch Darwins Hypothese bestätigt würde, dass Schwimmblase und Lunge homologe Organe seien. Nun lässt sich die Auffälligkeit, dass viele gleichartige Gene im Gewebe von Schwimmblase und Lunge ausgeprägt werden auch gut durch deren ähnliche Funktion erklären. Tatsumi et al. (2017) sehen das differenzierter und erklären, dass es „schwierig sei Lunge und Schwimmblase auf der aufgrund der Gene, die ausgeprägt werden und der Luftatmung zu definieren.“5
Kritische Würdigung der vorgestellten Studien
Zunächst einmal sei gewürdigt, dass in den hier vorgestellten Studien die Leistungsfähigkeit der modernsten Technologie zur Genomsequenzierung demonstriert und für das Studium der bislang umfangreichsten Genome angewendet wird. Es wird erwartet, dass es mit diesen Methoden (Nanopore-Sequenzierung) noch in diesem Jahr gelingen könnte, das Humangenom tatsächlich lückenlos von Anfang bis Ende zu sequenzieren (Eisenstein 2021).
Auffällig ist, dass beide Autorenteams ihre Studien zum Genom von Lungenfischen unter den Leitgedanken evolutionärer Konzepte stellen und nicht den naheliegenden ersten Schritt gehen, die neu gewonnene Einsicht in das Genom zu einem vertieften Verständnis der Lungenfische und deren Lebensweisen zu nutzen. Dabei ist es für den kritischen Leser auffällig und befremdlich, dass der Eindruck erweckt wird, als bestätigten die Resultate etablierte Evolutionsvorstellungen, was aber gar nicht der Fall ist.
Genomdaten geben zunächst einfach Auskunft über die genetische Ausstattung eines Lebewesens. Es ist noch viel Arbeit zu leisten, um zu verstehen, wie diese Ausstattung durch heutige Lebewesen genutzt wird und welches Potential darin für den Organismus liegt. Anhaltspunkte für Interpretationen genetischer Daten hinsichtlich dynamischer Entwicklungsprozesse oder gar zur Rekonstruktion vermuteter historischer Prozesse bietet das Genom nicht. Dieser Umgang mit den Daten entspringt einem „Voreingenommensein“, das nicht unüblich ist, aber in den oben vorgestellten Fällen zu einer starken Verzerrung führt, wenn der Eindruck erweckt wird, dass hier starke Bestätigungen für einen Übergang von Wasser- zum Landleben vorliegen. Die Genomdaten sagen im Blick auf ihr Gewordensein zunächst einmal gar nichts aus. Evolution, hier konkret der Übergang vom Leben im Wasser zum Festland, wird vorausgesetzt und isolierte genetische Befunde aus der Genomsequenz werden in diesen Zusammenhang eingefügt und dann die etablierte Geschichte neu erzählt. Die Genomdaten dokumentieren zunächst einmal einfach den molekulargenetischen Hintergrund der Lungenfische; inwieweit diese Daten dann in einem weiteren Schritt evolutionsbiologisch plausibel interpretiert werden können, steht auf einem anderen Blatt. Vergleichende bioinformatische Analysen basieren auf Modellen, die bereits Evolutionskonzepte beinhalten und sind daher keine unabhängigen Belege für evolutionäre Prozesse.
Die von Meyer et al. (2021) und Wang et al. (2021) dargestellten Befunde und ihre Interpretation als Beleg für einen Übergang vom Leben im Wasser ans Festland greifen einzelne Gene bzw. Genklassen heraus und stellen sie in besagten Zusammenhang. Für einen Übergang vom Wasser- zum Landleben reichen sie aber bei weitem nicht aus; dafür ist noch sehr viel mehr nötig. Vor allem wird von diesen Autoren – wie auch von vielen anderen – die grundlegende Frage nicht gesehen und bedacht, was denn Lebewesen wie die Lungenfische, die für ein Leben in ihrem Lebensraum hervorragend ausgestattet sind, veranlassen sollte, ihre Anlagen für die Eroberung eines neuen Ökosystems zu erweitern. Die Autoren gehen davon aus, dass es zunächst ausschließlich Leben im Wasser gab und Landlebewesen sich aus diesen Lebensformen entwickelt haben müssen. Sie können zeigen, dass eine kleine Auswahl der Daten sich in ein solches Szenario einfügen lässt. Sie vernachlässigen aber, dass die Daten an sich einen solchen Prozess nicht erforderlich erscheinen lassen und dass andere, alternative Szenarien denkbar sind.
Anmerkungen
1 Das griechische Wort dipnos könnte man etwa Doppelatmer wiedergeben.
2 “Genomic preadaptions in lungfish for the water-to-land transition of vertebrates”
3„cemetery of TEs“
4 So die Titelformulierung der Autoren: Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes.
5 „Therefore, it is difficult to define the lung and gas bladder based on the genes that are expressed and the air-breathing function“ (Tatsumi et al. 2016, 6).
Literatur
Bi X et al. (2021) Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes. Cell 184, 1–15; https://doi.org/10.1016/j.cell.2021.01.046
Einsenstein M (2021) Closing in on a complete human genome. Nature 590, 679–681.
Meyer A et al. (2021) Giant lungfish genome elucidates the conquest of land by vertebrates. Nature 590, 284–289.
Tatsumi N et al. (2016) Molecular developmental mechanism in polypterid fish provides insight into the origin of vertebrate lung. Sci. Rep. 6, 30680; doi:10.1038/srep30580
Wang W et al. (2021) African lungfish genome sheds light on the vertebrate water-to-land transition. Cell 184, 1–15. https://doi.org/10.1016/j.cell.2021.01.047
Autor dieser News: Harald Binder
© 2021, http://www.genesisnet.info/schoepfung_evolution/n289.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.11.19 Parmastega – neuer erster Vierbeiner?
Der neu entdeckte fossil erhaltene Vierbeiner (Tetrapode) Parmastega weist ein unerwartetes Merkmalsmosaik auf. Unter den gut erhaltenen Tetrapoden ist Parmastega zwar die geologisch älteste Gattung und steht gemäß einer cladistischen Analyse (Stammbaum-Rekonstruktion) an der Basis der Tetrapoden, weist aber mehrere markante Merkmale auf, die einer basalen Position deutlich widersprechen, allen voran ein weitgehend knorpeliges Skelett und ungewöhnlich hochstehende Augen. Das gesamte Merkmalsmosaik passt nicht in bestehende evolutionstheoretische Modellierungen, ist jedoch leichter verständlich, wenn man wie im Rahmen von Schöpfungsmodellen von einer freien Kombinierbarkeit von Merkmalen ausgeht.
Wenn fossile Arten neu entdeckt werden, wird häufig behauptet, sie würden immer wieder die Evolutionstheorie bestätigen oder füllten Lücken im evolutionären Stammbaum. Diese Aussage ist fast trivial, denn wenn man von einem gemeinsamen Stammbaum aller Arten ausgeht, wird jeder Fund automatisch eine Lücke füllen. Die interessantere Frage ist aber, ob neue Fossilien an zuvor erwarteten Stellen im bis dato geltenden Stammbaum eingeordnet werden können. Diese Frage muss bei neuen Funden sehr oft verneint werden. Vielmehr erfordern sie häufig die Hinzunahme eines neuen Astes im hypothetischen Stammbaum. Und nicht selten muss angenommen werden, dass bestimmte Merkmale konvergent entstanden sind, also zwei- oder mehrfach unabhängig, weil das gesamte Merkmalsmosaik der betreffenden Art nicht an eine Stelle des bisherigen Stammbaumes passt, oder es muss eine Rückentwicklung (Reversion) postuliert werden. Solche Fälle, in denen Ähnlichkeiten nicht auf gemeinsame Vorfahren zurückgeführt werden (können), werden unter „Homoplasien“ zusammengefasst. Sowohl das Auftreten von Homoplasien als auch die Notwendigkeit, eine zusätzliche evolutionäre Linie annehmen zu müssen, kann jedoch nicht als „Bestätigung der Evolutionstheorie“ gelten (wenn man umgekehrt auch nicht von Widerlegung sprechen kann).
Man kann immer, egal welche Merkmalsmosaike neue Funde liefern, Cladogramme erstellen; das sind Ähnlichkeitsbäume, die nach bestimmten Vorschriften bzw. Verfahren auf der Basis einer Merkmalsanalyse und der Verteilung von Merkmalen und Merkmalsausprägungen erstellt werden. Bestehende Cladogramme werden durch neue Funde oft umorganisiert. Eine Infragestellung der zugrunde liegenden Evolutionsanschauung ist auf diese Weise methodisch aber nicht möglich. Man kann nur eine Art Gütekriterium anwenden: Je mehr Homoplasien in einem Cladogramm auftreten, desto problematischer ist eine solche Situation für evolutionstheoretische Modellierungen.1
Parmastega. Die geschilderte Situation trifft beispielhaft auf einen neuen Fund eines geologisch alten Vierbeiners (Tetrapoden) zu: den auf 372 Millionen radiometrische Jahre (MrJ) datierten Parmastega aelidae aus dem Oberdevon (unteres Famennium) Russlands (Beznosov et al. 2019). (Bild einer Rekonstruktion hier: https://www.scinexx.de/news/geowissen/aeltester-gut-erhaltener-tetrapode-entdeckt/) Der Fund gilt als das älteste gut erhaltene Fossil eines frühen Tetrapoden; es wurden allerdings deutlich ältere Vierbeiner-Fußspuren (s. u.) und einige geringfügig ältere kleinere Knochenfragmente mutmaßlicher Tetrapoden gefunden, die jedoch keine Rekonstruktionen des Körperbaus ermöglichen. Zu den bisher geologisch ältesten gut erhaltenen Tetrapoden-Fossilien gehören das berühmte Ichthyostega und Acanthostega, die auf 360 MrJ datiert werden.

Abb. 1: Künstlerische Rekonstruktion von Parmastega aelidae. (Wikimedia: Entelognathus, CC BY-SA 4.0)
Von Parmastega wurden mehr als 100 gut erhaltene Knochen vor allem des Schädels und des Schultergürtels gefunden, die von vielen Individuen stammen und aus denen sich ein ca. ein Meter langes Tier rekonstruieren lässt, das wesentliche Merkmale der Tetrapoden im Bereich des Schädels aufwies. Allerdings fällt ein Merkmal des Schädels ziemlich aus dem Rahmen: Auf der Oberseite des Kopfes saßen größere Augen in einer Position, wie man sie bei Krokodilen kennt. Daher vermuten die Forscher, dass Parmastega an der Wasseroberfläche schwamm, ähnlich wie heute die Krokodile, und dort vermutlich auf Beute lauerte. Für seine räuberische Lebensweise spricht sein großes Maul mit spitzen, langen Zähnen.
Anders als bei heutigen Krokodilen war die Position der Nasenöffnung jedoch tief in der Nähe der Kiefer und somit im Wasser, woraus eine Kiemenatmung erschlossen wird. Parmastega besaß in seinem Schädel aber auch ein großes Atemloch (Spiraculum, Spritzloch), was eine Luftatmung ermöglicht haben könnte.
Beine anstelle paariger Flossen wurden allerdings nicht gefunden; ihre Existenz wurde vielmehr aus dem Bau der erhaltenen Teile des Schultergürtels erschlossen (Beznosov et al. 2019). Da die Schultern nur teilweise und Wirbel gar nicht verknöchert waren, glauben die Wissenschaftler, dass Parmastega trotz Besitz von Beinen nicht an Land gehen konnte. Auffällig sei außer der Abwesenheit von Beinknochen auch das Fehlen von Rippen, Wirbeln oder Hüftknochen trotz augenscheinlich guter Erhaltungsbedingungen. Wären diese Skeletteile verknöchert gewesen, wären also auch fossile Reste davon zu erwarten. Die fossile Abwesenheit dieser Skelettteile spreche daher dafür, dass auch diese Skelettelemente nicht verknöchert, sondern knorpelig waren. Wenn Parmastega aufgrund eines relativ weichen Skeletts nicht auf Land gehen konnte, stellt sich allerdings die Frage, wozu unter diesen Umständen Tetrapodenbeine ausgebildet waren. Für ein Leben im Wasser spricht, dass im Bereich des Schädels das Seitenlinienorgan gut ausgebildet war; dieses Organ enthält Sensoren, mit denen Wasserströmungen wahrgenommen werden können.
Insgesamt ist die Lebensweise von Parmastega etwas rätselhaft: Die hochstehenden Augen machen nur Sinn, wenn der Blick über die Wasseroberfläche ging und es eine Interaktion mit der Umgebung auf dem Land gab (Beznosov et al. 2019, 530). Aber was soll Parmastega dort gesucht haben? Da das Tier vermutlich nicht landgangtauglich war, mutmaßen die Forscher, dass es im Wasser gelauert und am Ufer trinkende oder ruhende Beute ins Wasser gezogen haben könnte – ähnlich wie Krokodile das heute tun. Nach gängigen evolutionstheoretischen Vorstellungen gab es aber noch keine größeren Landlebewesen als geeignete Beute (Fröbisch & Witzmann 2019, 494). Die Ausstattung mit großen Reißzähnen spricht jedenfalls klar dafür, dass größere Tiere erbeutet wurden; vielleicht auch am Ufer liegende Kadaver. Hier gibt es offensichtlich noch ein Rätsel zu lösen: Wie war die Fortbewegungsweise von Parmastega wirklich, und gab es wirklich noch keine größeren landlebenden Tiere? Immerhin sind eindeutige Vierbeinerspuren bekannt, die auf bis zu 390 Millionen radiometrische Jahre und somit 18 Millionen Jahre älter als Parmastega datiert werden (Niedzwiedzki et al. 2010; Ahlberg 2018; vgl. Junker 2019).
Evolutionstheoretische Betrachtungen. In der von Beznosov et al. (2019) durchgeführten cladistischen Analyse wird Parmastega an der Basis aller anderen Tetrapoden platziert, allerdings ist diese Position nicht sonderlich stabil und die Auflösung des Cladogramms schwach. Die Position von Parmastega im Cladogrammen wird auf der Basis des gesamten vorliegenden Merkmalsspektrums ermittelt und auf dieser Basis passt sie zu den gegenwärtigen evolutionstheoretischen Vorstellungen. Nimmt man jedoch einzelne Merkmale in Augenschein, ergibt sich ein deutlich anderes Bild: Einen teilweise krokodilartigen Kopf hätte man bei einem frühen Tetrapoden evolutionstheoretisch sicher nicht erwartet. Fröbisch & Witzmann (2019, 494) kommentieren überrascht: „This eye shape and position is surprising because it indicates that this water dweller was looking above the surface of the water.“ Noch rätselhafter finden Beznosov et al. (2019, 530), dass ein größerer Teil des Skeletts knorpelig ausgebildet ist, denn alle näher verwandten Formen – mutmaßliche Vor- und Nachfahren – haben ein gut verknöchertes Skelett2, so dass Parmastega in dieser Hinsicht völlig aus dem phylogenetischen Rahmen fällt.
Der Hirnschädel von Parmastega ist morphologisch zwischen dem von Ichthyostega, von Acanthostega und Ventastega angesiedelt und könnte als hypothetische Ausprägung eines Vorfahren dieser recht verschiedenen Gattungen interpretiert werden, doch eine darauf aufgebaute Phylogenie führt zu einer „nicht-trivialen Homoplasie“ entweder beim Hirnschädel oder bei anderen Teilen des Skeletts (Beznosov et al. 2019, 530), d. h. zu Merkmalswidersprüchen. Beznosov et al. (2019, 530) kommen daher zum Schluss, dass Parmastega phylogenetisch zwar am ehesten an der Basis der gut erhaltenen Tetrapoden steht, jedoch nicht unbedingt charakteristisch für die primitiven Bedingungen für die Gruppe sei.3 Und sie sprechen von „erheblichen morphologischen Homoplasien unter devonischen Tetrapoden“ (S. 530f.).
Damit erweist sich Parmastega als ausgeprägte Mosaikform, die keineswegs eine bisher bestehende Lücke im Stammbaum füllt, sondern eher neue Lücken aufreißt und evolutionstheoretisch als weitere Seitenlinie angesehen werden muss. Das gilt auch, weil wie oben erwähnt viel älter datiere eindeutige Tetrapoden-Fußspuren bekannt sind. Auch daher scheidet Parmastega genauso wie andere frühe Tetrapoden als evolutionäre Übergangsform aus. Allem Anschein nach war Parmastega adaptiv für eine bestimmte Lebensweise optimiert und nicht „auf dem Weg zu irgendetwas“ (eine Formulierung von Ahlberg 2018, der dies über die den Tetrapoden nahe stehenden Elpistostegiden sagt). Fröbisch & Witzmann (2019, 495) kommentieren: „Diese Entdeckung erinnert uns auch daran, dass im nächsten spannenden Kapitel dieser Detektivgeschichte noch viel zu lernen bleibt.“4
Das Merkmalsmosaik von Parmastega passt besser zur Vorstellung einer freien Kombinierbarkeit von Merkmalen bzw. Merkmalskomplexen und zu netzförmigen Ähnlichkeitsbeziehungen, die aus einer Schöpfungsperspektive leichter verständlich sind. Dagegen haben stammesgeschichtliche Rekonstruktionen mit Parmastega mit weiteren teils schwerwiegenden Homoplasien zu kämpfen, was nicht den evolutionstheoretischen Erwartungen entspricht (vgl. Anmerkung 1).
Literatur
Ahlberg PE (2018) Early Vertebrate Evolution. Follow the footprints and mind the gaps: a new look at the origin of tetrapods. Earth Environ. Sci. Trans. R. Soc. Edinb., 1–23. doi:10.1017/S1755691018000695
Beznosov PA, Clack JA, Lukševičs E, Ruta M & Ahlberg PD (2019) Morphology of the earliest reconstructable tetrapod Parmastega aelidae. Nature 574, 527-531.
Fröbisch NB & Witzmann F (2019) Early tetrapods had an eye on the land. Nature 574, 494-495.
Junker R (2019) Entstehung der Vierbeiner – doch kein glatter Übergang. Stud. Integr. J. 26, 106-108.
Niedzwiedzki G, Szrek P, Narkiewicz K, Narkiewicz M & Ahlberg PE (2010) Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43-48.
Anmerkungen
1 Der Grund dafür ist: Bei der Erstellung von Cladogrammen soll die Anzahl der anzunehmenden Konvergenzen bzw. Homoplasien minimiert werden, weil Homoplasien als evolutionstheoretisch unwahrscheinlich gelten. Cladogramme werden auf der Basis gemeinsamer abgeleiteter („höherentwickelter“) Merkmale erstellt, die als Marker für gemeinsame Vorfahren verwendet werden. Dese werden als Synapomorphien bezeichnet. Wenn Synapomorphien genauso wahrscheinlich wären wie Homoplasien, könnten cladistische Analysen gar nicht stammesgeschichtlich interpretiert werden.
2 „Even more puzzling is the fact that this poorly ossified postcranial skeleton is apomorphic: elpistostegids are well-ossified, as are the majority of tetrapodomorph fishes.“
3 „Parmastega is phylogenetically the least-crownward of all of the non-fragmentary tetrapods, but it is not necessarily representative of the primitive conditions for the group.“
4 „This discovery also reminds us that much still remains to be learnt in the next gripping chapter of this detective story.“
Autor dieser News: Reinhard Junker
© 2019, http://www.genesisnet.info/schoepfung_evolution/n273.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.12.17 Komplexaugen: Komplex von Anfang an
In den ältesten Schichten des Unterkambriums und – evolutionstheoretisch – somit an der Basis der „kambrischen Explosion“ wurden bei einem Trilobiten erstmals zellulär erhaltene Komplexaugen entdeckt. Sie zeigen im Wesentlichen den gleichen grundsätzlichen Aufbau eines Komplexauges wie bei Augen heutiger Bienen oder Libellen.
Die Entstehung von Augen ist eines der Lieblingsbeispiele von Evolutionstheoretikern, mit denen sie zeigen möchten, dass mittlerweile im Wesentlichen verstanden sei, wie ein komplexes Organ schrittweise evolutiv entstehen konnte. Auch in der Wikipedia wird dieser Eindruck vermittelt. Er entspricht jedoch in keiner Weise den Tatsachen. Die Aneinanderreihung von sechs unterschiedlich komplexen Augentypen werden der Komplexität dessen, was evolutionstheoretisch durch blinde Prozesse erklärt werden müsste, in keiner Weise gerecht (Ullrich et al. 2006). Augenserien, mit denen eine evolutive Entstehung plausibel gemacht werden soll, entsprechen zudem nicht einmal hypothetischen stammesgeschichtlichen Abfolgen konkreter Organismen, sondern sind theoretische Modellreihen. Zudem müsste aufgrund der sehr unsystematischen Verteilung von Augentypen im Tierreich im Rahmen evolutionstheoretischer Modellbildungen angenommen werden, dass etwa der Linsenaugen-Typ, wie ihn z. B. Wirbeltiere besitzen, vielfach unabhängig aus einfacheren Augentypen entstanden sein müsste (Jonasova & Kozmik 2008, fig. 1). Ähnliches gilt auch für andere Augentypen.
Weniger bekannt scheint zu sein, dass der Fossilbericht eine Evolution komplexer Augen in keiner Weise unterstützt – im Gegenteil: Sowohl Linsenaugen des Kameratyps als auch Komplexaugen (Facettenaugen) sind unter den ältesten vielzelligen Tieren der „kambrischen Explosion“ anzutreffen. Der räuberisch lebende Anomalocaris aus dem Unterkambrium besaß sogar Facettenaugen, die zu den größten und bestauflösenden bekannten Facettenaugen gehören und diesbezüglich nur von den Komplexaugen einiger Libellen übertrumpft werden (Paterson et al. 2011).

Abb. 1: Rekonstruktion von Anomalocaris. (Wikimedia: Junnn11, CC BY-SA 4.0)
Bislang konnte man anhand der fossilen Erhaltung der kambrischen Fossilien allerdings nur die äußere Struktur von Komplexaugen untersuchen. Ein neuer hervorragend erhaltener Fund mit zellulär erhaltenen Augenstrukturen erlaubt nun aber auch Einblicke in den inneren Feinbau eines Trilobiten-Auges. Die Zoologin Brigitte Schoenemann von der Universität Köln und ihre Kollegen beschreiben die Details eines Komplexauges des Trilobiten Schmidtiellus reetae, der in den untersten Schichten des unteren Kambriums in Estland gefunden wurde. Trilobiten sind eine im Kambrium weit verbreitete Gruppe der Gliederfüßer. Die fossilen Augen zeigen, dass ihr Aufbau im Wesentlichen dem Bau heutiger sogenannter Appositionsaugen wie z. B. von Bienen oder Libellen entspricht. Bei Appositionsaugen ist jedes Einzelauge (Ommatidium) durch Pigmente von seinen Nachbaraugen abgeschirmt. Es fehlt bei Schmidtiellus allerdings die Linse, was jedoch durch einen besonderen Bau der Einzelaugen ausgeglichen wird, so dass nach Abschätzung der Wissenschaftler die Lichtstärke des Auges nicht geringer war als bei vergleichbaren heutigen Einzelaugen. „There is an elegant physical solution, however, of how to develop a quality image of modern type“ (Schoenemann et al. 2017, 1); „So, in total, the system of the trilobite S. reetae is in a state similar to modern aquatic arthropods, even without a lens“ (Schoenemann et al. 2017, 5).
Das Auge besteht aus etwa 100 Einzelaugen, die im Vergleich zu heutigen Formen relativ weit auseinander stehen. Jedes Ommatidium bestand wie in modernen Facettenaugen aus acht Sinneszellen: ein zentrales Lichtleiterstäbchen (Rhabdom) und sieben sich darum gruppierende Elemente. Der Fund zeige, dass „hochentwickelte (‚sophisticated’) Augen schon zu Beginn des Fossilberichts höherer Organismen existierten“, stellen die Wissenschaftler fest (Schoenemann et al. 2017, 1).
Mit nur etwa 100 „Pixeln“ war die Sehleistung freilich gering, doch ermöglichte die Anordnung der ca. 1 x 0,5 cm großen Augen ein breites Sichtfeld nach vorne und zur Seite, innerhalb dessen Bewegungen und Helligkeitsverteilungen wahrgenommen werden konnten. Doch wie erwähnt sind aus dem Unterkambrium auch ausgesprochen hochauflösende Komplexaugen bekannt; Schoenemann et al. (2017, 5) weisen selbst auf einen weiteren baltischen Trilobiten, Holmia kjerulfi, hin, der in nur wenig jünger datierten Schichten gefunden wurde und hochauflösende Augen mit dicht gepackten Einzelaugen besaß, eine Konstellation, die den Augen heutiger Libellen vergleichbar war.
Schoenemann et al. (2017, 1) merken an, dass der Ursprung der Trilobiten insgesamt unklar sei. Der fossile Nachweis von Facettenaugen, deren Grundaufbau im Wesentlichen „modern“ ist, in den ältesten kambrischen Schichten entspricht ziemlich dem Gegenteil dessen, was evolutionstheoretisch zu erwarten wäre.
Literatur
Jonasova K & Kozmik Z (2008) Eye evolution: Lens and cornea as an upgrade of animal visual system. Sem. Cell Dev. Biol. 19, 71-81.
Paterson RJ, García-Bellido DC, Lee MSY, Brock GA, Jago JB & Edgecombe GD (2011) Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes. Nature 480, 237-240.
Schoenemann B, Pärnaste H & Clarkson ENK (2017) Structure and function of a compound eye, more than half a billion years old. Proc. Natl. Acad. Sci., doi:10.1073/pnas.1716824114.
Ullrich H, Winkler N & Junker R (2006) Zankapfel Auge. Ein Paradebeispiel für „Intelligent Design“ in der Kritik. Stud. Integr. J. 13, 3-14. http://www.wort-und-wissen.de/sij/sij131/sij131-1.html
Genesisnet-Artikel zur „Kambrischen Explosion
- Kambrische Explosion, https://www.genesisnet.info/index.php?Artikel=42842&Sprache=de&l=1
- Kambrische Explosion: Darwins Dilemma gelöst?, https://www.genesisnet.info/index.php?News=202
Autor dieser News: Reinhard Junker
© 2017, http://www.genesisnet.info/schoepfung_evolution/n252.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
10.10.16 Von Fischen zu Vierbeinern? Neues von Acanthostega
Ein Fisch mit Fingern – Acanthostega, mutmaßliches Bindeglied zwischen Fischen und Vierbeinern – wurde neu untersucht. Dabei stellte sich heraus, dass fast alle Funde von noch nicht ausgewachsenen Tieren stammen. Es bliebt dabei: Acanthostega war allem Anschein nach ausschließlich wasserlebend und passt nicht gut als Übergangsform.
Der sogenannte „Spitzpanzer“ – Acanthostega – aus dem Oberdevon spielt eine prominente Rolle in evolutionären Hypothesen zur Entstehung der Tetrapoden (Vierbeiner) ausgehend von Fischen. Denn er lebte zwar wie Fische ausschließlich im Wasser, besaß aber Finger und damit ein typisches Vierbeiner-Merkmal.

Abb. 1: Rekonstruktion des Skeletts von Acanthostega gunnari. (Wikimedia: Ryan Somma, CC BY-SA 2.0)
Eine detaillierte Neu-Untersuchung von 200 fossilen Acanthostega-Knochen und 14 Schädel-Fossilien dieser Gattung zeigte nun, dass es sich dabei fast ausnahmslos um Jungtiere handelt (Sanchez et al. 2016; vgl. Fröbisch 2016). Das macht die Beurteilung der Lebensweise der ausgewachsenen Formen schwieriger als sie ohnehin schon war. Die neuen Befunde sprechen dafür, dass die Tiere im Jugendstadium nicht in der Lage waren, an Land zu kriechen. Die Funde stammen von mindestens 20 Individuen und befinden sich alle von einem kleinen Areal der Britta-Dal-Formation des Oberdevons Ostgrönlands; Sanchez et al. (2916) vermuten, dass sie gemeinsam bei einer Dürre nach einer Schichtflut verendet sind.
Anders als in populärwissenschaftlichen Darstellungen behauptet, ist das aber keine neue Erkenntnis. Denn schon bisher deuteten viele anatomische Merkmale darauf hin, dass Acanthostega ausschließlich wasserlebend war – trotz des Besitzes von acht Fingern, eine ungewöhnliche Merkmalskombination (vgl. zusammenfassende Darstellung bei Junker 2004). Gründe dafür waren u. a.: Die Bezahnung gleicht insgesamt zeitgenössischen Fleischflosser-Fischen und keinem Tetrapoden, der Schädel ist mit dem Schultergürtel relativ fest verbunden, Acanthostega ähnelt im Kiemenskelett kiemenatmenden Lungenfischen, die Wirbelsäule ist von vorne bis hinten auffallend gleichförmig, ähnlich wie bei Fischen wie dem Quastenflosser Eusthenopteron, eine Verbindung Becken-Wirbelsäule bestand vermutlich nur durch Bänder; die Hüfte konnte dadurch kaum das Körpergewicht tragen; insgesamt wirken die Extremitäten eher als Paddel denn als Füße (Clack 2002, 122-127; vgl. Zusammenfassung bei Junker 2004). Clack (2002, 124) stellt als Gesamteindruck fest: Fast alle Merkmale von Acanthostega legen ein ausschließliches Wasserleben nahe. Die relativ starren Extremitäten waren vermutlich gut geeignet, um mit kräftigen Paddelstößen plötzlich aus der Ufervegetation Beute zu erhaschen. Vermutlich lebte Acanthostega in vegetationsreichen Uferzonen, in denen eine tetrapodenartige Extremität passend war. Wie alle anderen oberdevonischen Tetrapoden ist auch Acanthostega ausschließlich zusammen mit Fischen fossil überliefert (Clack 2002, 110).
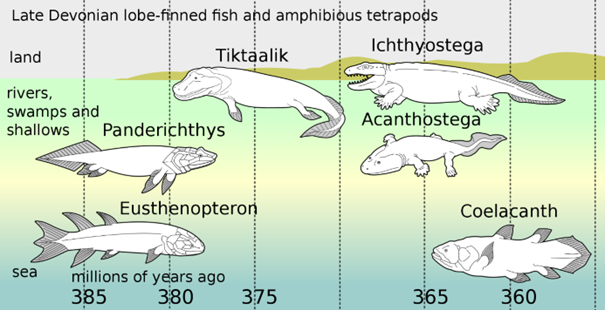
Abb. 2: Rekonstruktionen von verschiedenen Tetrapoden sowie von deren vermeintlichen Vorfahren samt (radiometrisch datieren) Fundalter. (Wikimedia: Original: Dave Souza Vector: Pixelsquid – derived from: Fishapods.jpg: Dave Souza, CC BY-SA 3.0)
Dass es sich um Jungtiere handelt, schließen die Forscher daraus, dass das Oberarmskelett der Tiere noch nicht verknöchert war. Außerdem zeigte eine Untersuchung der inneren Feinstruktur der Knochen mit einem Röntgensynchrotron, dass die Tiere zwar schon sechs Jahre und älter waren, ihre Beinknochen aber noch keine Anzeichen für eine Verlangsamung des Wachstums zeigten – ebenfalls ein Hinweis darauf, dass die Tiere noch nicht ausgewachsen waren. Ein knorpeliger Oberarmknochen wäre für die Bewegung an Land ungeeignet gewesen. Sanchez et al. (2016) stellten weiter fest, dass es zwei Größenklassen gab, möglicherweise bedingt durch Entwicklungs-Plastizität (d. h. unterschiedliche Ausprägungen werden durch verschiedene Umweltreize ausgelöst).
Dass Acanthostega wasserlebend war, ist also nicht das Überraschende, wohl aber, dass es sich bei den (nur) in Grönland gefundenen Fossilien um Jungtiere handelt. Damit ist unklar, wie die erwachsenen Tiere ausgesehen haben, welche Rolle sie bei der Entstehung der Vierbeiner gespielt haben könnten und ob sie vielleicht doch auch an Land leben konnten. Die neuen Befunde stellen bisherige Hypothesen in Frage, wonach vermutet worden war, dass die Larven der ersten Tetrapoden die ersten Landgänger waren. Dazu waren die Acanthostega-Jungtiere jedenfalls kaum in der Lage.
Zur Frage nach der Evolution der Vierbeiner steuern die neuen Befunde nichts Wesentliches bei, das nicht schon bekannt war. Es bleibt dabei, dass einzelne Merkmale markant verändert im Vergleich zu potentiellen Vorläufern auftauchen (z. B. im Bereich des Hirnschädels oder der Besitz von Fingern). Das Handgelenk von Acanthostega war späteren Tetrapoden sehr unähnlich (Clack 2002, 137). Neben Merkmalen, in denen Acanthostega abgeleiteter ist als das berühmte Ichthyostega (Carroll 1992, 60), gibt es auch Eigenschaften, bei denen das Umgekehrte zutrifft. Die Wangen und das Schädeldach sind bei Acanthostega und Ichthyostega fest verbunden, im Gegensatz sowohl zu den möglichen Fisch-Vorfahren als auch zu anderen frühen Tetrapoden (Carroll 1992, 60). Das heißt, evolutionär gesehen wäre hier ein Zickzackkurs gefahren worden. Carroll (1992, 60) bemerkt: Verschiedene Spezialisierungen schließen die Möglichkeit einer direkten Vorfahrenschaft für spätere Tetrapoden aus.
Interessant ist auch die Einschätzung von Sanchez zur Bedeutung der neuen Erkenntnisse: „Unsere Studie gibt uns nur einen allerersten Einblick in die Lebensweise eines frühen Tetrapoden. … Wir wollen nun auch die Lebensgeschichte anderer früher Vierbeiner untersuchen. Möglicherweise stoßen wir dabei auf Erkenntnisse, die unser Lehrbuchwissen völlig verändern“ (http://www.scinexx.de/wissen-aktuell-20592-2016-09-08.html).
Literatur
Carroll RL (1992) The primary radiation of terrestrial vertebrates. Annu. Rev. Earth Planet. Sci. 20, 45-84.
Clack JA (2002) Gaining ground. The origin and evolution of Tetrapods. Bloomington and Indianapolis: Indiana University Press.
Fröbisch NB (2016) Evolution: Teenage tetrapods. Nature 537, 311-312.
Junker R (2004) Vom Fisch zum Vierbeiner – eine neue Sicht zu einem berühmten Übergang. Teil 2: Ichthyostega, Acanthostega und andere Tetrapoden des höheren Oberdevons. Stud. Integr J. 11, 59-66.
Sanchez S, Tafforeau P, Clack JA & Ahlberg PE (2016) Life history of the stem tetrapod Acanthostega revealed by synchrotron microtomography. Nature 537, 408-411.
Autor dieser News: Reinhard Junker
© 2016, http://www.genesisnet.info/schoepfung_evolution/n240.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
04.07.16 Amphibische Fische: Mindestens 33-mal unabhängig entstanden
Eine detaillierte Untersuchung über das Vorkommen von Fischen, die zeitweise auch an Land gehen, zeigt, dass diese Fähigkeit mindestens 33-mal unabhängig entstanden sein muss. Aus evolutionstheoretischer Sicht war ein solcher Befund nicht erwartet worden. Die Erklärung, dass der Erwerb dieser Fähigkeit gar nicht so schwierig sei, beruht auf einem Zirkelschluss.
Bei Amphibien, also Tieren, die teils an Land und teils im Wasser leben, denkt man an Frösche, Kröten, Unken oder Salamander. Dass auch unter Fischen amphibisch lebende Arten vertreten sind, ist weniger bekannt; doch es gibt eine ganze Menge davon. Manche von ihnen verbringen nur einige Sekunden oder Minuten an Land, andere dagegen Stunden oder sogar Tage. Besonders bekannt unter Letzteren sind die Schlammspringer (Gattung Periophthalmus), die mehr Zeit an Land als im Wasser verbringen und dafür eine Reihe spezieller Bauplanbesonderheiten besitzen, oder die Aale, die längere Wanderungen auf Land durchführen können und dabei über die Haut atmen.

Abb. 1: Der Schlammspringer Periophthalmus modestus. (Wikimedia: Alpsdake, CC BY-SA 4.0)
Wegen der besonderen Anforderungen für ein zeitweiliges Leben an Land könnte man denken, dass amphibische Fische auf evolutivem Wege eher selten entstanden sind; das bringt auch Terry Ord, der mit Georgina Cooke jüngst eine Studie über deren Vorkommen durchgeführt hat, explizit zum Ausdruck („Because of the challenges fish face in being able to breathe and move and reproduce on land, it had been thought this was a rare occurrence“, http://phys.org/news/2016-06-fish-common-thought.html). Doch schon Coates & Clack (1995, 301) erwähnen, dass unter den Strahlflosser-Fischen, die nicht eine Verbindung zum Landgang der Vierbeiner gebracht werden, 59 amphibisch lebende Arten aus 16 marinen* und limnischen* Familien bekannt seien, von denen keine tetrapodenartige* Beine entwickelt hat.
*Begriffe:
- marine (= im Meer lebend)
- limnisch (= im Süßwasserbereich vorkommend)
- tetrapodenartig (= vierbeinerartig)
In einer jüngst veröffentlichten detaillierten phylogenetischen Studie wiesen Ord & Cooke (2016) insgesamt sogar 130 amphibisch lebende Fischarten nach. Diese sind auf 33 Familien verteilt, und die Autoren interpretieren diesen Befund so, dass damit auch die Mindestanzahl einer unabhängigen (konvergenten) Entstehung amphibischer Fische bei 33 liegt. Vermutlich ist diese Zahl noch deutlich höher, denn alleine bei der genauer untersuchten Familie der Blenniidae schließen die Autoren auf eine siebenmalige konvergente Entstehung eines „in hohem Maße amphibischen Lebensstils“. Die Fisch-Familien, bei denen es amphibische Arten gibt, sind ökologisch z. T. sehr unterschiedlich und kommen sowohl in Süß- als auch Salzwasser vor.
Aus diesem unerwarteten Befund schließen die Autoren, dass die evolutive Entstehung der amphibischen Lebensweise nicht so schwer sein könne wie bisher angenommen, da sie sich so häufig ereignet habe. Diese Schlussfolgerung ist unlogisch. Denn die Häufigkeit des Vorkommens dieser Lebensweise hat mit der Frage, wie schwierig die dafür nötigen Voraussetzungen zu erfüllen sind, nichts zu tun. Schließlich war es beispielsweise auch nicht deshalb leichter als gedacht, eine Mondrakete zu bauen, weil das sowohl Russen als auch Amerikaner unabhängig geschafft haben. Vielmehr handelt es sich hier um einen evolutionären Zirkelschluss; er funktioniert wie folgt: 1. Eigentlich ist es für einen Fisch schwierig, evolutiv eine amphibische Lebensweise zu entwickeln. 2. Daher ist zu erwarten, dass das nicht oft vorkommt. 3. Es kommt aber oft vor, daher kann es doch nicht so schwierig sein. Diese Logik funktioniert natürlich nur, wenn man Evolution schon voraussetzt. In Wirklichkeit ist der Befund des häufigen Vorkommens der amphibischen Lebensweise jedoch eine schwerwiegende Herausforderung für evolutionsbiologische Hypothesen, da auf der Basis zukunftsblinder Mechanismen ein vielfacher Übergang zu anspruchsvollen ähnlichen Fähigkeiten nicht zu erwarten ist.
Interessant sind die Ergebnisse von Ord & Cooke auch in einer weiteren Hinsicht. Die zahlreichen amphibischen Fischarten sind keine Vorstufen zu landlebenden Formen. Die amphibische Lebensweise von Fischen scheint also keinen Startvorteil für einen Übergang an ein dauerhaftes Landleben darzustellen, wie es bei den Landwirbeltieren verwirklich ist. Auf dieses Paradox weist auch Clack (2002, 103) in ihrer Monographie zum Ursprung der Landwirbeltiere hin: Es gibt Strahlflosser mit Anpassungen ans Landleben und solche, die fingerartige Strukturen von Flossenstrahlen ausgebildet haben. Diese zwei Gruppen überlappen sich nahezu nicht. Fische mit Fingern sollen – auch aus evolutionärer Sicht – nicht dauerhaft an Land gegangen sein, und solche, die es im evolutionären Verlauf geschafft haben sollen, zweitweise auf Land zu überleben, haben keine fingerartigen Strukturen an ihren Extremitäten.
Literatur
Clack JA (2002) Gaining Ground. The origin and evolution of Tetrapods. Bloomington and Indianapolis.
Coates MI & Clack JA (1995) Romer’s gap: tetrapod origins and terrestriality. In: Arsenault M, Lelièvre H & Janvier P (eds) Studies on early vertebrates (VIIth International Symposium, Miguasha Parc, Quebec). Paris: Bull. Mus. Natl. Hist. Nat. 17, 373-388.
Ord JJ & Cooke GM (2016) Repeated evolution of amphibious behavior in fish and its implications for the colonization of novel environments. Evolution, early view, DOI: 10.1111/evo.12971.
Autor dieser News: Reinhard Junker
© 2016, http://www.genesisnet.info/schoepfung_evolution/n239.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
16.07.15 Collinsium – am Anfang war die Komplexität
Einer der ältesten fossil erhaltenen Lobopoden, Collinsium ciliosum, ist ausgesprochen komplex gebaut. Die Fossilüberlieferung zeigt bei dieser Gruppe keine Abfolge von „einfacher“ zu „komplexer“. Das stellt die evolutionstheoretisch zu erwartende Abfolge auf den Kopf.
Evolution verläuft vom Einfacheren zum Komplexeren; mindestens müssen alle Baupläne des Lebens einmal einfach begonnen haben, auch wenn im weiteren Verlauf wieder Rückbildungen erfolgen können. Es zeigt sich aber immer wieder, dass bereits sehr früh in der Fossilüberlieferung Tiergruppen mit komplexen Formen überliefert sind. Die ältesten Formen einer Gruppe sind oft komplexer als jüngere. Über ein eindrucksvolles Beispiel dieser Art berichten Yang et al. (2015). Die Forscher beschreiben Collinsium ciliosum, einen ungewöhnlich gepanzerten, ca. 8,5 cm langen Lobopoden aus der unterkambrischen Xiaoshiba-Lagerstätte in Südchina (Rekonstruktion z. B. unter http://cdn4.sci-news.com/images/enlarge/image_2962_1e-Collinsium-ciliosum.jpg). Lobopoden sind ausgestorbene wurmartige Organismen mit füßchenartigen Anhängen; sie haben im Meer (marin) gelebt und werden gewöhnlich zu den auch heute lebenden Stummelfüßern (Onychophora, „Würmer mit Füßchen“) gerechnet und als deren mögliche Vorfahren diskutiert. Die etwa 180 heute lebenden Stummelfüßer sind recht einheitlich gebaut und bilden einen eher wenig bedeutenden Anteil am Ökosystem von Regenwäldern (Yang et al. 2015, 1). Eine stammesgeschichtliche Verbindung zu den marinen kambrischen Formen erscheint aufgrund der Ökologie der heute bekannten Formen nicht naheliegend.
Nicht nur in Bezug auf den Lebensraum fallen die Unterschiede auf, auch der Körperbau des kambrischen Collinsium ist für einen Lobopoden ungewöhnlich komplex. Am vorderen Teil des wurmartigen Körpers befindet sich ein Paar antennenartiger Anhänge, es folgen sechs Paare verlängerter Anhänge mit feiner Fiederung, die als Filtriereinrichtung gedeutet werden, mit der Kleinlebewesen erbeutet werden. Dahinter befinden sich bis zum Körperende neun Beinchen mit Klauen, mit denen sich Collinsium vermutlich auf fester Unterlage (Felsen oder auch auf Schwämmen) festhalten konnte. Doch damit nicht genug. Auf dem Rücken und an der Seite war das Tier mit insgesamt 72 harten, spitzen Stacheln unterschiedlicher Größe ausgestattet, die dem Tier Schutz gegen Räuber boten. Im Vergleich zu anderen Gattungen der Lobopoden ist Collinsium deutlich die komplexeste und gehört zugleich zu den ältesten und wird in die Familie der Luolishaniidae gestellt. Diese Familie steht im Dendrogramm (Ähnlichkeitsbaum) an abgeleiteter („hochentwickelter“) Position, obwohl sie in der geologischen Schichtenfolge an der Basis der Überlieferung der Lobopoden auftritt. Die Luolishaniidae weisen das größte Ausmaß an Spezialisierung der Körperanhänge von Lobopoden des Paläozoikums („Erdaltertum“ von Kambrium bis Perm) auf und sind sehr viel verschiedenartiger als Vertreter der (später fossil überlieferten) Kronengruppen der Stummelfüßer (Yang et al. 2015). Eine Abfolge von „einfach“ Richtung „komplex“, wie evolutionär ursprünglich verlaufen sein müsste, ist damit auf den Kopf gestellt. Warum diese Gruppe trotz üppiger Ausstattung ausgestorben ist, darüber kann nur spekuliert werden. Man spricht in solchen Fällen häufig (und so auch hier) von gescheiterten „evolutionären Experimenten“, womit aber nichts erklärt wird. Eine denkbare – aber spekulative – Erklärung könnte die Zerstörung des Lebensraumes der kambrischen Formen sein.
„Alle modernen Stummelfüßer sind ziemlich ähnlich in ihrem allgemeinen Körperbau und nicht so aufregend in ihrer Lebensweise. Aber im Kambrium waren ihre entfernten Verwandten erstaunlich vielfältig und traten in einer überraschenden Vielfalt bizarrer Formen und Größen auf“, wird einer der Mitautoren, Javier Ortega-Hernández von der Universität von Cambridge, UK, zitiert (http://www.sci-news.com/paleontology/science-collinsium-ciliosum-cambrian-spiky-worm-02962.html). Die Lobopoden sind nicht die einzige Gruppe, deren Fossilüberlieferung in maximaler Verschiedenartigkeit startet; Yang et al. (2015, 5) nennen als weitere Beispiele die Crinoiden (Seelilien und Haarsterne) und Brachiopoden (Armfüßer), bei denen dies auch der Fall ist. Es könnten noch mehr Gruppen genannt werden.
Literatur
Yang J, Ortega-Hernández J, Gerber S, Butterfield NJ, Hou JB, Lan T & Zhang XG (2015) A superarmored lobopodian from the Cambrian of China and early disparity in the evolution of Onychophora. Proc. Natl. Acad. Sci. 112, 8678-8683.
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n229.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
15.09.14 Evolution vom Wasser ans Land: Gar nicht so schwer?
Lebewesen sind in der Lage, auf Umweltänderungen durch Veränderungen im Bau, in der Physiologie oder im Verhalten zu reagieren (Plastizität). Das Erbgut enthält also Variationsprogramme, die bei Bedarf abgerufen werden. Diese Fähigkeit machten sich Forscher zunutze und ließen lungenatmende Flösselhechte (Polypterus) auf Land aufwachsen. In der ungewohnten Umgebung zeigten die Fische als plastische Reaktion veränderte Verhaltensweisen und Änderungen im Bau einiger Knochen, die auf dem Land eine Verbesserung der Fortbewegung ermöglichen. Könnte diese plastische Fähigkeit in der angenommenen Evolution von Fischen zu Vierbeinern eine wichtige Rolle gespielt haben?
Lebewesen haben die faszinierende und anspruchsvolle Fähigkeit, sich unterschiedlichen Umweltbedingungen individuell anpassen zu können. Bestimmte Umweltreize können Veränderungen im Körperbau, in der Physiologie oder im Verhalten auslösen, die das Überleben ermöglichen oder erleichtern. Das Erbgut wird dabei nicht geändert, vielmehr sind im Erbgut Variationsprogramme verankert, die bei Bedarf abgerufen werden können. Diese Fähigkeit nennt man Plastizität („Formbarkeit“), im deutschen Sprachraum wird auch der Begriff Modifikation verwendet. Plastizität gehört sozusagen zur Standard-Ausstattung von Lebewesen. Ein typisches Beispiel ist die Bildung von Hornschwielen an Händen oder Füßen aufgrund verstärkter mechanischer Beanspruchung der Haut.
Plastizität und Evolution. In der Evolutionsforschung hat das Phänomen Plastizität in den letzten Jahren vermehrt Aufmerksamkeit gefunden. Dabei wird der Frage nachgegangen, ob plastische Reaktionen das Potential beinhalten könnten, zu evolutionären Neuheiten zu führen. Auf den ersten Blick erscheint das wenig plausibel, denn plastische Änderungen sind keine Merkmale, die erst neu erworben wurden, sondern gehören bereits zum Repertoire der Lebewesen. Dennoch lautet eine Idee in Bezug auf Evolution wie folgt: Unter extremen Umweltbedingungen könnten aufgrund einer plastischen Reaktion Merkmale ausgeprägt werden, die sonst nicht zum Vorschein kommen und Ansätze zu einer weitergehenden Evolution darstellen könnten. Doch mehr als eine Idee ist dieser Ansatz bisher nicht.
Flösselhechte als Modell. Eine Forschergruppe hat diese Idee kürzlich angewendet auf die Frage, wie aus Fischen Vierbeiner geworden sein könnten (Standen et al. 2014). Es sind hierzu zwar einige Fossilien bekannt, doch liefern diese kaum Information zur Beantwortung der Frage, wie die zahlreichen für diesen Übergang notwendigen Änderungen abgelaufen sein könnten. Dazu machten die Forscher nun Versuche mit dem Senegal-Flösselhecht (Polypterus senegalus). Flösselhechte gelten als urtümliche Strahlflosser-Fische, die an der Basis des Strahlflosser-Fisch-Stammbaums stehen, also unweit der Abzweigung der Fleischflosser, zu denen die mutmaßlichen Landwirbeltiervorläufer gehören. In gewissem Sinne stehen sie also den vermuteten ersten Landgängern nahe.

Abb. 1: Senegal-Flösselhecht (Polypterus senegalus) in Albino-Färbung. (Wikimedia: 5snake5, CC0)
Flösselhechte besitzen eine paarige Lunge, die zugleich als Schwimmblase fungiert, und sind zur Luftatmung befähigt. Sie leben zwar fast ausschließlich im Wasser in krautigen Uferbereichen, können aber an Land überleben und von einem Wasserloch zum nächsten gelangen. Das schaffen sie, weil sie sich mit Hilfe ihrer kräftigen Brustflossen auf Land fortbewegen können und in der Lage sind, ihren Kopf etwas anzuheben. An dieser Fähigkeit knüpften die Wissenschaftler an. Sie zogen die Fische acht Monate lang außerhalb des Wassers auf und machten dabei bemerkenswerte Beobachtungen: Im Vergleich zur im Wasser aufgezogenen Kontrollgruppe setzten die Fische ihre Flossen näher an der Körpermitte auf und machten kürzere Schritte, wodurch sie weniger abrutschen. Außerdem hielten sie ihren Kopf höher und bewegten Flossen und Schwanz weniger, was die Fortbewegung verbessert und Energie spart. Das veränderte Verhalten sei wahrscheinlich durch das häufige Üben erlernt, so die Forscher. Noch erstaunlicher sind Veränderungen in der Anatomie. Schlüsselbeine und Cleithrum (ein Knochen des Schultergürtels von Knochenfischen und einiger Vierbeiner) waren verlängert, wodurch der Platz zwischen dem Kiemendeckel und der Flosse vergrößert wird, was der Flosse mehr Bewegungsraum verleiht. Dies stellt ein typisches Beispiel von Plastizität dar. Interessanterweise schwimmen die auf Land aufgezogenen Fische nicht schlechter als die im Wasser aufgezogene Kontrollgruppe.
Die bei den Flösselhechten beobachteten Veränderungen ähneln den Ausprägungen bei einigen fossilen Formen, die als Vorläufer der Landwirbeltiere diskutiert werden (s. u.). Daher spekulieren die Forscher, dass auch bei diesen Formen zur Zeit des Devons (vor ca. 400 Millionen radiometrischen Jahren) die beim Flösselhecht beobachtete phänotypische Plastizität eine Rolle gespielt haben könnte.
Ein Beitrag zur Erklärung von Makroevolution? Das Experiment von Standen et al. (2014) liefert interessante Ergebnisse. Es erinnert an ein unfreiwilliges Experiment mit einer Ziege, die von Geburt an nur Vorderbeine hatte, es aber lernte damit umzugehen, was ebenfalls auch anatomische Veränderungen zur Folge hatte (Slijpers Ziege; Beschreibung z. B. bei West-Eberhard 2005, 611). Kirschner & Gerhart (2005) nennen diese Fähigkeit von Geweben und Organen, auf äußere Reize flexibel reagieren zu können, „exploratives Verhalten“. Explorative Systeme sind „antwortend“, d. h. sie reagieren auf äußere Signale. Allerdings entstehen dadurch keine neuen Bauelemente und keine sonstigen Neuerungen wie z. B. neue Muskelansatzstellen oder neue Knochenelemente.
Zur Einschätzung der möglichen Bedeutung von Plastizität für Evolution, insbesondere zur Frage, wie evolutionäre Neuheiten entstehen könnten (Makroevolution), sollen nachfolgend einige Aspekte zusammengestellt werden:
- Die Änderungen betreffen keine Neuheiten, sondern sind Variationen vorhandener Konstruktionselemente und Verhaltensweisen. Die Flösselhechte besaßen anfangs bereits die Fähigkeit, auf Land zu gehen und auf Land zu überleben. Über den Ursprung dieser Fähigkeit geben die Experimente keinen Aufschluss.
- Die Fische würden das experimentell hervorgerufene Verhalten in ihrem natürlichen Lebensraum nicht zeigen. Dort graben sie sich bei Wasserknappheit in den Schlamm. Das im Experiment erzwungene Verhalten der Tier wurde unter natürlichen Bedingungen noch nicht beobachtet.
- Standen et al. (2014) ziehen Vergleiche der Knochenveränderungen mit dem fossilen Quastenflosser Eusthenopteron und mit den Tetrapoden (Vierbeinern) Acanthostega und Ichthyostega, die zu den ältesten Tetrapodengattungen gehörten. Eusthenopteron war ein reines Wassertier und seine Merkmalsausprägungen sind daher kaum geeignet, eine beginnende Landanpassung zu belegen. Ähnliches gilt auch für Acanthostega. Dieser war zwar ein Vierbeiner und besaß acht Finger, wird bisher aber aufgrund verschiedener anatomischer Merkmale ebenfalls als ausschließlich im Wasser lebend interpretiert (Clack 2002, 124). Aussagekräftiger ist dagegen der Vergleich mit Ichthyostega, da diese Gattung sehr wahrscheinlich auch an Land ging und klassisch als das „erste Amphibium“ gilt. Dennoch: Ichthyostega war sehr ungewöhnlich konstruiert mit stark überlappenden Rippen und ist in dieser und anderer Hinsicht ganz einzigartig gebaut (Ahlberg et al. 2005). Diese Autoren betrachten Ichthyostega als eines von mehreren kurzlebigen evolutionären Experimenten; diese Gattung wäre daher ebenfalls kein idealer Kandidat für eine Modellierung des Übergangs von Wasser ans Land.
- Für die anatomischen Veränderungen, die fossilen Formen entsprechen, welche als Vorläufer der Vierbeiner diskutiert werden, gibt es eine funktionelle Notwendigkeit. Die Ähnlichkeiten sind daher wenig überraschend.
- Für einen Übergang vom Leben im Wasser zu einem Leben an Land sind zahlreiche sehr viel grundlegendere Änderungen notwendig als die hier vorgestellte Plastizität, die bei den Flösselhechten beobachtet wurde.
- Hutchinson (2014) weist darauf hin, dass Flösselhechte trotz ihrer Plastizität keine landlebenden Nachfahren haben.
- Ob die plastisch aufgetretenen Merkmale bei Polypterus an die Nachkommen weitergegeben werden, ist derzeit nicht geklärt; das soll eine Fortsetzung des Experiments über mehrere Generationen zeigen. Plastische Änderungen gehen bei Wegfall der betreffenden Bedingungen wieder verloren; sie müssen im Laufe vieler Generationen fixiert werden, um evolutive Bedeutung zu erlangen. Ob dies bei Polypterus möglich ist, ist nicht bekannt; Pennisi (2014) meint, das sei ein „Geheimnis“.
- Zahlreiche Studien zur Plastizität der Lebewesen zeigen übereinstimmend, dass es einen Trend von anfangs hoher Plastizität in Richtung verringerter Plastizität gibt (Überblick bei Junker 2014). Diese Beobachtung machten die Forscher auch bei den außerhalb des Wassers aufgezogenen Flösselhechten: Ihre Gehbewegungen weisen eine verringerte Variabilität auf (Standen et al. 2014, 56). Interessant wird hier sein, welche Tendenz sich bei den geplanten Mehr-Generationen-Experimenten zeigen wird.
- Der Ursprung der Plastizität ist hier, wie auch in anderen Fällen unbekannt (Hutchinson 2014); klar ist aber, dass sie im normalen Lebensraum der Flösselhechte nützlich sind.
Schlussfolgerungen. In der Tagespresse wurden die Befunde zur Plastizität der Flösselhechte teilweise stark überbewertet. Eine der Überschriften lautete: „Evolution in acht Monaten: Wie Fische laufen lernen“.1 Was bisher Theorie gewesen sei, sei nun in der Praxis ansatzweise nachgewiesen worden. Das ist irreführend und auch keineswegs die Aussage des Originalartikels. Standen et al. (2014, 54) schreiben vielmehr: „Unsere Resultate eröffnen die Möglichkeit, dass umweltinduzierte Entwicklungs-Plastizität die Entstehung der Land-Merkmale, die zu den Vierbeiner führten, erleichtert haben.“2 Ob sie wirklich zu dieser Entstehung beigetragen haben, ist angesichts der oben genannten Befunde und Probleme mehr als fragwürdig.
Anmerkungen
2 „Our results raise the possibility that environmentally induced developmental plasticity facilitated the origin of the terrestrial traits that led to tetrapods.“
Literatur
Ahlberg PE, Clack JA & Blom H (2005) The axial skeleton of the Devonian tetrapod Ichthyostega. Nature 437, 137-140.
Clack JA (2002a) Gaining Ground. The origin and evolution of tetrapods. Bloomington and Indianapolis: Indiana University Press.
Hutchinson J (2014) Dynasty of the plastic fish. Nature 513, 37-38, doi:10.1038/nature13743.
Junker R (2014) Plastizität der Lebewesen: Baustein für Makroevolution? W+W Special Paper B 14-2. http://www.wort-und-wissen.de/artikel/sp/b-14-2-plastizitaet.pdf
Kirschner MW & Gerhart JC (2005) The Plausibility of Life. Resolving Darwin’s Dilemma. New Haven and London.
Pennisi E (2014) Fish raised on land give clues to how early animals left the seas. http://news.sciencemag.org/biology/2014/08/fish-raised-land-give-clues-how-early-animals-left-seas.
Standen EM, Du TY & Larsson HCE (2014) Developmental plasticity and the origin of tetrapods. Nature 513, 54-58; doi:10.1038/nature13708
West-Eberhard MJ (2005a) Phenotypic accommodation: Adaptive innovation due to developmental plasticity. J. Exp. Zool. 304B, 610-618.
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n217.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
18.07.14 Neuer Text über die kambrische Explosion
Ein neuer ausführlicher Artikel ist seit kurzem als PDF verfügbar: https://www.wort-und-wissen.org/artikel/zur-neueren-diskussion-ueber-die-kambrische-explosion/
Im unteren Teil der erdgeschichtlichen Periode des Kambriums ist eine große Anzahl von Tiergruppen in enormer Verschiedenartigkeit fossil überliefert, während vergleichbare Formen oder passende Vorstufen in älteren Schichten des Präkambriums weitgehend fehlen. Diese markante fossile Diskontinuität – das ziemlich abrupte Erscheinen einer vielgestaltigen Tierwelt – wird als „kambrische Explosion“ bezeichnet.
Es ist mittlerweile zwar eine erhebliche Anzahl von Vielzellern auch im Präkambrium entdeckt worden, doch handelt es sich dabei häufig um Formen, die nicht als Vorstufen der kambrischen Tierwelt gewertet werden, sondern eigene Linien repräsentieren, die früh ausgestorben sind (v. a. die Ediacara-Fauna). Die präkambrischen Formen erscheinen ihrerseits in drei Faunenvergesellschaftungen ebenfalls relativ abrupt. Auch die vielfältigen winzig kleinen (< 2mm) beschalten Fossilien im untersten Kambrium (sog. „small shelly-Fauna“) eignen sich nur teilweise als mögliche Vorläufer; für ihr Auftreten können ökologische Gründe wahrscheinlich gemacht werden. In den unterkambrischen Stufen des Tommotiums und Atdabaniums (Stufen 2 und 3 in der neueren Nomenklatur) nimmt die Verschiedenartigkeit der Formen rasant zu; diese Phase von ca. 6-10 Millionen radiometrischen Jahren wird meistens als eigentliche kambrische Explosion bezeichnet. Radiationen im jüngeren Kambrium und Ordovizium führen zu keiner markanten Erhöhung der Verschiedenartigkeit der Tierstämme mehr; während des Kambriums wird in der Regel bereits das Maximum an Verschiedenartigkeit der Tierstämme erreicht. Nur relativ wenige Formen werden als mögliche Vorläufer der kambrischen Tierstämme diskutiert, ihre Natur ist meist umstritten. Eine Anzahl neuerer Fossilfunde vergrößert die kambrische Vielfalt noch weiter.
Worauf die kambrische Explosion zurückzuführen ist, konnte im Rahmen evolutionärer Modelle bisher nicht geklärt werden. Diskutiert werden genetische, ökologische und geologische Ursachen. Die Vielfalt der nachweisbaren Baupläne erfordert das Vorhandensein entsprechender genetischer Grundlagen. Ein Zusammenhang mit dem Auftreten von Hox-Genen oder mit Genduplikationen ist eher unwahrscheinlich. Häufig werden zahlreiche Neuverschaltungen von Genen als Ursache angenommen, doch ist experimentell nicht gezeigt, dass auf diese Weise neue Bauplanelemente entstehen können; im Gegenteil ist bekannt, dass Änderungen in Kernbereichen von Gen-Regulations-Netzwerken zu schweren Schädigungen führen. Dass dies zur Zeit des Kambriums grundlegend anders gewesen sein könnte, ist spekulativ und wenig plausibel. Zudem kann allgemein aus der Art der Unterschiede zwischen verschiedenen Organismen nicht auf den Modus ihrer Entstehung (und auch nicht auf Evolution) geschlossen werden, da Unterschiede keinen Mechanismus begründen können. Aufgrund von Genvergleichen heute lebender Formen verschiedener Tierstämme wird evolutionstheoretisch allgemein ein genetisch komplexer Vorfahre angenommen. Wie dieser hypothetische Organismus entstanden ist, ist unbekannt. Weiter stellt sich die Frage, wofür er die vielen Gene primär benötigte, die bei den später entstandenen kambrischen Tierstämmen für die Ausbildung von Organen genutzt werden, die beim gemeinsamen genetisch komplexen Vorfahren noch nicht existiert haben können.
Die Auffassung, der Fossilbericht des Kambriums und Präkambriums könnte sehr unvollständig sein (Artefakt-Hypothese), ist angesichts zahlreicher für Fossilisierung geeigneter Sedimentgesteine* sowie des Vorkommens von sehr kleinen Fossilien und von fossilisierten Weichteilen unplausibel. Molekulare Daten (Gensequenzvergleiche von Tieren aus verschiedenen heutigen Tierstämmen) lassen zwar den Schluss auf eine mehr oder weniger tief präkambrische Entstehung der Tierstämme zu (die dann erst im Kambrium fossil in Erscheinung treten). Daraus resultiert jedoch keine Erklärung für die Entstehung der kambrischen Formen, sondern die Befunde dokumentieren eine Diskrepanz zwischen morphologisch-paläontologischen und molekularen Daten.
Neben genetischen werden auch ökologische und geologische Veränderungen als Auslöser der kambrischen Explosion in Betracht gezogen. Aber auch diese beantworten nicht die offenen Fragen, sondern liefern lediglich ein Spektrum der zu berücksichtigenden Begleitumstände. So ermöglichen weder ein freier Raum, abiotische Faktoren wie Klimaänderungen oder ein hypothetisches Räuber-Beute-Wettrüsten an sich eine Evolution der Baupläne, sondern könnten allenfalls als Triebfedern dafür gewertet werden. Davon abgesehen sind alle Vorschläge für ökologische und / oder geologische Auslöser umstritten.
Zahlreiche Belege und ausführliche Begründungen dieser zusammenfassenden Aussagen finden sich im Artike: https://www.wort-und-wissen.org/artikel/zur-neueren-diskussion-ueber-die-kambrische-explosion/.
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n215.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
29.04.14 Ein weiterer Baustein der kambrischen Explosion: Gliederfüßer als Filtrierer
Die kambrischen Gesteinsschichten bergen eine große Anzahl verschiedenster Baupläne, die recht abrupt in der Fossilüberlieferung erscheinen. Die dort auch vertretenen Anomalocariden galten bisher als furchterregende Räuber. Nun hat sich herausgestellt, dass einige von ihnen Filtrierer waren, die Kleinstlebewesen aus dem Wasser siebten. Damit sind sie ein weiterer Baustein der kambrischen Tier-Vielfalt und wieder einmal ein Beispiel für Konvergenz – die mehrmalige unabhängige Entstehung eines komplexen Bauplanelements.
Im Jahr 1989 beschrieb der unvergessene Paläontologe Stephen J. Gould in seinem Buch „Wonderful Life“ (deutsch 1991) die ungewöhnliche Tierwelt des Kambriums, besonders des mittelkambrischen Burgess-Schiefers in Britisch-Kolumbien (Kanada). Unter dessen Fossilien klassifizierte er 20 neue Tierstämme mit großen Bauplan-Unterschieden, die er als „irre Wundertiere“ bezeichnete; die äußerst vielgestaltigen Tiere nannte er „einzigartige Gliederfüßer“ mit einem „Maximum an anatomisch leistungsfähigen Möglichkeiten“. Seiner Meinung nach übertreffen die Burgess-Fossilien „wahrscheinlich das gesamte Spektrum des wirbellosen Lebens in den heutigen Ozeanen“.
Zu diesen „irren Wundertieren“ gehören die Anomalocariden („ungewöhnliche Garnelen“). Mit einer Länge von bis zu zwei Metern waren die unter den größten Tieren der kambrischen Gewässer. Sie werden zu den Gliederfüßern (Arthropoda) gerechnet. Mit ihren Seitenlappen am segmentierten Rumpf waren sie wahrscheinlich gute und wendige Schwimmer.
Die Anomalocariden galten bislang als formidable Räuber, doch neuere Untersuchungen stellen das teilweise in Frage und liefern ein überraschendes Ergebnis. Neue Fossilfunde aus Nordgrönland legen nahe, dass einige Arten der Anomalocariden sich mit Hilfe einer ausgeklügelten Filtriertechnik von Plankton ernährten – ähnlich wie die heutigen Bartenwale wie z. B. der Blauwal. Untersucht wurden fünf fossil erhaltene, stachelbewehrte, ca. 12 cm lange Anhänge und zwei mit dem Kopfschild verbundene Anhänge von Tamisiocaris borealis aus der unterkambrischen Sirius-Passet-Konservat-Lagerstätte (Bild einer Rekonstruktion hier: http://d.ibtimes.co.uk/en/full/1370642/tamisiocaris.jpg). Dabei wurden die beiden langen, stachelbewehrten Anhänge am Kopf genauer unter die Lupe genommen. Die Greifer erwiesen sich als beweglich und die filigranen Fortsätze besitzen feine, ineinander greifende Stacheln. Diese interpretieren die Forscher als Werkzeuge, mit deren Hilfe Tamisiocaris Organismen ab einer Größe von etwa ½ Millimeter aus dem Wasser gefischt hat (Vinther et al. 2014). „Ausgestreckt bilden sie eine Art Kamm, den der Urzeit-Gliederfüßer durch das Wasser gezogen haben könnte wie eine Art Netz. Rollte er dann die Anhänge zum Mund hin ein, bildeten ihre Borsten einen dichten Käfig, in dem gefangene Organismen hängenblieben“ (Podbregar 2014).

Abb. 1: Grafische Darstellung der Vielfalt der Größen und Ernährungsgewohnheiten bei Radiodonta (zu denen auch die Familie Anomalocarididae gehört). (Wikimedia: Rudy Lerosey-Aubril and Stephen Pates – https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6138677/, CC BY 4.0)
Aufgrund der Filtrierer-Fähigkeit stellen die Forscher Tamisiocaris in die neue Gruppe der Cetiocaridae (Walgarnelen; cetus, lat. = Wal; „caris“ steht für Garnele). Die Autoren schließen, dass ein vielfältiges Ökosystem mit gut entwickelten Nahrungsketten in den offenen Meeren ausgeprägt gewesen sei.1 Denn um als Filtrierer leben zu können, ist eine hohe Primärproduktion und ausreichend Plankton Voraussetzung. Vinther et al. (2014) geben auch eine Übersicht über die Vielfalt des Baus der Anhänge der Anomalocariden, z. B. solche mit dreizackigen Stacheln oder mit scherenartigen Anhängen und stellen fest (S. 498), dass die außerordentliche morphologische Vielfalt der Anhänge zeige, dass es sich bei den Anomalocariden nicht um ein gescheitertes Evolutionsexperiment handle, sondern um eine umfangreiche Radiation im Laufe der kambrischen Explosion.2
Die Untersuchung ist in zweierlei Hinsicht bemerkenswert. Zum einen vergrößern die Beobachtungen an Tamisiocaris die Vielfalt an ausgeklügelten Einrichtungen, die bereits im Unterkambrium ohne erkennbare Vorfahren präsent waren. Zum anderen zeigen sie einmal mehr, dass komplexe Einrichtungen mehrfach (oft vielfach) unabhängig auftauchen – in diesem Fall die Fähigkeit der Nahrungsfiltrierung. Eine ganze Reihe von Filtrierern gibt es außer den bereits erwähnten Bartenwalen auch unter den Gliederfüßern; und die Filtrierer sind nun um ein interessantes Mitglied erweitert worden, das diese Fähigkeit unabhängig erworben haben muss (Konvergenz). Vinther et al. (2014) erwähnen, dass bei den Cetiocaridae ähnliche Einrichtung zum Filtrieren verwirklicht sind wie bei anderen Filtrieren unter den Gliederfüßern (z. B. den Rankenfußkrebsen, Cirripedia). Dabei muss bedacht werden, dass diese Fähigkeit sehr anspruchsvoll ist und viele Details zusammenstimmen müssen. Ein allmählicher Erwerb oder – wie man evolutionstheoretisch vermutet – ein Umbau von einer räuberisch lebenden Ausgangsform sind sehr fragwürdig. Denn um ernährungsmäßig über die Runden kommen zu können, muss die Filtrierfunktion gut ausgebaut sein. Und der Verlust des räuberischen Nahrungserwerbs kann selektiv wohl kaum begünstig werden, solange die neue Art des Nahrungsgewinns nicht ausreichend funktioniert.
Vinther et al. (2014, 499) dagegen werten die Existenz der filtrierenden Gattung Tamisiocaris und das Ausmaß unabhängiger (konvergenter) Entstehung ähnlicher Konstruktionen als Beleg für Vorhersagbarkeit und Kanalisierung der Evolution. Worin diese Kanalisierung aber bestehen soll, ist völlig unklar und wird nicht thematisiert. Die Annahme einer zielorientierten, sprich schöpferischen Verursachung ist viel naheliegender.
Literatur
Gould SJ (1989) Wonderful Life. New York: W. W. Norton & Co. (dt. 1991: Zufall Mensch. München – Wien)
Vinther J, Stein M, Longrich NR & Harper DAT (2014) A suspension-feeding anomalocarid from the Early Cambrian. Nature 507, 496-500.
Podbregar N (2014) Sanfte Riesen im Urzeitmeer. http://www.wissenschaft.de/erde-weltall/palaeontologie/-/journal_content/56/12054/3250966/Sanfte-Riesen-im-Urzeitmeer/.
Anmerkungen
1 „The Cambrian pelagic food web was therefore highly complex containing multiple trophic levels, including pelagic predators and multiple tiers of suspension feeders” (Vinther et al. 2014, 499).
2 „This extraordinary range of appendage morphologies shows that, far from being a failed experiment, anomalocarids staged a major adaptive radiation during the Cambrian explosion, evolving to fill a range of niches as nektonic predators, much like the later radiations of vertebrates and cephalopods, including suspension feeders“ (Vinther et al. 2014, 498).
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n210.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.01.14 Eichelwürmer – vom Kambrium bis heute unverändert
Eichelwürmer aus dem Mittelkambrium ähneln heutigen Formen in erstaunlichem Maße. Nachdem kürzlich in der Antarktis erstmals Eichelwürmer entdeckt wurden, die Röhren bilden, ist die Ähnlichkeit mit den ebenfalls röhrenbildenden fossilen Formen perfekt. Die zwischenzeitlich diskutierte Vorstellung, die fossilen Eichelwürmer könnten Bindeglieder zu den Flügelkiemern (Pterobranchia) sein, ist damit nach Ansicht der Entdecker nicht haltbar.
Anfang des Jahres wurden schon länger fossil bekannte wurmartige Organismen aus dem mittelkambrischen Burgess-Schiefer eindeutig als Eichelwürmer identifiziert (Caron et al. 2013; Gee et al. 2013). Die betreffende Art wurde als Spartobranchus tenuis klassifiziert und gleicht heutigen Formen aus der Gruppe der Harrimaniiden bis in Details.1 Sie sind damit auch ein Beispiel für Stasis – das nahezu unveränderte „Stehenbleiben“ eines Bauplans über geologische Zeiträume hinweg. Der Nachweis von Eichelwürmern im Kambrium vergrößert die ohnehin immense Vielfalt der während der kambrischen Explosion auftretenden Formen noch weiter (vgl. Kambrische Explosion, https://www.genesisnet.info/index.php?Artikel=42842&Sprache=de&l=1 und Kambrische Explosion: Darwins Dilemma, https://www.genesisnet.info/index.php?News=200 gelöst?).
Die Eichelwürmer (Enteropneusta) werden zusammen mit den Flügelkiemern (Pterobranchia) zu den Hemichordaten gestellt. Die Arten dieser auch als Kiemenlochtiere benannten Gruppe besitzen einen weichen, wurmähnlichen, aber innen ungegliederten Körper und sind äußerlich in Kopfschild, Kragen und Rumpf dreigeteilt. Die bis zu 10 cm langen Eichelwürmer leben am Meeresboden und graben Gänge. Sie bewegen sich im Meeresboden durch wellenförmige Muskelkontraktionen vorwärts und leben von im Schlamm enthaltenen organischen Partikeln mit Hilfe eines Nahrungstrichters am Vorderende. Da auch die Pterobranchia aus dem Kambrium bekannt sind, ist klar, dass der Ursprung der Hemichordaten entsprechend ins Unterkambrium verlegt werden muss (Caron et al. 2013, 503). Einen Unterschied schien es jedoch bei den fossilen Formen im Vergleich zu den heutigen zu geben. Bei den Fossilien wurden häufig faserige Röhren gefunden; in einem Fall war ein Eichelwurm mit dieser Röhre assoziiert. Daher wurde angenommen, dass die kambrischen Eichelwürmer anders als die heutigen zeitweise in Röhren lebten. Dieser Befund wurde von S. Conway Morris, einem der Bearbeiter so interpretiert, dass es sich hier um einen Hinweis handle, dass die fossilen Eichelwürmer Übergangsformen zu den Pterobranchia seien (http://www.bbc.co.uk/nature/21745029), die generell solche Röhren bilden. Es wurde allerdings auch diskutiert, dass auch die Eichelwürmer ursprünglich ebenso wie die Pterobranchia solche Röhren ausgebildet haben könnten, diese später aber verloren haben (Caron et al. 2013, 505).
Die letztere Deutung erhielt jüngst Unterstützung. Denn es wurden heute lebende Eichelwürmer in der Tiefsee der Antarktis entdeckt, die ebenfalls Röhren ausbilden und in eine neue Eichelwürmer-Familie, Torquaratoridae, gestellt werden (Halanych et al. 2013). Die Autoren nehmen in ihrem Artikel Bezug auf die mittelkambrischen Eichelwürmer, die ebenfalls Röhren bilden, und stellen die enorme Ähnlichkeit der kambrischen mit den heutigen Formen heraus. Der faserige Bau der fossilen Röhren konnte durch den Vergleich mit den heutigen Formen als Erhaltungsartefakt erklärt werden. Denn einige Röhren waren mit Sediment bedeckt, was ihnen ein geripptes Aussehen verleiht, das der fossilen Erhaltung ähnlich ist. Die Autoren schließen aus dem ähnlichen Röhren-Design zwischen S. tenuis und den antarktischen Torquaratoridae auf ähnliche Verhaltensweisen, die sich demnach über einen Zeitraum von etwa 500 Millionen Jahren erhalten hätten.
Damit sei die Vorstellung, dass S. tenuis ein Bindeglied zwischen den Eichelwürmern und Pterobranchiern sei, unglaubwürdig geworden. Die Autoren schreiben: „Da die Trennung zwischen Eichelwürmern und Pterobranchiern vor dem Mittelkambrium erfolgt sein muss, ist die Röhre von S. tenuis kein Vorläufer des Coeneciums [Röhren] der Pterobranchier“ (Halanych et al. 2013). Es sei anzunehmen, dass bereits der gemeinsame Vorfahre Röhren gebildet habe. Fossile Spuren dieses hypothetischen Vorfahren sind nicht bekannt.
Am Rande sei vermerkt, dass die Deutung der kambrischen Eichelwürmer als mögliche Bindeglieder in der Wissenschaftspresse mehrfach aufgegriffen wurde, nicht aber das nachfolgende Dementi.
Literatur
Caron JB, Conway Morris S & Cameron CB (2013) Tubicolous enteropneusts from the Cambrian period. Nature 495, 503-506.
Gee H (2013) Tubular worms from the Burgess Shale, Nature 495, 458-459.
Halanych KM, Cannon JT, Mahon AR, Swalla BJ & Smith CR (2013) Modern Antarctic acorn worms form tubes. Nature Communications 4, No. 2738, doi: 10.1038/ncomms3738.
Anmerkung
1„Es ist fast so, als wenn man ein Foto eines heutigen Exemplars
machen würde“ wird Christopher Cameron zitiert, der heutige Eichelwürmer
untersucht (http://www.cbc.ca/news/technology/story/2013/03/13/science-phallus-fossil-acorn-worm.html).
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n205.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
04.12.13 Frühe Verschiedenartigkeit
Eine systematische Studie über die Fossilabfolgen verschiedener Organismengruppen zeigt – entgegen evolutionstheoretischen Erwartungen – einen klaren Trend: Das größte Ausmaß der Verschiedenartigkeit1 der untersuchten Gruppen wird frühzeitig erreicht und nimmt im Laufe der Fossilüberlieferung ab.
In Charles Darwins Hauptwerk On the origin of species ist eine einzige Abbildung abgedruckt: Ein Baum mit allmählich zunehmender Verzweigung und zunehmender Verschiedenartigkeit1. Die aufgezweigten Formen werden also im Laufe der Zeit allmählich unterschiedlicher. Am Anfang sind die Unterschiede zwischen den aufgespaltenen Formen noch gering, doch mit der Zeit nimmt die Verschiedenartigkeit immer mehr zu. Diese Darstellung ist folgerichtig, wenn die Lebewesen durch graduelle Evolution entstanden sind. Die experimentelle Forschung der Evolutionsmechanismen unterstützt die Erwartung gradueller Veränderungen. Größere sprunghafte Änderungen (wie z. B. vier- statt zweiflügelige Taufliegen) kommen zwar auch vor, ihre Träger sind aber weniger konkurrenzfähig, falls sie überhaupt lebensfähig sind. Neuere Hypothesen, wonach selbst morphologische* Evolution sprunghaft durch Änderungen in der Verschaltung von (Regulations-)Genen ablaufen könne, sind weitgehend spekulativ, da nicht durch experimentelle Daten gedeckt.
*morphologische (= die Gestalt, den Bau betreffend)
Dementsprechend könnte man erwarten, dass die Fossilüberlieferung von Tier- und Pflanzengruppen einem sich allmählich verzweigenden Baum ähnelt. Doch wie die paläontologische Fachliteratur schon lange zeigt, trifft das nicht zu. Vielmehr scheinen die Stammbäume in gewisser Hinsicht auf dem Kopf zu stehen. Besonders ausgeprägt ist das bei der sogenannten „kambrischen Explosion“ der Fall (Valentine 2004, Erwin & Valentine 2013, vgl. Kambrische Explosion: Darwins Dilemma gelöst?, https://www.genesisnet.info/index.php?News=202). Nun haben Hughes et al. (2013) diesen Befund durch eine umfangreiche Studie bestätigt.
Die Autoren führten eine Meta-Analyse von 98 Vielzellergruppen durch und ermittelten das Ausmaß an Verschiedenartigkeit der Gruppen im Laufe der Zeit. Sie stellten fest, dass eine früh etablierte große Verschiedenartigkeit das vorherrschende Muster während des gesamten Phanerozoikums2 ist, d. h. seit Beginn der fossilen Vielzeller-Überlieferung (kambrische Explosion) bis zur Gegenwart. Dabei nimmt die Verschiedenartigkeit nach einem frühen Maximum im Laufe der Zeit wieder ab. Es sei auch kein Trend in den Verschiedenartigkeitsprofilen während dieser gesamten Zeitspanne erkennbar. Ausnahmen von diesem Befund gebe es nur bei Massenaussterbeereignissen sowie bei Formengruppen, die sich erst in jüngerer erdgeschichtlicher Zeit entfaltet haben und gegenwärtig noch existieren.
Dieser Befund steht den evolutionstheoretischen Erwartungen entgegen. Als Erklärung schlagen die Autoren vor, die schnell etablierte große Verschiedenartigkeit sei Folge der Evolution von Schlüsselneuheiten. Weniger wahrscheinlich seien dagegen Änderungen, die Umweltbedingungen als Ursache hatten, oder die durch katastrophische Umweltveränderungen eingetreten seien.
Kommentar. Alle genannten Erklärungsversuche sind jedoch ungeeignet, da sie nur als Begleiterscheinungen und nicht als Ursachen gelten können. Mit dem Nachweis von Begleitumständen ist aber niemals eine kausale Begründung gegeben. Wenn z. B. das Auftreten evolutionärer Neuheiten mit großer Verschiedenartigkeit am Beginn der Entfaltung einhergeht, resultiert daraus keine Erklärung, woher diese Neuheiten kommen und wie sie entstanden sind. Vielmehr wird das Ausmaß an Verschiedenartigkeit gerade an der Zahl unterschiedlicher Baupläne gemessen, die in evolutionstheoretischer Perspektive natürlich mit dem Auftreten von Neuheiten korreliert. Darin eine Ursache für das Phänomen der schnell etablierten Verschiedenartigkeit zu sehen, wäre ein Zirkelschluss.
Dass die Verschiedenartigkeit im Laufe der Zeit tendenziell eher abnimmt, könnte – so die Autoren – mit wachsenden Entwicklungszwängen (developmental constraints) zusammenhängen. Das heißt, je mehr die verschiedenen Organe der Lebewesen im Laufe der Zeit miteinander in Wechselwirkung treten, desto weniger konnte das Gesamtgefüge geändert werden. Auch diese Vorstellung ist spekulativ, darüber hinaus wenig plausibel, denn evolutionstheoretisch gesehen müssten vielfach Konstruktionszwänge wieder aufgebrochen worden sein. Wie dies funktioniert und wie anschließend auf diese Weise immer wieder vergleichsweise schnell eine große Verschiedenartigkeit relativ frühzeitig in der (Fossil-)Geschichte von Organismengruppen etabliert wurde, ist unbekannt. Zudem ist unbekannt, wie komplexere Lebewesen mit deutlich geringeren Konstruktionszwängen. existieren könnten.
Anmerkungen
1 Mit Verschiedenartigkeit (disparity) wird die morphologische Variation, also die Unterschiedlichkeit von Bauplänen innerhalb einer größeren Formengruppe bezeichnet, im Gegensatz zur Vielfalt (diversity) innerhalb eines engen Verwandtschaftskreises. Beispielsweise würde man zehn Hunderassen als vielfältig bezeichnen, dagegen zehn sehr verschiedene Säugetierarten (etwa von Fledermaus bis Blauwal) als verschiedenartig.
2 Das Phanerozoikum umfasst alle geologischen Systeme, die in nennenswertem Umfang Vielzeller-Fossilien bergen, das sind alle Systeme vom Kambrium bis heute. „Phanerozoikum“ bedeutet „das Leben erscheint“ (in fossil erhaltener Form). Mittlerweile sind auch im jüngeren Präkambrium eine Reihe von Vielzellergruppen entdeckt worden; diese zeigen im Großen und Ganzen jedoch wenige Ähnlichkeiten mit den Fossilien seit dem Kambrium.
Literatur
Erwin D & Valentine JW (2013) The Cambrian explosion. The construction of animal biodiversity. Greenwood Village, Colorado.
Hughes M, Gerber S & Wills MA (2013) Clades reach highest morphological disparity early in their evolution. Proc. Natl. Acad. Sci. 110, 13875-13879.
Valentine JW (2004) On the origin of phyla. Chicago and London.
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n204.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
13.11.13 Kambrische Explosion: Darwins Dilemma gelöst?
Das plötzliche fossile Erscheinen verschiedenster Tierstämme und vieler ihrer Untergruppen im Kambrium („kambrische Explosion“) wird auch als „Darwins Dilemma“ bezeichnet. Die Ergebnisse einer Studie über die Geschwindigkeit der Evolution der Gliederfüßer zur Zeit des Kambriums sollen Darwins Dilemma auflösen. Diese Behauptung ist jedoch durch die präsentierten Daten aus verschiedenen Gründen nicht gedeckt, vor allem da die Entstehung der Tierstämme gar nicht Gegenstand der Untersuchung war.
Das plötzliche fossile Erscheinen verschiedenster Tierstämme zu Beginn des Kambriums wird bis heute als „kambrische Explosion“ bezeichnet (Erwin & Valentine 2013). Für Charles Darwin war dieser Befund ein Problem, es wurde auch bekannt als „Darwins Dilemma“, denn ein abruptes Auftreten ganz verschiedener Baupläne passte nicht zu seinen Vorstellungen einer graduellen Evolution. Es gibt viele Versuche, dieses Problem zu lösen oder wenigstens zu entschärfen – ohne befriedigendes Ergebnis (Meyer 2013). Nun behaupten Lee et al. (2013), dass die Resultate einer von ihnen durchgeführten Studie über die Geschwindigkeit der Evolution zur Zeit des Kambriums Darwins Dilemma auflösen könnten. Sie ermittelten, unter Voraussetzung von Evolution, evolutionäre Veränderungsraten beim größten Tierstamm, den Gliederfüßern (Arthropoda; dazu gehören vor allem Insekten, Spinnentiere und Krebse). Es wurden Evolutionsraten von verschiedenen Merkmalen ermittelt: Sowohl von morphologisch-anatomischen (gestaltlichen, phänotypischen) als auch molekularen (genetischen). Die Änderungsraten dieser Merkmale während des Kambriums verglichen sie mit den Raten in nachfolgenden Zeiträumen und gelangten zum Ergebnis, dass es im Kambrium eine ca. 4-fach schnellere phänotypische und ca. 5,5-fach schnellere molekulare Evolution gab, verglichen zu Veränderungen in jüngeren Abschnitten des Phanerozoikums1. Für manche Tiergruppen innerhalb der Gliederfüßer ermittelten sie eine bis zu 10-fach erhöhte molekulare und bis zu 16-fach höhere morphologische Änderungsrate im Kambrium.
Die Ergebnisse basieren auf 395 morphologischen und 62 molekularen Merkmalen der heute lebenden Gliederfüßer. Die zeitliche Einordnung der mutmaßlichen Gabelungspunkte der jeweiligen Gruppen erfolgte anhand von Fossilfunden. Mit dieser Datenbasis konnten unter Voraussetzung von Evolution die Evolutionsraten in den verschiedenen Zeiträumen abgeleitet werden.
Die Autoren kommen zum Schluss, dass auch noch die höchsten Evolutionsraten im Einklang mit der Entstehung durch natürliche Selektion stünden, womit Darwins Dilemma letztlich gelöst sei. Diese Einschätzung ist insofern überraschend, als ihre Untersuchungen überhaupt nichts zu den Mechanismen der Entstehung der kambrischen Baupläne aussagen, unabhängig davon, wie schnell die Veränderungen abgelaufen sind. Welche Rolle natürliche Selektion hatte, kann aus den Daten nicht abgeleitet werden. Allenfalls kann eventuell aus den Daten gefolgert werden, dass die kambrische Explosion nicht so markant ist wie oft dargestellt.
Aber auch diese Schlussfolgerung ist durch die von Lee et al. präsentierten Daten nicht gedeckt – aus mehreren Gründen (Luskin 2013a): Zunächst betrifft die Untersuchung gar nicht den zentralen Aspekt der kambrischen Explosion, nämlich den Ursprung der Tierstämme und vieler Tierklassen. Vielmehr geht es in ihrer Untersuchung um Veränderungen innerhalb eines einzigen Tierstamms (der Gliederfüßer), nachdem dieser bereits entstanden ist. Die Entstehung der Stämme und die Geschwindigkeit dieser Evolution ist jedoch nicht der Gegenstand der Analysen von Lee et al. Die Unterschiede innerhalb von Tierstämmen sind aber deutlich geringer als zwischen verschiedenen Tierstämmen. Die Evolutionsraten innerhalb der Gliederfüßer wurden zudem durch Vergleiche heutiger Arten und mittels Eichung durch Fossilfunde erschlossen (s. u.). Raten innerhalb nur fossil bekannter Formen (wie z. B. Trilobiten) wurden nicht bestimmt, was z. B. vom Paläontologen Douglas Erwin kritisiert wird, denn diese Daten könnten Einfluss auf die ermittelten Evolutionsraten haben (Servick 2013).
Im Übrigen tauchen die Gliederfüßer fossil abrupt im Unterkambrium auf; dieser Sachverhalt wird durch die Studie von Lee et al. nicht berührt. Sie nehmen zwar an, dass die Gliederfüßer im Ediacarium, dem letzten geologischen System vor dem Kambrium entstanden sind, eindeutige fossile Belege dafür sind jedoch nicht bekannt. Unklar ist, welchen Zeitraum die Autoren genau unter „frühes Kambrium“ und „kambrische Explosion“ fassen, es scheint ein Zeitraum von etwa 40 Millionen Jahren zu sein (Lee et al. 2013, 1890). Die meisten Experten sind jedoch der Auffassung, dass sich die eigentliche „Explosion“ während nur 5-10 Millionen Jahren ereignete (Belege bei Luskin 2013b), was die von Lee et al. ermittelten Evolutionsraten entsprechend vervielfachen würde.
Es gibt noch einen ganz anderen Kritikpunkt an den Schlussfolgerungen, die aus den Ergebnissen von Lee et al. gezogen werden, nämlich die Auswahl der zugrundeliegenden Gene. Die Autoren haben die Datenbasis aus einer anderen Untersuchung entnommen, deren Ziel es war, die Abstammungs- bzw. Verwandtschaftsverhältnisse unter den sehr unterschiedlichen Gliederfüßern aufzuklären. Zu diesem Zweck sind (unter evolutionstheoretischen Vorgaben) solche Gene besonders geeignet, deren Änderungsraten möglichst klein sind, und das trifft auf Gene zu, die wichtige Grundfunktionen im Zellgeschehen haben (sog. „housekeeping genes“). Bei solchen Genen sammeln sich nur (nahezu) neutrale Mutationen an, die wenig Einfluss auf die Funktion haben, und dies geschieht relativ konstant während der Evolutionsgeschichte (sofern sich die Unterschiede überhaupt auf evolutivem Wege etabliert haben). Nun sind aber gerade solche Gene mit großer Wahrscheinlichkeit von der Entstehung neuer morphologischer Baupläne, wie sie während der kambrischen Explosion entstanden sind, kaum oder gar nicht beeinflusst. Viel wichtiger sind laut Evolutionstheoretikern vor allem Regulationsgene, um neue Baupläne zu erzeugen (vgl. Erwin & Valentine 2013, 2522). Daraus folgt, dass die Untersuchung von Evolutionsraten-Änderungen im Zuge der kambrischen Explosion mittels der von Lee et al. verwendeten 62 Gene denkbar ungeeignet ist.
Fazit: Die Studie von Lee et al. (2013) bietet weder Klärungen noch Hinweise auf die Ursachen der kambrischen Explosion; daher ist die Behauptung, die erhöhten Evolutionsraten im Kambrium, und damit auch die kambrische Explosion, sei mittels natürlicher Selektion erklärbar, durch die präsentierten Daten nicht gedeckt. Die Untersuchung legt den Fokus nicht auf die Entstehung der Stämme (die eigentliche „Explosion“), sondern auf Untergruppen innerhalb des Stamms der Gliederfüßer. Daher haben die Ergebnisse eine nur sehr eingeschränkte Aussagekraft zur kambrischen Explosion der Tierstämme.
Anmerkungen
1 Das Phanerozoikum umfasst alle geologischen Systeme, die in nennenswertem Umfang Vielzeller-Fossilien bergen, das sind alle Systeme vom Kambrium bis heute. „Phanerozoikum“ bedeutet „das Leben erscheint“ (in fossil erhaltener Form). Mittlerweile sind auch im jüngeren Präkambrium eine Reihe von Vielzellergruppen entdeckt worden; diese zeigen im Großen und Ganzen jedoch wenige Ähnlichkeiten mit den Fossilien seit dem Kambrium.
2 „When it comes to understanding the genetic bases of the morphological richness and disparity of the Cambrian explosion, our primary interest is in the genes involved in specifying the development of body plans and other morphological features. That is to say, it is not the genes that control basic cellular functions (so-called housekeeping genes) that are of interest, but the genes that regulate the development of morphology from egg to adult.“
Literatur
Erwin DH & Valentine JW (2013) The Cambrian explosion. The construction of animal biodiversity. Greenwood Village, Colorado.
Lee MSY, Soubrier J & Edgecombe GD (2013) Rates of Phenotypic and Genomic Evolution during the Cambrian Explosion. Curr. Biol. 23, 1889-1895.
Luskin C (2013a) Current Biology Paper’s Assumptions and Methodology Dramatically Underestimate „Rates of Change“ in the Cambrian Explosion. http://www.evolutionnews.org/2013/10/current_biology078581.html
Luskin C (2013b) How „Sudden“ Was the Cambrian Explosion? Nick Matzke Misreads Stephen Meyer and the Paleontological Literature; New Yorker Recycles Misrepresentation. http://www.evolutionnews.org/2013/07/how_sudden_was_074511.html
Meyer SC (2013) Darwin’s Doubt. The explosive origin of animal life and the case for intelligent design. New York.
Servick K (2013) Evolution’s Clock Ticked Faster at the Dawn of Modern Animals. Science NOW, http://news.sciencemag.org/evolution/2013/09/evolution%E2%80%99s-clock-ticked-faster-dawn-modern-animals
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n202.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
06.11.13 Gliederfüßergehirn: Von Beginn an „modern“
Einmal mehr zeigt sich, dass Organe von Tieren, die während der „kambrischen Explosion“ fossil auftauchen, in ihrer Komplexität heutigen Formen gleichen, diesmal beim Gehirn eines Gliederfüßers aus Chengjiang (Südwestchina) mit spinnentypischer Struktur.
Vergleichende Studien und Fossilfunde haben in der jüngeren Vergangenheit wiederholt nahegelegt, dass unter evolutionstheoretischen Voraussetzungen eine früh etablierte große Komplexität von Organen angenommen werden muss und dass sich deren Bau bis heute nicht mehr wesentlich verändert hat. Beispiele aus jüngerer Zeit sind das Gehirn von Fliegen (Strausfeld & Hirth 2012), das Nervensystem von Gliederfüßern (Ma et al. 2012) oder Komplexaugen (Lee et al. 2011). Erst kürzlich stellte sich heraus, dass das Gehirn des kambrischen Krebses Fuxianhuia in Proto-, Deutero- und Tritocerebrum gegliedert ist – genauso wie bei vielen heute lebenden Höheren Krebsen und Insekten. Nun bestätigt ein weiterer Gliederfüßer aus der Zeit der „Kambrischen Explosion“ (Kambrische Explosion) die frühe Etablierung eines modernen Gliederfüßer-Gehirns von Beginn der Fossilüberlieferung der betreffenden Tiergruppe an. Es handelt sich dabei um Alalcomenaeus sp. aus der berühmten unterkambrischen der Chengjiang-Lagerstätte in Südwestchina (Alter: ca. 520 Millionen radiometrische Jahre). Gengo Tanaka (Japan Agency for Marine-Earth Science and Technology, Yokosuka) und Mitarbeiter beschrieben dieses Fossil kürzlich in einem Nature-Artikel.
Der fossile ca. 3 cm lange Gliederfüßer gleicht einigen typischen Vertretern der damaligen Fauna. Sein Körper ist segmentiert und das Tier besitzt ein Dutzend Beinpaare mit leicht verbreiterten Anhängen, die sowohl ein Krabbeln auf dem Meeresgrund als auch das Schwimmen ermöglichten. Erhalten sind auch paarige Augen mit einem Durchmesser von je 0,75 mm und zusammengesetzt aus 15 Mikrometer großen Facetten. Am Kopf trug das Tier zwei mit großen Klauen ausgestattete Anhänge, aufgrund derer es zu den Megacheira („große Hände“) gestellt wird.
Bei Fossilien ist es allerdings schwer zu entscheiden, ob die Scheren vom Krebstyp oder vom Spinnentyp sind. Die Scheren beider Gruppen sind zwar ähnlich, entstehen ontogenetisch aber auf verschiedene Weisen und entsprechen verschiedenen Körperteilen. Bei Alalcomenaeus half jedoch die spinnentypische Struktur des Gehirns, eine Entscheidung bezüglich der Zugehörigkeit zu treffen. Es konnte nachgewiesen werden, dass die großen, paarigen Scheren vom gleichen Hirnteil kontrolliert wurden wie bei den heutigen Spinnentieren (Webspinnen, Weberknechte, Skorpione und Milben) und Pfeilschwanzkrebsen, die als Kieferklauen- oder auch Scherenträger (Chelicerata) zusammengefasst werden. Somit ist klar, dass die Megacheira als die bislang ältesten bekannten Vorfahren der heutigen Spinnentiere angesehen werden können.
Mittels Computertomografie und einer Laser-Scanning Technik konnten Tanaka et al. (2013) die Anatomie des Nervensystems von Alalcomenaeus genau untersuchen – dank der sehr guten Erhaltung des Fossils. Der Urzeit-Gliederfüßer aus dem Kambrium ist so gut erhalten, dass sein Gehirn und Nervensystem weitgehend intakt konserviert sind. Dabei zeigten sich große Ähnlichkeit mit Gehirn und Nervensystem der heutigen Cheliceraten („Scherenträger“). Die Ähnlichkeiten betreffen die Anordnung der paarigen optischen Neuropile (Sehzentrum), das Gehirn (eine Art Vorhirn sowie ein aus vier Nervenknoten verschmolzenes Gehirn) und die Ganglien (Tanaka et al. 2013, 364).

Abb. 1: Alalcomenaeus cambricus (links) und Fuxianhuia protensa (rechts). (Wikimedia: Captmondo, CC BY-SA 3.0; Jessica Utrup – https://www.gbif.org/occurrence/351363777, CC0)
Einmal mehr zeigt sich, dass Organe von Tieren, die während der „kambrischen Explosion“ fossil auftauchen, in ihrer Komplexität heutigen Formen gleichen. Mit Alalcomenaeus und Fuxianhuia sind nun aus Chengjiang Fossilien überliefert, deren Gehirn modernen Vertretern der beiden Hauptgruppen der Gliederfüßer ähnelt. Die Autoren stellen fest: „Alalcomenaeus und Fuxianhuia protensa zeigen, dass die beiden Hauptkonfigurationen des Gehirns, wie wir sie bei modernen Gliederfüßern kennen, nämlich der Chelicerata und Mandibulata, im frühen Kambrium evolviert waren.“1 Damit ist auch klar, dass diese beiden Gruppen bereits im Unterkambrium getrennt waren, denn Alalcomenaeus und Fuxianhuia lebten gleichzeitig, sind aber den beiden verschiedenen Gruppen zuzuordnen.
Anmerkung
1 „Alalcomenaeus and Fuxianhuia protensa demonstrate that the two main configurations of the brain observed in modern arthropods, those of Chelicerata and Mandibulata, respectively, had evolved by the early Cambrian“ (p 364). Zu den Mandibulata gehören Krebse, Insekten und Tausendfüßer; sie gehören zusammen mit den Chelicerata zu den Gliederfüßern.
Literatur
Lee MSY, Jago JB, García-Bellido DC, Edgecombe GD, Gehling JG & Paterson JR (2011) Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia. Nature 474, 631-634.
Ma X, Hou X, Edgecombe GD & Strausfeld NJ (2012) Complex brain and optic lobes in an early Cambrian arthropod. Nature 490, 258-261.
Strausfeld NJ & Hirth F (2012) Deep Homology of Arthropod Central Complex and Vertebrate Basal Ganglia. Science 340, 157-161.
Tanaka G, Hou K, Ma X, Edgecombe GD & Strausfeld NJ (2013) Chelicerate neural ground pattern in a Cambrian great appendage arthropod. Nature 502, 364-367.
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n201.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.10.13 Fisch mit Gesicht stellt Abstammung auf den Kopf
Bestätigen neue Fossilfunde immer wieder Evolution? Der Fund eines Fischfossils aus dem oberen Silur Chinas stellt eine etablierte Abstammungsvorstellung auf den Kopf: Knochenfische sollen nun vor den Knorpelfischen entstanden sein. Das hätte allerdings evolutionstheoretisch zur Folge, dass ein knöcherner Kiefer zweimal unabhängig entstand oder dass die Entstehung der Knorpelfische mit zahlreichen Rückbildungen einherging. Beides ist nicht gerade plausibel.
Die Einschätzung, dass eine allgemeine Evolution der Lebewesen eine Tatsache sei, wird häufig damit begründet, dass neue Befunde sich immer wieder in das evolutionstheoretische Theoriegebäude einfügen ließen und aus Evolutionstheorien abgeleitete Vorhersagen sich erfüllten. So wird beispielsweise auf passende fossile Übergangsformen verwiesen. Tatsächlich aber entsprechen viele neue Fossilfunde jedoch gerade nicht den evolutionstheoretisch motivierten Erwartungen.
Ein jüngst entdecktes Fossil verlängert die Reihe evolutionstheoretisch völlig unerwarteter Formen. Von diesem Fund, der den Stammbaum der Wirbeltiere kräftig „durcheinanderwirbelt“ (http://www.dradio.de/dlf/sendungen/forschak/2266356/) berichteten Min Zhu und Kollegen von der Chinesischen Akademie der Wissenschaften in der Wissenschaftszeitschrift Nature. Es handelt sich dabei um das Fossil eines etwa 20 cm langen Fisches, der die etablierte Theorie über die stammesgeschichtlichen Beziehungen von Knochen- und Knorpelfischen komplett in Frage stellt (Zhu et al. 2013).
Bisher war man davon ausgegangen, dass Knorpelfische ähnlich den heutigen Haien Vorfahren der Knochenfische seien. Die beiden Gruppen bilden zusammen mit den ausgestorbenen Panzerfischen (Placodermi) und den Acanthodii die Kiefermäuler (Gnathostomata). Zu den Knorpelfischen gehören Haie, Rochen und Seekatzen. Sie besitzen keine Knochen; ihr Skelett ist aus Knorpel aufgebaut, der jedoch durch Kalkeinlagerung eine hohe Festigkeit erreicht, die die enorme Größe mancher Haie erlaubt. Richtiges Knochengewebe wird nur ganz selten bei großen alten Haien in den Wirbeln ausgebildet. Der Großteil der Fische (über 95 % der Arten) hat dagegen ein knöchernes Skelett, das als evolutionäre Weiterentwicklung betrachtet wird. Im Kopfbereich besitzen die Knochenfische anders als die Knorpelfische feste Platten und hochentwickelte Kieferknochen, was den Fischen sozusagen ein „richtiges Gesicht“ verleiht. Dagegen besitzen die Knorpelfische im Schädelbereich nur winzige Schuppen.
Der Fund des neuen Fossils, das den Gattungsnamen Entelognathus erhielt, stammt aus dem oberen Silur der Yunnan-Provinz Chinas und wurde auf 419 Millionen Jahre datiert. Entelognathus ähnelt mit seiner knochigen Schulter- und Schädelpartie den Panzerfischen, die als frühe Abspaltung der Linie gelten, aus denen sich Knorpel- und Knochenfische entwickelt haben sollen. Es stellte sich jedoch heraus, dass die neu entdeckte Art auch differenzierte Kieferknochen besaß, wie sie bisher nur von Knochenfischen bekannt sind, ein komplexes Arrangement kleinerer Knochen, die als Prämaxilla und Maxilla im Oberkiefer, als Dentale im Unterkiefer und als Wangenknochen bekannt sind (Zhu et al. 2013). Es handelt sich also um einen Fisch mit Gesicht, der damit der älteste bekannte Fisch mit Gesicht ist.

Abb. 1: Künstlerische Rekonstruktion von Entelognathus primordialis. (Wikimedia: Entelognathus, CC BY-SA 4.0)
Das hat weitreichende Folgen für die stammesgeschichtliche Einordnung: Entelognathus könnte zu einer Schwestergruppe der Panzerfische gehören oder eng mit dem letzten gemeinsamen Vorfahren von Knorpel- und Knochenfischen verwandt sein (siehe Abbildung http://www.nature.com/nature/journal/vaop/ncurrent/images_article/nature12690-f1.jpg). In jedem Fall kann der hochentwickelte Kiefer nicht mehr als Alleinstellungsmerkmal der Knochenfische angesehen werden. Der knöcherne Kiefer als Struktur könnte somit zweifach unabhängig (konvergent) entstanden sein. Oder aber, und dazu scheinen die Wissenschaftler zu neigen, die Schädelknochen sind in der Knorpelfisch-Linie wieder verlorengegangen, wenn die Knochenfische vor den Knorpelfischen entstanden sein sollen (Friedman & Brazeau 2013). Beide Möglichkeiten sind wenig plausibel – nicht umsonst wurde die Stammesgeschichte der beiden großen heute lebenden Fischgruppen bisher genau anders herum rekonstruiert: Knorpelfische vor Knochenfischen.
{kind=link}
Unerwartete Funde und Evolution
Auf der Basis etablierter stammesgeschichtlicher Hypothesen war das neue Fossil mit der Kombination von Merkmalen aus Placodermen und Knochenfischen nicht vorhergesagt worden. Es erfordert die Annahme einer Umkehr der Evolutionsrichtung gegenüber den bisher favorisierten Hypothesen. Natürlich kann die (Re-)Konstruktion von Stammbäumen immer an neue Fossilfunde angepasst werden; Fossilien können die Evolution als konzeptionelle Vorgabe nicht „umstürzen“. Aber die Anpassung der Theorie an die Befunde erfordert, hier wie in vielen anderen Fällen, eine Verkomplizierung evolutionstheoretischer Hypothesen: Entweder muss eine zweimal unabhängige Entstehung von knöchernem Kiefer und Knochenplatten oder deren Verlust bei den Knorpelfischen angenommen werden. Man kann schwerlich behaupten, dass der neue Fund das evolutionäre Paradigma einmal mehr bestätigt hätte.
Literatur
Zhu M, Yu X et al. (2013) A Silurian placoderm with osteichthyan-like marginal jaw bones. Nature, doi:10.1038/nature12617.
Friedman M & Brazeau MD (2013) A jaw-dropping fossil fish. Nature, doi:10.1038/nature12690.
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n199.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
25.01.13 Modernes Denken im Kambrium
Nicht nur bei Fossilien von vermuteten Vorfahren des Menschen ist es schwierig, die Leistungsfähigkeit des Gehirns einzuschätzen. Größe allein zum Beispiel kann spätestens seit dem Fund des sogenannten „Hobbit-Menschen“ (Homo floresiensis) kein verlässlicher Indikator mehr sein, die strukturelle Architektur hingegen lässt eher Aussagen zur Leistungsfähigkeit zu (falls Schädel-Innenabdrücke vorhanden sind). Aber auch bei einfacheren Lebewesen, wie z. B. Krebstieren, war die Situation nicht leichter. Neue Beschreibungen ungewöhnlich gut erhaltener Fossilien sehr früher Gliederfüßer lassen allerdings auf ein bereits sehr modernes Nervensystem schon im Kambrium schließen.
Rückschlüsse auf Fähigkeiten oder die Organisation des Nervensystems bei frühen, fossil erhaltenen Gliederfüßern (Arthropoden, zu ihnen gehören Insekten, Spinnen, Krebstiere u. v. a.) zu ziehen, war bisher nicht einfach, man war auf indirekte Hinweise angewiesen. Ausgehend von der Gehirn-Organisation bei heute lebenden Krebstieren ging man davon aus, dass der letzte gemeinsame Vorfahr aller Gliederfüßer wahrscheinlich ein einfaches, zweigliedriges Gehirn aufwies, wie man es heute noch bei Branchiopoden (Kiemenfußkrebse, z. B. der Wasserfloh, Daphnia) findet. Fossile Funde von großen leistungsfähigen Komplexaugen bei sehr frühen, kambrischen Lebewesen (Anomalocaris) legten allerdings schon einige Zeit die Annahme nahe, dass die neurologischen Fähigkeiten ihrer Träger nicht wesentlich geringer sein konnten, als die der heutigen Vertreter, denn die Komplexaugen waren denen heutiger Lebewesen weder in Größe noch in der Form, noch in der optischen Auflösung unterlegen (Paterson et al. 2011; vgl. „Moderne Optik“ im frühen Kambrium).
Sehr gut erhaltene kambrische1 Fossilien von Gliederfüßern aus der südwestchinesischen Provinz Yunnan gewähren nun detaillierte Einblicke in die Organisation des Nervensystems von Lebewesen, die man als dem letzten gemeinsamen Vorfahren aller Gliederfüßer sehr nahe stehend einstuft (Fuxianhuia). Sie werden auf mehr als eine halbe Milliarde radiometrische Jahre datiert (520 Millionen Jahre). Schon an Fossilien des berühmten kanadischen Burgess Schiefers (ebenfalls Kambrium) konnte gezeigt werden, dass fossilisierte interne Strukturen sehr gut vergleichbar sind mit Gehirn und Ganglien (Ansammlung von Nervenzellen) heutiger Gliederfüßer. Offensichtlich konnte ihr Nervengewebe, obwohl es als sehr anfällig für schnellen Zerfall angesehen wird, in seltenen Fällen doch gut fossil erhalten werden. An den ca. 7,5 cm langen Fossilien der Art Fuxianhuia protensa aus China lässt sich die Gliederung dieses Nervensystems jetzt besonders gut studieren.
Ganglien und Nervenstränge sind in den in Kalkstein erhaltenen Fossilien als dunkle, eisenhaltige Strukturen erkennbar. Erstaunlicherweise ist das Gehirn von Fuxianhuia wie bei vielen heute lebenden Höheren Krebsen und Insekten dreigegliedert in Proto-, Deutero- und Tritocerebrum, und nicht nur zweigliedrig, wie eigentlich erwartet. Paarige Nervenstränge versorgen die auf Stielen sitzenden Komplexaugen, die dahinter liegenden Antennen und eine dritte, noch nicht eindeutig identifizierte paarige Struktur, wahrscheinlich Anhänge des Verdauungssystems. An der Basis der Stielaugen kann man insgesamt drei untereinander verbundene optische Neuropile erkennen, signalverarbeitende Nervenstrukturen mit einer hohen Dichte an Nervenfasern, die wiederum typisch sind für heutige höhere Krebse und Insekten. Sie fehlen dagegen bei den als primitiver eingestuften Kiemenfußkrebsen. Die relativ großen Augen besitzen einen gegliederten optischen Aufbau und sind offensichtlich wie bei heutigen Krebsen auf ihren Stielen gut beweglich. Außerdem haben diese Komplexaugen vorne einen größeren Radius als seitlich, was eine bessere Auflösung in die Hauptsehrichtung erlaubt, wiederum genau wie man es bei heutigen Insekten und Krebstieren findet. Der Körperbau von Fuxianhuia ist ansonsten deutlich unterschiedlich von heute lebenden Gliederfüßern, recht einfach, aus ca. 30 Segmenten aufgebaut, was dann mit dem Etikett „primitiv“ versehen wird, wobei die ersten Kopfsegmente von einem zweigeteilten Schild geschützt sind.

Abb. 1: Fuxianhuia protensa. (Wikimedia: Jessica Utrup – https://www.gbif.org/occurrence/351363777, CC0)
Die hohe Komplexität im Gehirn sehr früher, äußerlich einfach gebauter Gliederfüßer, die bisher als dem postulierten letzten gemeinsamen Vorfahren aller Gliederfüßer sehr nahestehend eingestuft wurden, ist evolutionstheoretisch unerwartet. Ein Stammbaum der Gliederfüßer, der sich am Merkmal „Komplexität des Nervensystems“ orientiert, lässt sich nicht in Einklang bringen mit dem Stammbaum, der vom Merkmal „Komplexität der äußeren Erscheinung“ abgeleitet wird. Das hat zur Konsequenz, dass entweder heute neurologisch einfacher strukturierte Gliederfüßer, wie die oben genannten Kiemenfußkrebse, aber auch Spinnen und Skorpione, die zuvor evolvierte Komplexität im Laufe der Zeit wieder eingebüßt haben müssten. Andernfalls müsste, wenn man die einfach strukturierten Kiemenfußkrebse weiterhin als „primitiv“ und Modell für den letzten gemeinsamen Vorfahren der Gliederfüßer einstufen will, das dreigliedrige Gehirn heutiger höherer Krebse und Insekten als konvergent zu dem von Fuxianhuia angesehen werden, das heißt, es muss sich später noch einmal unabhängig von Fuxianhuia entwickelt haben.
Beide Szenarien sind wenig plausibel. Vielmehr zeigen sie, zu welch problematischen Hypothesen der hier diskutierte objektive Datenbefund zwingt, wenn man eine Evolution von nichts zu etwas, von einfach zu komplex, wie sie gemäß dem naturalistischen Weltbild als gegeben gesetzt ist, bei Berücksichtigung aller Merkmale aufrechterhalten will.
Interessanterweise ist es ein Spezialist auf dem Gebiet der kambrischen (Burgess Schiefer) Fossilien, der sich angesichts der Merkmalverteilung in der belebten Natur genötigt sieht, diese Verkomplizierung der Hypothesen (in diesem Fall die Annahme vielfacher konvergenter Evolution) zum System zu erheben (Braun 2012a).
Die mehrfach zitierte2 Pressemitteilung der Universität von Arizona (Adresse eines der Hauptautoren) ist bezogen auf die beschriebenen Fossilien irreführend, sie spricht von Fuxianhuia als einem „missing link“, während in der gleichen Mitteilung konstatiert wird: „Niemand hatte erwartet, ein so hochentwickeltes Gehirn bei einem so frühen Lebewesen zu finden.“3 Es handelt sich hier sicher nicht um ein lang gesuchtes Bindeglied bzw. ein fehlendes definiertes Puzzleteil. Die Originalarbeit enthält darauf keinerlei Hinweis, sondern schildert das genaue Gegenteil, einen völlig unerwarteten Befund, ein Mosaik einfacher und komplexer Merkmale, wie er gemäß der bisher angenommenen Evolutionsgeschichte der Gliederfüßer nicht erwartet worden ist.
In populärwissenschaftlichen Veröffentlichungen wird oft der Eindruck erweckt, Evolution und Höherentwicklung seien die unwiderlegbare, naheliegende und zwingende Schlussfolgerung aus dem Fossilbefund und aus vergleichenden Untersuchungen an heutigen Organismen. Auf den ersten Blick scheint der allgemeine Fossilbefund durch äußerlich einfachere Fossilien in älteren Schichten tatsächlich eine Entwicklung von einfachen zu „höheren“ Organismen nahezulegen. Dagegen zeigt sich wie im hier beschriebenen Fall bei neuen Fossilfunden regelmäßig an vielen Details, dass selbst äußerlich einfach erscheinende Lebewesen hochkomplex sind und dies bereits zu Beginn ihrer fossilen Überlieferung. Ein weiteres sehr aktuelles Beispiel findet sich bei Zhang (2013), wonach Kelchtiere (Entoprocta) ebenfalls schon im frühen Kambrium nachgewiesen wurden (520 Millionen Jahre, wie Fuxianhuia), wobei diese Fossilien sogar auf einen komplexeren Körperbau hinweisen, als ihn die heutigen Vertreter des Stammes haben. Die Evolution hin zu diesen komplexen Strukturen muss noch früher in der Erdgeschichte postuliert werden, ohne dass dazu bisher fossile Hinweise existieren. Sie wird im Evolutionsmodell vorausgesetzt, bisher ist sie nicht durch Daten gestützt.
Fuxianhuia jedenfalls zeigt, dass das dreigliedrige Gehirn der heutigen höheren Krebse und Insekten schon seit mehr als einer halben Milliarde radiometrischen Jahren existiert und sich zumindest morphologisch nicht erkennbar geändert hat. Man könnte es damit als ein weiteres Beispiel für sogenannte „lebende Fossilien“ anführen, analog zu Quastenflosser und Pfeilschwanzkrebs.
Beide Befunde sind sperrig gegenüber einem Modell allmählicher Evolution im Sinne einer Höherentwicklung von einfach zu komplex. Wie auch schon bei ähnlichen Beobachtungen (Braun 2012b) können sie einfacher schöpfungstheoretisch, im Sinne eines der geschaffenen Natur zugrunde liegenden „Baukastensystems“ gedeutet werden, dessen Komponenten keine größeren Abwandlungen über die Zeit mehr erfahren.
Literatur
Ma X, Hou X, Edgecombe GD & Strausfeld NJ (2012) Complex brain and optic lobes in an early Cambrian arthropod. Nature 490, 258-261, doi:10.1038/nature11495.
Braun HB (2012a) Warten auf einen neuen Einstein. Stud. Int. J. 19, 12-18.
Braun HB (2012b) Kryptische Krebse: Moderne Crustaceen-Fossilien aus dem Kambrium. Stud. Int. J. 19, 93-94. (vgl. Kryptische Krebse: Moderne Crustaceen-Fossilien aus dem Kambrium, https://www.genesisnet.info/index.php?News=186).
Budd GE (2012) Cambrian nervous wrecks. Nature 490, 180-181.
Paterson RJ, García-Bellido DC, Lee MSY, Brock GA, Jago JB & Edgecombe GD (2011) Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes. Nature 480, 237-240.
Zhang Z, Holmer LE et al. (2013) A sclerite-bearing stem group entoproct from the early Cambrian and its implications. Scientific Reports 3, doi:10.1038/srep01066.
Anmerkungen
1 Das Kambrium besteht aus geologischen Schichten, die konventionell auf etwa eine halbe Milliarde Jahre datiert werden. Bekannt ist dieses geologische System vor allem durch die sogenannte „Kambrische Explosion“, das unvermittelte Auftreten von Fossilien der meisten bekannten Tierstämme ohne klar zuordenbare Vorläufer im Präkambrium, den unmittelbar darunterliegenden Schichten. (vgl. Kambrische Explosion, https://www.genesisnet.info/index.php?Artikel=42842&Sprache=de&l=1)
2 http://www.welt.de/wissenschaft/umwelt/article109746112/Gliederfuesser-der-Urzeit-besass-modernes-Gehirn.html; http://www.wissenschaft.de/wissenschaft/hintergrund/316281.html
3 http://uanews.org/story/cambrian-fossil-pushes-back-evolution-complex-brains: “No one expected such an advanced brain would have evolved so early in the history of multicellular animals,” said Strausfeld, a Regents Professor in the UA department of neuroscience.
Autor dieser News: Hans-Bertram Braun
© 2013, http://www.genesisnet.info/schoepfung_evolution/n192.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
03.08.12 Kryptische Krebse: Moderne Crustaceen-Fossilien aus dem Kambrium
Einmal mehr zeigen sehr alt eingestufte Fossilfunde, in diesem Fall von Krebsartigen aus dem Kambrium, große Ähnlichkeit zu heute lebenden Organismen. Fast eine halbe Milliarde Jahre lang und seit ihrem ersten Auftreten haben sich damit einige der hier beschriebenen Krebsartigen nicht sichtbar weiterentwickelt.
Crustacea (Krebsartige) sind die dominante Form der Gliederfüßer im heutigen Meer, während die Insekten das Land dominieren. Krebsartige sind bekannt für ihre große Diversität, Verschiedenartigkeit, Komplexität und ökologische Breite. Sie reichen vom kleinen Wasserfloh bis zu riesigen Tiefseekrabben, landlebend von der Kellerassel bis zum Palmendieb. Die frühe Geschichte der Krebsartigen ist von Lücken im Fossilbericht geprägt. Ihr Fossilnachweis ist vor allem auf Formen beschränkt, die selbst mineralisierte Gehäuse bilden, im Gegensatz zu normalerweise schlecht fossilisierenden Chitinpanzern. Krebsartige wurden bisher vor allem in Schichten beschrieben, die jünger als das Kambrium sind. Das Kambrium besteht aus geologischen Schichten, die konventionell auf etwa eine halbe Milliarde Jahre datiert werden. Bekannt ist dieses geologische System vor allem durch die sogenannte „Kambrische Explosion“, das unvermittelte Auftreten von Fossilien der meisten bekannten Tierstämme ohne klar zuordenbare Vorläufer im Präkambrium, also in den unmittelbar darunterliegenden Schichten. In kambrischen Schichten selbst wurden bisher nur sehr kleine (<2mm) „Orsten-Type“ Formen der Krebsartigen entdeckt (sehr gut dreidimensional erhaltene Fossilien; siehe http://www.core-orsten-research.de/01%20intro%201.html), die allerdings keine große Vielfalt bei der Differenzierung der Körperanhänge zeigen, wie sie sonst bei Crustaceen zu finden ist.
Neue Funde geben nun weitere Aufschlüsse über frühe Krebstiere (Harvey et al. 2012). Es handelt sich dabei nicht um Fossilien kompletter Tiere, sondern um sehr kleine Körperanhänge, die, obwohl nicht im Verbund gefunden, unzweifelhaft von Krebstieren stammen. Die Funde dieser Small Carbonaceous Fossils (SCFs) wurden im Nordwesten Kanadas in Flachmeer-Ablagerungen aus dem mittleren bis späten Kambrium gemacht. Sie wurden aus Bohrkernen von Ölbohrungen herauspräpariert. Aus geringen Mengen Bohrkernmaterial konnten mehrere tausend Körperanhänge ausgewaschen werden. Die verkalkten Fossilien (man schätzt die Körpergröße der ehemaligen Besitzer auf ca. 10-15 mm Gesamtgröße) sind zwar flachgedrückt und klein, aber dennoch mit sehr vielen Details erhalten. Es handelt sich um Mundwerkzeuge (mindestens vier unterschiedliche Typen von Mandibeln), um sehr gut erkennbare komplexe Filteranhänge und andere Extremitäten. Sowohl durch ihre allgemeine Form als auch anhand detaillierter Ornamentierung konnten sie verschiedenen Typen von Krebsartigen zugeordnet werden, z. B. Branchiopoden (Blattfuß- oder „Urzeitkrebse“), Copepoden (Ruderfußkrebse) und Ostracoden (Muschelkrebse). Durch Vergleiche kann man darauf schließen, dass die relativ größeren Typen sich von pflanzlicher Nahrung ernährten, während die kleineren Formen eher Allesfresser waren. Trotz des angenommenen hohen Alters und trotz der Tatsache, dass es sich hier z. T. um erste fossile Dokumentationen der Gruppen handelt, zeigen sie teils überraschende Ähnlichkeit zu heute lebenden Formen. Selbst ausgeprägte Rechts- oder Links„händig“keit, die noch heute bei Krebsen beobachtet wird (z. B. besonders deutlich bei der Winkerkrabbe) findet man bei den hier beschriebenen Fossilien. Damit lässt sich dieses Merkmal unverändert über einen Zeitraum von einer halben Milliarde Jahren nachweisen. Nach Ansicht der Autoren verleitet das „moderne“ Aussehen der SCF dazu, ihre Träger als abgeleitete (d. h. höher evolvierte Formen) einzustufen, obwohl sie wegen ihrer Fundlage theoretisch eigentlich eher ursprünglich sein sollten.
Die detaillierten Ähnlichkeiten zu heute lebenden Formen zeugen von einem frühen Ursprung und anschließender im Wesentlichen unveränderter Überdauerung mehrerer komplexer Futterbeschaffungs-Anpassungen vom Kambrium bis heute, einschließlich der oben erwähnten mandibularen Asymmetrie. Die Fossilien zeigen aber auch deutliche Änderungen in der Ökologie bezüglich Körpergröße und ökologischer Verteilung. Blattfußkrebse leben heute vor allem im Süßwasser, und Ruderfußkrebse sind heute eher kleiner als die hier fossil beschriebenen. Während diese Änderungen potentiell durch mikroevolutive Anpassungen erklärbar sind, bleibt es weiter rätselhaft, warum sich nicht nur in Ausnahmefällen, sondern durchaus häufig wie auch hier im Falle der kambrischen Krebse Organismen vom ersten fossilen Auftreten an zumindest makroskopisch über hunderte Millionen Jahre nicht nennenswert verändert haben.
Die Autoren stellen fest, dass die früheste Radiation von Krebsartigen, die zu den oben beschriebenen verschiedensten Formen geführt haben muss, „im Fossilbericht kryptisch“ sei. Mit anderen Worten, sie existiert im Fossilbericht nicht, es gibt keine Hinweise, wie sich die verschiedenen Untergruppen der Crustaceen herausgebildet haben. Sie erscheinen plötzlich, eine Teil-Explosion der großen kambrischen Explosion, die sie ins Dasein katapultiert, und sie mehr oder weniger unverändert bis heute erfolgreich sein lässt.
Literatur
Harvey THP, Vélez MI & Butterfield NJ (2012) Exceptionally preserved crustaceans from western Canada reveal a cryptic Cambrian radiation. PNAS 109, 1589-1594. Published online before print January 17, 2012, doi: 10.1073/pnas.1115244109.
Autor dieser News: Hans-Bertram Braun
© 2012, http://www.genesisnet.info/schoepfung_evolution/n186.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
06.07.12 Eine Lücke weniger? Vierbeiner aus der Romer-Lücke
Im unteren Teil des Karbons waren bisher fast keine Wirbeltierfossilien bekannt. Nach dem berühmten Paläontologen Alfred S. Romer wird diese Fossillücke als Römerlücke bezeichnet. Nun wurden einige Fossilien aus dieser Lücke beschrieben. Die neuen Funde dehnen die Zeiträume, in denen einige Gattungen existierten, aus, schließen aber keine morphologischen* Lücken.
*morphologisch (= die Gestalt bzw. Baupläne betreffend)
Der Schritt vom Wasser ans Land gehört zu den populärsten Übergängen in der hypothetischen Evolution der Wirbeltiere (vgl. Entstehung der Vierbeiner, https://www.genesisnet.info/index.php?Artikel=42843&Sprache=de&l=1. Wie wurden aus Fischen landlebende Vierbeinern (Tetrapoden)? Die ältesten Tetrapodenfossilien sind aus dem Oberdevon bekannt, es handelt sich um relativ große Formen (ca. 1 m lang und noch größer) wie das achtfingrige wasserlebende Acanthostega oder das berühmte „erste Amphib“ Ichthyostega, das auch wasserangepasst war. Tetrapodenspuren, die eindeutig auf dem Land hinterlassen wurden, sind bereits aus dem untersten Mitteldevon bekannt (Niedzwiedzki et al. 2010; vgl. Frühe fossile Fährten rangieren Übergangsformen aus, https://www.genesisnet.info/index.php?News=141). Am Ende des Oberdevons gab es ein größeres Massensterben, dem auch viele Wirbeltiere zum Opfer fielen. Die dem Oberdevon folgende Fossilüberlieferung ist zunächst äußerst dürftig. Die Schichtfolgen des unteren Unterkarbons lieferten bisher nur wenige Fossilfunde. Bis auf wenige Ausnahmen werden erst nach einer zeitlichen Lücke von etwa 20 Millionen Jahren wieder Tetrapodenfossilien gefunden, dann aber in einer großen Vielfalt von Formen, die komplett an das Leben an Land angepasst waren. Benton (2007, 101) spricht von „völlig anders zusammengesetzten Wirbeltierfaunen des Oberdevons“ gegenüber der ältesten bekannten unterkarbonischen terrestrischen (landlebenden) Wirbeltiergemeinschaft. Sogar Stammgruppen von Amnioten (Tiere mit einer Amnionhöhle, hier sind Reptilien gemeint) „mit einem im Wesentlichen modernen Aspekt“ (Smithson et al. 2012, 4532) und hochspezialisierte sekundär wasserlebende Formen sind darunter.
Die Überlieferungslücke ist so markant, dass sie eigens eine besondere Benennung erhielt: „Romer-Lücke“ – nach dem berühmten Wirbeltierpaläontologen Alfred S. Romer. Für den Großteil der Fossilgruppen beträgt die Lücke sogar 30 Millionen Jahre: „Im Gegensatz zu unseren detaillierten Kenntnissen über Acanthostega und Ichthyostega ist der Fossilbericht der ersten 30 Millionen Jahre des Karbons karg und verwirrend“ (Carroll 2009, 61).
Bisher war unklar, ob es sich um eine reine Überlieferungslücke handelt – dass also in der betreffenden Zeit viele Vierbeiner lebten, jedoch nicht fossil überliefert wurden – oder ob es in der betreffenden Zeit in Wirklichkeit kaum Vierbeiner gab und aus diesem Grunde fast keine Fossilien gefunden wurden. Als Gründe für letztere Deutung des Fehlens von Fossilien gab es nur Mutmaßungen, z. B. ungünstige Atmosphärenbedingungen. Nun berichten Smithson et al. (2012) von einer größeren Anzahl von Funden von Tetrapoden und Gliederfüßern an vier Lokalitäten im Süden Schottlands. Sie werden geologisch ins Tournaisium und damit zeitlich in die Romer-Lücke gestellt. Unter den Fossilien waren sowohl wasser- als auch landlebende Formen, und unter den Tetrapoden-Fossilien kleine und große Formen. Zu den Wirbeltierfunden zählen neben den Vierbeinern auch Lungefische. Die Funde stammten aus mehreren verschiedenen Horizonten. Damit ist für die Forscher klar, dass die Romer-Lücke ein Artefakt ist und auf einen bisher mangelnden Erfolg der Fossiliensucher zurückzuführen ist.
Was wurde gefunden? Smithson et al. (2012) heben unter den Funden in Burnmouth eine kleine, fünffingrige Extremität hervor, die bekannten unterkarbonischen Gattungen, aber auch einer oberkarbonischen Gattung ähnele. Ein Unterkieferbruchstück kann der Gattung Crassigyrinus zugeordnet werden, die bisher erst ab dem späten Viséum bekannt war. Die Fossilien von Burnmouth gleichen damit eher späteren karbonischen als früheren devonischen Formen, so die Autoren. Am Fundort „Willie’s Hole“ wurden zwei neue Formen gefunden, die in einigen Merkmalen Ähnlichkeiten teils mit Pederpes, teils mit Crassigyrinus aufweisen. Pederpes war bisher eine der wenigen gut erhaltenen Gattungen aus der Romer-Lücke (Clack & Finney 2005). Die Autoren fassen zusammen, dass die Existenz einiger Linien weiter in die Vergangenheit gezogen werden müsse, und zwar bis zu 20 Millionen Jahre; das gilt insbesondere für die Fünffingrigkeit. Man muss daher evolutionstheoretisch annehmen, dass viele Tetrapodenlinien sehr viel früher entstanden sind als bisher angenommen. Die Tetrapoden hätten sich nach dem Massenaussterben am Ende des Devons deutlich schneller erholt als bisher gedacht.
Bewertung. Gleichgültig, ob es sich um eine Überlieferungslücke handelt oder ob tatsächlich nur wenige Vierbeiner existierten: der Ursprung der Vielfalt der ab dem Viséum (mittleres Unterkarbon) überlieferten Formen bleibt so rätselhaft wie zuvor. Die neuen Funde dehnen die Zeiträume, in denen einige Gattungen existierten, aus, schließen aber keine morphologischen (die Baupläne betreffenden) Lücken. Im zeitlichen Sinne wird die Romer-Lücke daher mit einigen Formen gefüllt. Die morphologischen Lücken zwischen den wassergebundenen, großen oberdevonischen Tetrapoden und den kleineren, vollständig landlebenden Formen des Karbons bleiben aber im Wesentlichen unverändert. Die Vielfalt der Formen als dem Viséum ist unverändert sehr viel größer als das Spektrum der älteren karbonischen Formen aus dem Tournaisium. Formulierungen der Tagespresse über „spektakuläre Entdeckungen“, die Lücken schließen, sind daher missverständlich.
Literatur
Benton MJ (2007) Paläontologie der Wirbeltiere. München. Carroll R (2009) The rise of amphibians. 365 million years of evolution. Baltimore.
Clack JA & Finney SM (2005) Pederpes finneyae, an articulated tetrapod from the tournaisian of Western Scotland. J. Syst. Pal. 2, 311-346.
Niedzwiedzki G, Szrek P, Narkiewicz K, Narkiewicz M & Ahlberg PE (2010) Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43-48.
Smithson TR, Wood SP, Marshall JEA & Clack JA (2012) Earliest Carboniferous tetrapod and arthropod faunas from Scotland populate Romer’s Gap. Proc. Natl. Acad. Sci. 109, 4532-4537.
Autor dieser News: Reinhard Junker
© 2012, http://www.genesisnet.info/schoepfung_evolution/n185.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.07.11 „Moderne Optik“ im frühen Kambrium
Die Evolution der Augen ist ein vieldiskutiertes Thema in der Evolutionsbiologie und unter Evolutionskritikern (vgl. Ullrich et al. 2005). Dabei stehen Vergleiche verschiedener Augentypen heutiger Arten und theoretische Überlegungen über Neuerwerb einzelner Bestandteile komplexer Augen z. B. durch sogenannte Kooptionen* im Vordergrund. Welchen Beitrag aber liefern Fossilfunde? Hier ist weitgehend Fehlanzeige zu verzeichnen. „Der Fossilbericht war bis jetzt unzureichend, um Einblicke in die frühe Evolution der Augen während der anfänglichen Radiation vieler Tiergruppen zu ermöglichen, die als kambrische Explosion bekannt ist“ (Lee et al. 2001, 631). Außer von Trilobiten-Augen sei kaum etwas über die Details des optischen Designs der kambrischen Tierwelt, aus der die ältesten fossil erhaltenen Augen stammen, bekannt, trotz z. T. sehr guter fossiler Erhaltung. Nun berichten Lee et al. (2011) von sehr gut erhaltenen fossilen Augen aus dem Frühen Kambrium des Emu Bay-Schiefers in Australien (auf 515 Millionen Jahre datiert), die sie als „sehr hochentwickelte“ (highly advanced) Komplexaugen (Facettenaugen) charakterisieren. Sie sind in fein geschichtetem Tonstein eingebettet.
*Kooptionen (= Übernahme, Wiederverwendung in einem neuen Zusammenhang)
Es handelt sich um mehrere isolierte Augen, die aus jeweils über 3.000 ommatidischen Linsen in dichter und in ausgeprägt sechseckiger Anordnung bestehen, was die kompakteste und effizienteste Anordnung darstellt. (Als Ommatidien werden die Einzelelemente von zusammengesetzten Augen bezeichnet, wie sie in der heutigen Tierwelt bei Gliederfüßern vorkommen.) Der Durchmesser der Augen beträgt 7-9 mm. Die Forscher ordnen die Augen aufgrund ihrer Größe einem Gliederfüßer zu, der vermutlich räuberisch lebte und unter schwachen Lichtverhältnissen sehen konnte. Eine genauere Zuordnung ist nicht möglich. Die Augen sind komplexer als fossile Augen zeitgleich existierender Trilobiten und so hochentwickelt wie die Augen vieler heute lebender Formen. Die Augen besitzen im Zentrum große ommatidische Linsen, die eine besonders lichtempfindliche „helle Zone“ („bright zone“ oder „acute zone“) bilden, während die Linsen in den Randbereichen kleiner sind. Diese Spezialisierungen seien für viele moderne Taxa charakteristisch. Die Wissenschaftler schließen aus der Anordnung der verschiedenen Linsengrößen, dass die Augen auch im Lebenszustand flach waren. Die extrem reguläre Anordnung der Linsen übertrifft sogar die Anordnung bei manchen heute lebenden Formen wie dem Pfeilschwanz Limulus. Damit besaßen einige der ältesten Gliederfüßer optische Systeme, die denen heutiger Formen glichen.
Literatur
Lee MSY, Jago JB, García-Bellido DC, Edgecombe GD, Gehling JG & Paterson JR (2011) Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia. Nature 474, 631-634.
Ullrich H, Winkler N & Junker R (2005) Zankapfel Auge. Ein Paradebeispiel für „Intelligent Design“ in der Kritik. Stud. Int. J. 13, 3-14; online: http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html.
Autor dieser News: Reinhard Junker
© 2011, http://www.genesisnet.info/schoepfung_evolution/n173.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.05.11 Eintagsfliege hinterlässt fossile Spuren
Eintagsfliegen treten vergleichsweise früh im Fossilbefund auf. Sie werden als ursprüngliche Fluginsekten angesehen und stellen heute eine Ordnung der Insekten mit weltweit mehr als 2500 bekannten Arten dar (Europa: ca. 300). Hier wird ein fossiler Abdruck eines Insektes vorgestellt, den die Autoren mit guten Gründen den Eintagsfliegen zugeordnet haben. Diese Insektengruppe ist von Beginn ihres fossilen Nachweises als solche erkennbar.
Fossilien, ausgegrabene Reste und Spuren vergangener Lebewesen, erwecken immer wieder öffentliches Interesse oder sind Objekt der Sammelleidenschaft von Menschen. Die Wahrscheinlichkeit, mit der ein Lebewesen Spuren hinterlässt, die in irgendeiner Art und Weise erhalten bleiben, hängt von unüberschaubar vielen Faktoren ab. Für Lebewesen mit beständigen, mineralisierten Körperbestandteilen würde man eher mit fossiler Erhaltung rechnen als beispielsweise bei Weichtieren. Lebewesen können aber auch indirekte fossile Hinweise hinterlassen, wie z. B. Trittsiegel (von Dinosauriern, aber auch von Menschen) oder Fraßspuren, Koprolithen („Kotsteine“). und vieles andere mehr. Unter Spurenfossilien (Seilacher 2007) versteht man eine ganz besondere Klasse von Fossilien, die indirekte Hinweise auf Organismen oder deren Aktivitäten überliefern, die oft nur schwer oder gar nicht einem spezifischen Urheber zugeordnet werden können.
Aus der Wamsutta-Formation (Oberes Karbon) aus dem Südosten von Massachusetts, USA beschreiben Knecht et al. (2011) in einem roten sehr feinen Sandstein den Abdruck, den eine Eintagsfliege durch die Landung in weichem Schlamm hinterlassen hat. Sehr gut abgebildet sind die Bauchseite des Insektenkörpers und Eindrücke der Beine bzw. deren verschiedene Bodenkontakte (und möglicherweise darüber hinaus noch Spuren von Flügelschlägen). Es ist also nicht das Insekt selbst fossil erhalten, sondern nur der Eindruck, den das Tier bei der Landung hinterlassen hat, was sehr spezielle Umstände erforderte. Der fossile Abdruck liegt nun in einem Stück (ca. 10 x 6 x 1 cm) und zwar sowohl die Platte mit den Vertiefungen (Epirelief) als auch die zugehörige überlagernde Platte mit deren Ausfüllungen (Hyporelief). Die Platten enthalten auch zwei leicht asymmetrische Rippelmarken.
Die im Sandstein abgebildete Körperstruktur der Eintagsfliege hat eine Länge von 3,6 cm und enthält schwache Andeutungen des Kopfbereichs, an den sich durch einen kurzen (1,5 mm) Übergangsbereich (Prothorax) die Brust (Thorax) 0,5 mm tief ins Sediment eingedrückt hat. Die drei Beinpaare sind in Thoraxnähe ebenfalls abgebildet. Im segmentierten Hinterleib (Abdomen, Länge: 20,6 mm) ist durch die Abbildung der (mindestens 10 unterscheidbaren) Segmente angedeutet, dass der am tiefsten liegende Körperbereich bei Segment 8 liegt und das Ende angehoben wurde. Am Ende des Abdomens sind möglicherweise Körperanhänge (Cerci) abgebildet.
Aufgrund der abgebildeten Merkmale identifizieren die Autoren den Verursacher dieser eingedrückten Spuren als Vertreter der Eintagsfliegen (Ephemeropterida). Weiter stellen sie fest, dass es sich bei der Eintagsfliege um ein erwachsenes, d. h. geschlechtsreifes Exemplar oder eines im letzten Lavenstadium handeln sollte. Aufgrund der fehlenden Information über die Flügelstruktur kann eine genauere Zuordnung nicht vorgenommen werden, da die paläozoischen Ephemeropterida vor allem nach dem Muster der Flügeladern klassifiziert werden. Knecht et al. spekulieren darüber, dass Syntonopteridae und Protereismatidae aufgrund biogeographischer Gegebenheiten die wahrscheinlichsten Kandidaten sind.
Im frühen Karbon ist fossil eine Vielfalt von fliegenden Insekten belegt (typischerweise durch isolierte Flügel). Die Befunde eröffnen allerdings keine Einsicht in die Lebenszusammenhänge dieser Lebewesen.
Bei dem von Knecht et al (2011) vorgestellten fossilen Körperabdruck (full-body impression) handelt es sich um den frühesten eines fliegenden Insekts. Aus dem unter dem Karbon liegenden Perm sind indirekte fossile Hinweise gefunden worden (Rhyniognatha: Mandibeln, Mundwerkzeuge), die aufgrund ihrer Ausprägung fliegenden Insekten zugeordnet werden (Engel & Grimaldi 2004; Grimaldi & Engel 2005).
Es ist interessant, dass der älteste bekannte Körperabdruck eines Insekts genügend spezifische Hinweise beinhaltet, dass der Verursacher erstaunlich genau eingegrenzt werden kann. Hier liegt also ein weiterer Beleg dafür vor, dass Organismen beim ersten fossilen Auftreten einer bestimmten Gruppe zugeordnet werden können, weil sie deren charakteristische Merkmale aufweisen. Fossile Hinweise auf eine evolutionäre Vorgeschichte fehlen bisher.
Literatur
Engel MS & Grimaldi DA (2004) New light shed on the oldest insect. Nature 427, 627–630.
Grimaldi D & Engel MS (2005) Evolution of the Insects. Cambridge.
Knecht RJ, Engel MS & Benner JS (2011) Late Carboniferous paleoichnology reveals the oldest body impression of a flying insect. Proc. Nat. Acad. Sci USA, doi: 10.1073/pnas.1015948108
Seilacher A (2007) Trace Fossil Analysis. Berlin.
Autor dieser News: Harald Binder
© 2011, http://www.genesisnet.info/schoepfung_evolution/n167.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
13.04.11 Erhebliche Unterschiede in der Hirnphysiologie
Biochemische Charakterisierung des menschlichen Gehirns und Vergleich zu anderen Primaten
Die komplexen Leistungen des Gehirns spiegeln sich nicht nur in seiner Struktur sondern auch im Stoffwechsel des neuronalen Netzwerkes wieder. Die bisher umfangreichste vergleichende Studie von Metaboliten in verschiedenen Gehirnbereichen von Primaten zeigt interessante Besonderheiten des menschlichen Gehirnstoffwechsels. Dies wirft Fragen nach ihrer Entstehung auf.
Mit immer leistungsfähigeren Methoden werden ungeheure Mengen an molekularbiologischen Daten produziert. Diese Datenfülle kann trotz riesiger Rechnerkapazitäten und unterschiedlichster Programme bisher nur eingeschränkt genutzt werden. Gegenwärtig haben sich die Hoffnungen, den Menschen auf der Basis seiner stofflichen Beschaffenheit zu charakterisieren und letztlich zu verstehen (noch?) nicht erfüllt. Die wachsenden Fossilbefunde und vergleichende Genomuntersuchungen haben Einblicke in verschiedenste hoch interessante Aspekte des Menschen eröffnet, ohne jedoch die erhoffte Lösung um die Rätsel seines Wesens liefern. Es ist derzeit nicht absehbar wie und ob mit naturwissenschaftlichen Methoden die Frage: „Was macht den Mensch zum Menschen?“ beantwortet werden kann.
Fu et al. (2011) haben nun eine Untersuchung vorgestellt, bei der ein internationales Team unter Leitung von Willmitzer und Khaitovich vom Max Planck Institut für Evolutionäre Anthropologie in Leipzig mehr als 100 Stoffwechselprodukte (Metabolite) im vorderen Bereich der Großhirnrinde (Gehirnlappen, Cortex), dem Präfrontalen Cortex (genauer: Gyrus frontalis superior) und aus dem Kleinhirn des Menschen, Schimpansen und Rhesusaffen analysiert wurden.1 Aus Gewebeproben von je 100 mg wurden die Metabolite extrahiert, chemisch modifiziert und dann gaschromatographisch und massenspektrometrisch untersucht. Für die Untersuchung standen Proben von 49 Menschen (0-98 Jahre), 11 Schimpansen (0-40 Jahre) und 45 Rhesusaffen (Macaca mulatta) (0-28 Jahre) zur Verfügung. Fu et al konzentrierten sich auf 118 Stoffwechselprodukte, wovon 61 chemisch identifiziert wurden und 57 unbekannt sind, d.h. letztere konnten nicht eindeutig Standartsubstanzen zugeordnet werden.
Die möglichen Veränderungen der Konzentrationen der Substanzen nach Eintritt des Todes diskutieren die Autoren detailliert, beziehen das in ihre Überlegungen mit ein und argumentieren entsprechend vorsichtig. Sie versuchen den Einfluss dieser empirischen Unsicherheiten durch Anwendung statistischer Methoden zu verringern.
Nach Angaben von Fu et al. sind 49 % der Unterschiede im Vorkommen der Metabolite auf Unterschiede zwischen den Arten zurückzuführen, 17 % auf das Alter und 9 % auf die Entnahme aus den zwei unterschiedlichen Gehirnregionen. Es ist also möglich, den Unterschied im Metabolismus verschiedener Arten von Lebewesen wie auch verschiedener Alter anhand von Analysen von Stoffwechselprodukten aus beiden Gehirnbereichen darzustellen.
Fu et al. untersuchten dann artspezifische Unterschiede, indem sie auf der Basis der Daten für Schimpansen die Unterschiede der Stoffwechselprodukte zwischen Menschen und Rhesusaffen in spezifisch menschlich, spezifisch für Makaken und unspezifisch zu kategorisieren versuchen. Im Kleinhirn konnten sie 6 für Menschen und 33 für Rhesusaffen spezifische Unterschiede im Stoffwechsel ermitteln. Im präfrontalen Cortex dagegen fanden sie 24 für Menschen spezifische und 20 für Rhesusaffen spezifische Unterschiede im Metabolismus. Das bedeutet, dass im vorderen Bereich der Großhirnrinde – die nach Standard-Lehrbuch-Aussagen evolutionär jünger und beim Menschen im Vergleich zu anderen Primaten besonders ausgeprägt ist – die 4-fache Anzahl an Stoffwechseländerungen im Vergleich zum – evolutionär „älteren“ – seitlichen Teil der Großhirnrinde vorliegt. Weder bei Schimpansen noch bei Rhesusaffen konnten ähnliche Unterschiede zwischen den beiden Gehirnbereichen festgestellt werden.
Legt man eine gemeinsame Abstammung der Primaten zugrunde, dann sprechen diese Befunde für eine erstaunlich intensive und vergleichsweise schnelle Veränderung im Gehirn des Menschen im Vergleich zu den andern Primaten. Die biochemischen Veränderungen sind besonders im evolutionär jung gedachten und beim Menschen besonders ausgeprägten präfrontalen Cortex. Die Daten an sich belegen zunächst aber einfach einmal bemerkenswerte Unterschiede in der Gehirnphysiologie zwischen Menschen Schimpansen und Rhesusaffen. Sie werfen im evolutionstheoretischen Rahmen aber auch neue Fragen nach den zugrundeliegenden Mechanismen für die schnellen Veränderungen auf.
In einer weiteren Versuchsreihe wurden die biosynthetisch erzeugten (exprimierten) Proteine, die als Enzyme am Stoffwechsel beteiligt sind, in Proben aus dem Präfrontalen Cortex untersucht. Fu und Mitarbeiter bestimmten 2747 Proteine bei Menschen, 2343 bei Schimpansen und 2842 bei Rhesusaffen, wobei 1951 in allen Arten vorkommen.
Die Autoren fanden eine signifikante Übereinstimmung zwischen den spezifisch menschlichen Stoffwechselprofilen in der vorderen Großhirnrinde und den entsprechenden exprimierten Enzymen.
Für alle drei untersuchten Primatenarten stellen die Autoren fest, dass sich der Gehirnstoffwechsel im Verlauf der Individualentwicklung mit dem Alter signifikant verändert, für 88 % der Metabolite ändert sich die Konzentration während der Lebensdauer in mindestens einer der untersuchten Arten oder in einer Gehirnregion.
Fu et al. betont besonders die 4-fache Änderung des Stoffwechsels im Frontalbereich der Großhirnrinde im Vergleich zum Kleinhirn.
In der Diskussion ihrer Ergebnisse fokussieren Fu und Mitarbeiter auf Glutaminsäure (Glutamat) als einem für den Menschen spezifischen Metaboliten von besonderer Bedeutung sowohl im Energiestoffwechsel als auch Weiterleitung von Nervenimpulsen. In der menschlichen Großhirnrinde stellt der Stoffwechsel von Glutamat, d.h. dessen Synthese, Freisetzung und das Recycling den bedeutendsten Anteil dar, für ihn werden 60-80 % der Energie, die durch Oxidation von Glucose gewonnen wird, aufgewendet. Glutamat-Dehydrogenase ist ein wichtiges Enzym im Glutamat- und Energiestoffwechsel der Zelle. Fu et al. interpretieren Befunde aus anderen Arbeiten über dieses Enzym als Hinweis auf Genduplikation und Veränderung in der Funktionalität in einem postulierten gemeinsamen Vorfahren von Menschen und Affen, die zu einem für Affen und Menschen spezifischen Gen (GLUD2) geführt haben. Die Untersuchungen der hier vorgestellten Arbeit zeigen, dass weitere Veränderungen des Glutamatstoffwechsels bei Menschen in dessen Präfrontalen Cortex, nicht aber im Kleinhirn stattgefunden haben.
Die Autoren erhoffen sich von weiteren und umfangreicheren Studien dieser Art Einblicke in die gesamten Änderungen des Stoffwechsels, die sich im Verlauf der Evolution des Menschen ereignet haben. Man darf auf weitere Untersuchungen und die daraus resultierenden Erkenntnisse über molekulare Charakterisierungen des Menschen in unterschiedlichen Lebensaltern und im Vergleich zu anderen Lebewesen gespannt sein.
Festzuhalten aber bleibt, dass derzeit nicht zu erkennen ist, wie durch diese Art von Einsichten wesentliche Antworten auf die Frage: „Was macht den Menschen zum Menschen?“ erzielt werden können. Mit den hier vorgestellten Befunden und Erkenntnissen werden deutliche Unterschiede zwischen Mensch und anderen Primaten dokumentiert, die eine Herausforderung für evolutionäre Erklärungen darstellen und mit einer Begriffswahl, wie „Turboevolution“2 verschleiert werden.
Literatur
Fu X, Giavalisco P, Liu X, Catchpole G, Fu N, Ning Z-B, Gou S, Yan Z, Somel M, Pääbo S, Zeng R, Willmitzer L & Khaitovich P (2011) Rapid metabolic evolution in human prefrontal cortex. Proc Nat. Acad. Sci. 108, 6181-6186; doi/10.1073/pnas.1019164108, mit umfangreichem zusätzlichen Datenmaterial unter: www.pnas.org/lookup/suppl/doi:10.1073/pnas.1019164108/-/DCSupplemental.
Anmerkungen
1 Das Autorenteam benutzte Gewebeproben von toten Organismen, die keine relevanten pathologischen Erscheinungen aufwiesen von verschiedenen Gewebebanken auf der ganzen Welt. Für alle Proben von Menschen lagen schriftliche Einverständniserklärungen von deren nächsten Angehörigen vor.
2 Dieser Begriff wurde in populärwissenschaftlichen Darstellungen dieser Untersuchung und in Meldungen der Tagespresse verwendet.
Autor dieser News: Harald Binder
© 2011, http://www.genesisnet.info/schoepfung_evolution/n165.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
17.03.10 Täuschen Verwesungsstadien evolutionäre Abfolgen vor?
Der Erhaltungszustand und der Informationsgehalt von Fossilien hängen stark von ihrem Verwesungsgrad und dem Zeitpunkt ihrer Überdeckung durch Sediment ab. Besonders die nicht-mineralisierten Weichteile wie Muskeln, innere Organe oder Hautstrukturen zersetzen sich schnell, während Skelett und Zähne länger widerstehen und darum auch öfter gefunden werden.
Bisher war allgemeine Meinung, dass der Zerfall der Weichteile verendeter Organismen keiner speziellen Regel folge und damit ohne Konsequenzen für deren systematische Einordnung bei der Rekonstruktion ihrer Phylogenese sei. Untersuchungen von Sansom et al. (2010) zeigten jetzt, dass diese Annahmen falsch sind (vgl. auch den Kommentar von Briggs 2010). Sansom et al. töteten Lanzettfischchen (Branchiostoma lanceolatum, ein relativ einfach gebautes Chordatier) und Larven des Flussneunauges (Lampetra fluviatilis, ein einfaches Wirbeltier) ab, lagerten sie in Meerwasser und beobachteten, in welcher Reihenfolge die Körperteile verwesten. So zeigte sich, dass Teile des Kopfes regelhaft vor denen des Körperstammes verwesen. Beim Flussneunauge im Speziellen war das mehrkammerige Herz nach 11, die Augen nach 64 und Darm und Leber erst nach 130 Tagen verschwunden. Beim Lanzettfischchen ging zuerst der Augenfleck verloren, während wie beim Flussneunauge die Muskelblöcke (Myomere) des Körperstammes am längsten nachweisbar blieben.
Das interessanteste Ergebnis ist, dass die grundlegenden, bauplantypischen Merkmale, die als stammesgeschichtlich alt bewertet werden mit dem Zeitpunkt ihres Verschwindens während des Verwesungsprozesses auffällig korrelieren: Die nach evolutionären Hypothesen zuletzt erworbenen Merkmale (apomorphe Merkmale) verwesen bis auf wenige Ausnahmen regelhaft zuerst, die phylogenetisch alten und von gemeinsamen Vorfahren übernommenen Merkmale (plesiomorphe Merkmale) lösen sich zuletzt auf. Je länger also die Kadaver des Lanzettfischchens oder des Flussneunauges ungestört zerfallen konnten, umso mehr ähnelten sie ihrer gemeinsamen hypothetisch-evolutionären Stammform.
Aus diesen empirischen Befunden ergeben sich, so die Autoren, tiefgreifende Konsequenzen für die Interpretation des Fossilberichtes bezüglich der Herleitung der Wirbeltiere. Die bereits existierende Unsicherheit bei der Interpretation von Fossilien von Chordatieren und verwandten Formen und damit bei der Erstellung evolutionärer Stammbäume wird noch weiter verschärft. Da anzunehmen ist, dass die am Flussneunauge und am Lanzettfischchen beobachtete Phänomene – die evolutionär „jüngeren“ Merkmale verwesen zuerst und die evolutionär „älteren“ zuletzt – auch für die fossil bekannten Formen gelten, sind wahrscheinlich viele Fossilien zu Unrecht als zu „einfach“ beschrieben worden. Das heißt „moderne“ Formen können auch die Quelle für Fossilien sein, die bisher als repräsentativ für primitive Formen galten. Die auf der Basis des morphologischen (= gestaltlichen) Vergleichs von Fossilien rekonstruierten stammesgeschichtlichen Abfolgen von „primitiv“ zu „komplex“ könnten deshalb in bestimmten Fällen einen evolutionären Wandel nur vortäuschen. Die Autoren machen klar, dass bisher keine methodischen Werkzeuge verfügbar sind, um zwischen evolutionär bedingter und einer durch den Zerfall eines toten Körpers hervorgerufenen Bauplaneinfachheit zu unterscheiden.
Die durch diese Zusammenhänge mögliche Fehlbewertung von Lebensformen als evolutionär primitiv bzw. ursprünglich („stem-ward-slippage“, etwa „Stammgruppenschlüpfrigkeit“, wie die Autoren sie nennen), erschwere die Erstellung der evolutionären Stammbäume insbesondere für die basalen („ursprünglichen“) Gruppen der großen Tierstämme erheblich.
Literatur
Briggs DEG (2010) Decay distorts ancestry. Nature 463, 741-743.
Sansom RS, Gabbott SE, Purnell MA (2010) Non-random decay of chordate characters causes bias in fossil interpretation. Nature 463, 797- 800.
Autor dieser News: Wolfgang Lindemann
© 2010, http://www.genesisnet.info/schoepfung_evolution/n144.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
12.01.10 Frühe fossile Fährten rangieren Übergangsformen aus
Fossile Fußabdrücke von Tetrapoden (Vierbeinern) aus dem unteren Mitteldevon Polens sprengen das bisherige Bild von der evolutiven Entstehung der Vierbeiner. Die bisher als Übergangsformen interpretierten tetrapodenähnlichen Fischgattungen Panderichthys und Tiktaalik (Elpistostegalia) verlieren diesen Status und rücken auf einen Seitenzweig, da sie in deutlich jüngeren Schichten gefunden wurden. Auch die bisherigen Vorstellungen über die ökologischen Bedingungen der Entstehung der Vierbeiner sind mit den neuen Funden sehr in Frage gestellt. Für die frühen Tetrapoden wie das bekannte Ichthyostega und andere Gattungen sowie die tetrapodenähnlichen Fische muss nun angenommen werden, dass sie als „ghost taxa“ lange Zeit in geologisch bislang nicht überlieferten Lebensräumen existierten. Die neue Situation erlaubt noch mehr als bisher, die Abfolge der devonischen Fossilien ausschließlich unter ökologischen Gesichtspunkten zu interpretieren. Die Funde reihen sich in die große Zahl von Fossilen ein, die evolutionstheoretisch nicht vorhergesagt worden waren.
Eine Sensation. Überraschend frühe fossile Fußabdrücke von Tetrapoden (Vierbeiner) werden in der ersten Ausgabe 2010 der führenden Wissenschaftszeitschrift „Nature“ beschrieben. Sie stammen aus dem tiefsten Teil des unteren Mitteldevons und werden auf ca. 395 Millionen Jahre datiert. Damit sind sie ca. 18 Millionen Jahre älter als die ältesten bisher bekannten Körperfossilien von Tetrapoden, aber auch ca. 10 Millionen Jahre älter als ihre mutmaßlichen fischartigen Vorfahren der Elpistostegalia wie Panderichthys (s. Abb. 1) und Tiktaalik (Ahlberg & Clack 2006; Daeschler et al. 2006, Shubin et al., vgl. Junker 2006 und den Artikel Tiktaalik – erstklassiges Bindeglied?, https://www.genesisnet.info/index.php?News=63 sowie Abb. 2).

Abb. 1: Panderichthys aus dem unteren Oberdevon. nach Clack JA (2002) Gaining Ground. The Origin and Evolution of Tetrapods. Bloomington, IN.

Abb. 2: Schädel von Panderichthys, Tiktaalik und Acanthostega. Nach Clack JA (2002) Gaining Ground. Bloomington and Indianapolis, sowie Daeschler EB, Shubin NH & Jenkins FA (2006) A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763.
Das polnisch-schwedische Forscherteam um Grzegorz Niedzwiedzki hält aufgrund dieser Funde eine „radikale Neubewertung“ des Zeitpunktes und der ökologischen Umstände des Übergangs von den Fischen zu den Vierbeinern für zwingend geboten (Niedzwiedzki et al. 2010). Janvier & Clément (2010) kommentieren die Funde so, als hätten die Forscher eine „Granate“ in das bisherige Bild der Evolution der Vierbeiner geworfen.
Die Funde. Die fossilen Fußspuren waren zwischen 2002 und 2007 in einem aufgelassenen Steinbruch im Heiligkreuzgebirge bei Zachelmie im Südosten Polens entdeckt worden. Es handelt sich um mehrere Fährtenzüge und um einige einzelne größere Fußabdrücke. Bei einem Fährtenzug sind unterschiedlich große Vorder- und Hinterfüße zu erkennen. Aufgrund der Größe und des Abstands der Fährten-Abdrücke schließen die Forscher auf eine Größe des Tieres von 40-50 cm. Die einzelnen Spuren dagegen sind sehr viel größer und dürften von einem Tier stammen, das ca. 2,5 m lang war. Bei einigen Abdrücken sind die Zehen deutlich erkennbar. Die Spuren können daher nur von einem Tetrapoden stammen, der zu Landgängen in der Lage war. Auch Janvier & Clément (2010) halten diese Deutung in einem Kommentar für überzeugend. Die Spuren passen „beeindruckend“ gut zur Fußanatomie von Ichthyostega; wären sie auf Schichtflächen des Oberdevons gefunden worden, würde man sie ohne Zögern einem Ichthyostega-ähnlichen Tier zuordnen, meinen diese beiden Autoren (zu Ichthyostega siehe Abb. 3 und Artikel Entstehung der Vierbeiner, https://www.genesisnet.info/index.php?Artikel=42843&Sprache=de&l=1). (Bilder gibts bei http://scienceblogs.com/pharyngula/2010/01/tetrapods_are_older_than_we_th.php).
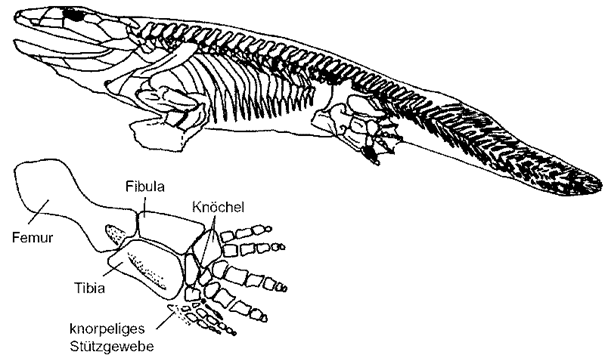
Abb. 3: Ichthyostega aus dem Oberdevon mit 7 Fingern an der Hinterextremität. Quelle: Clack JA (2002) Gaining Ground. The Origin and Evolution of Tetrapods. Bloomington, IN.
Schleifspuren von Körper oder Schwanz wurden nicht beobachtet, was nach Ansicht der Wissenschaftler als Hinweis zu werten ist, dass die Tiere sich im seichten Wasser fortbewegt haben. Die Sedimente, die die Spuren enthalten, weisen aber auch viele Feinschichten (Laminite) mit Trockenrissen und fossilisierten Regentropfeneinschlägen auf. Die Forscher schließen daher auf eine extrem flache marine, vielleicht lagunenartige Umgebung. Es handelt sich nicht nur um die ältesten zweifelsfreien Tetrapodenspuren, sondern auch um die bei weitem reichsten im Devon. Die zeitliche Einordnung wurde anhand von Conodonten-Leitfossilien bestimmt und gilt als sicher, zumal sie auch durch biostratigraphische Einordnung der über- und unterlagernden Sedimente bestätigt wurde.
Evolutionstheoretische Konsequenzen. Die fossilen Fußspuren aus Polen passen in mehrerer Hinsicht nicht zur bisher etablierten Sicht der Evolution der Tetrapoden. Wie schon erwähnt wurden sie dafür in deutlich zu alten Schichten gefunden. Das hat zur Folge, dass sowohl die bislang als ältesten Tetrapoden geltenden Gattungen Ichthyostega, Acanthostega (s. Abb. 4) und andere als auch ihre evolutionstheoretisch-hypothetischen Vorfahren, die Elpistostegalia, in geologisch bislang nicht überlieferten Lebensräumen als sog, „Geisterlinien“ („ghost lineages“) gelebt haben müssen. (Solche Linien müssen aufgrund der mutmaßlichen Stammesgeschichte zu einer bestimmten Zeit existiert haben, sind aber nicht als Fossilien dokumentiert.) Für alle frühen Tetrapoden und für alle Elpistostegalia, die tetrapodenähnlichsten Fische – das sind insgesamt über zehn Gattungen – müssen „ghost ranges“ von ca. 10-20 Millionen Jahren angenommen werden.

Abb. 4: Acanthostega nach Clack JA (2002) Gaining Ground. The Origin and Evolution of Tetrapods. Bloomington, IN.
Bislang war man davon ausgegangen, dass die Elpistostegalia eine kurzlebige „Übergangsstufe“ von Fischen zu Vierbeinern darstellten und dass ihre Ablösung ein evolutionäres Ereignis war. Aufgrund der nun anzunehmenden langen Koexistenz mit Tetrapoden ist diese Deutung nach Auffassung von Niedzwiedzki et al. (2010, 46) nicht mehr haltbar. Die Autoren schreiben: „Das bedeutet, dass der Körperbau der Elpistostegiden kein kurzes Übergangsstadium war, sondern selber ein stabiler angepasster Rang.“1 Diese Situation erinnert sie an die ähnlichen Verhältnisse unter den frühen Vögeln, wo ebenfalls eine sehr lange Koexistenz mit mutmaßlichen Vorfahren zu verzeichnen ist.2
Die Entstehung von Beinen und Fingern muss nun mindestens ins obere Unterdevon (Ems-Stufe) wenn nicht noch früher vorverlegt werden. Aus diesen Schichten sind aber kaum Fossilien von den Fleischflosser-Fischen bekannt, die aufgrund ihrer Anatomie als Vorfahrengruppe am ehesten in Frage kämen, und von diesen eignet sich keines gut als möglicher Vorfahre der Tetrapoden oder Elpistostegalier (Janvier & Clément 2010, 41).
Schließlich müssen auch die ökologischen Vorstellungen über die Begleitumstände der Entstehung der Vierbeiner revidiert werden. Galt es zuletzt als ausgemacht, dass der Übergang zu Vierbeinern im Wasser im Süßwasser-Uferbereich und Brackwasser mit starkem Pflanzenbewuchs stattfand, wo der Besitz von Extremitäten mit Fingern sinnvoll erscheint, plädieren Niedzwiedzki et al. (2010, 47) nun für eine Entstehung im Lagunen-Schlick von Korallenriffen (s. auch Janvier & Clément 2010, 41). Dieser Lebensraum sei für den Übergang ans Land günstig, weil gezeitenbedingt durch die Flut in diesem Lebensraum zweimal täglich Nahrung aus dem Meer herangespült wird.
Ökologische Deutung. Niedzwiedzki et al. (2010, 47) stellen die Frage, warum einerseits die Elpistostegalier (also die mutmaßlichen letzten Tetrapodenvorfahren unter den Fischen) gleichzeitig mit den frühen Tetrapoden gelebt haben, diese andererseits in der Fossilabfolge früher als letztere auftauchen, also in einer Abfolge, die zu einem evolutionären Nacheinander passen würde. Warum passt z. B. die Abfolge „Tiktaalik (u. a.) –> Ichthyostega (u. a.)“ stratigraphisch zu einer vermeintlichen Phylogenie, die aber gar nicht mehr stimmen kann? Die Stratigraphie täuscht sozusagen Evolution vor. Die Autoren sprechen von „falscher stratophylogenetischer Abfolge“! Diese irreführende Abfolge sei „mehr als ein Rätsel“, vielleicht ist die Abwesenheit von Tetrapodenfossilien in den älteren Schichten „einfach Ausdruck ökologischer Präferenz“. „Wenn ihr erstes Erscheinen als Körperfossilien die Zeit widerspiegelt, in der sie erstmals Lebenswelten mit Potential zur Erhaltung besiedelt haben, wie es zu sein scheint, kamen die Elpistostegiden offenbar den Tetrapoden zuvor. Der Grund dafür ist wahrscheinlich ökologisch, kann derzeit aber nicht ermittelt werden“ (S. 47).3 Für eine ökologische Deutung der Fossilien des Devons sprachen schon bisher manche Befunde (es sei dazu auf die zusammenfassende Darstellung von Junker [2005] verwiesen); z. B. die Tatsache, dass die karbonischen Tetrapoden sich nicht leicht an die devonischen anschließen lassen. Benton (2007, 101) spricht von „völlig anders zusammengesetzten Wirbeltierfaunen des Oberdevons“ gegenüber der ältesten bekannten unterkarbonischen terrestrischen Wirbeltiergemeinschaft. Entsprechend wurden viele devonische Tetrapoden auch schon als tote Seitenzweige der Evolution interpretiert; vielleicht gilt das – selbst unter evolutionstheoretischen Vorgaben – für alle.4
Zeitlicher Rahmen. Dass Spurenfossilien zeitlich deutlich vor Körperfossilien fossil erscheinen, ist in einem Kurzzeitszenario und in ökologischer Perspektive viel leichter zu verstehen als in einem Jahrmillionen-Zeitraum. Ein einziges Tier kann fast beliebig viele Spuren hinterlassen, es ist also zu erwarten, dass häufig Spurenfossilien vor Körperfossilien auftauchen (s. u.). Dass aber dazwischen große Zeiträume liegen, ist nicht plausibel und nicht zu erwarten, auch wenn hier nicht zwingend argumentiert werden kann. Eindeutige Schlussfolgerungen verbieten sich schon deshalb, weil der Fossilbefund nur eine Momentaufnahme sein kann – ob er nun zu bestimmten theoretischen Vorstellungen passt oder nicht. Die Fußspuren aus dem polnischen Mitteldevon haben nach Ansicht der Bearbeiter und wissenschaftlichen Kommentatoren erhebliche Konsequenzen für das Verständnis der Evolution der Vierbeiner – neue Funde können jederzeit auch für weitere Überraschungen gut sein.
Weitere Beispiele: Fußspuren lange vor Körperfossilien. Fährten verschiedener Reptilien (z. B. Stammwurzelzähner-Reptilien, „Urkrokodile“, Echsenbecken-Dinosaurier) treten erheblich früher auf als ihre Skelettreste – nach herkömmlicher Datierung 3-10 Millionen Jahre (Stephan 2002, 141). Beispielsweise äußert Haubold (1984, 150) über Fährten mehrerer Reptilienformen der Newark-Gruppe (Trias; östliche USA): „Sie alle eilen wiederum den bis dato durch Skelettreste bekannt gewordenen Formen um Jahrmillionen voraus.“ Das gilt auch für das erste Auftreten von Dinosaurier-Fährten überhaupt. Denn Fährten dreizehiger Dinosaurier erscheinen weltweit erstmals im Benker Sandstein Oberfrankens (unterster Mittel-Keuper, unterste Obertrias), mit ca. 231 Millionen Jahre angegeben.5 Die ältesten Dinosaurier-Skelettreste liegen mit mehreren Arten aus Argentinien (Herrerasaurus, Eoraptor und Pisanosaurus) und ähnlichen Formen in New Mexiko (USA) vor; sie werden in die obere Karn-Stufe (untere Obertrias) datiert (Haubold & Klein 2000, 61) deren Obergrenze mit ca. 220 Millionen Jahren angegeben wird.6 Im Interpretationshorizont der Historischen Geologie liegen dazwischen ca. 10 Millionen Jahre – zwischen den ersten Fährten und den ersten Körperfossilien! Das Problem wird von Schoch (2007, 33) auch für Süddeutschland gesehen; hier ist ebenfalls „das erste Auftreten der Dinosaurier besonders rätselhaft, denn zunächst finden sich nur ihre Fährten“, also ab dem Benker Sandstein (s. o.), und erst im „Unteren Stubensandstein treten dann die ersten Skelettreste auf.“
Übergangsformen kommen und gehen – oder warum historische Evolutionsforschung sich mit Vorhersagen schwer tut. Der Elpistostegalier Tiktaalik wurde vielfach als grandioser Erfolg evolutionärer Theoriebildung und als hervorragende Bestätigung evolutionstheoretischer Vorhersagen gefeiert (so z. B. Shubin 2009, 34). Diese Vorhersage ist mit den neuen Funden stark relativiert. Wenn man überhaupt von „Vorhersagen“ sprechen kann, hätte man dieses Fossil auch im Kontext einer ökologischen Deutung vorhersagen können, denn in Übergangsökologien sind auch am ehesten „Übergangsformen“ zu erwarten, und auch am ehesten dort, wo man bislang schon fündig war – auch dann, wenn man die Fossilabfolge nicht in einen evolutionären Kontext stellt.Das Beispiel der Tetrapodenfährten aus dem Mitteldevon zeigt beispielhaft, dass die historische Evolutionsforschung allenfalls vage Vorhersagen machen kann, was dem Charakter historischer Theorien entspricht. Natürlich widersprechen die von Niedzwiedzki et al. beschriebenen fossilen Spuren nicht einer evolutiven Entstehung, aber weder ihre stratigraphische Position noch ihr Fundgebiet noch ihre Ökologie wurden vorhergesagt. Man hätte eher vorhergesagt, dass man diese Spuren nicht im unteren Mitteldevon und nicht im heutigen Europa findet, wenn man Tiktaalik als stratigraphisch, geographisch und ökologisch vorhergesagtes Bindeglied interpretiert. Genauso wenig wurden vierflügelige Vögel im Oberjura (Hu et al. 2009; Witmer 2009) und Vogelspuren im Grenzbereich Obertrias/Unterjura (Genise et al. 2009) vorgesagt, und diesen Beispielen ließen sich beliebig viele weitere anfügen. Historische Evolutionsforschung deutet Indizien (Fossilien, Merkmale geologischer Schichten usw.) im Nachhinein und passt ihre theoretischen Entwürfe den Indizien immer wieder neu an. Diese Vorgehensweise ist auch in einem schöpfungstheoretischen Ansatz möglich.
Dank: Den Abschnitt „Weitere Beispiele: Fußspuren lange vor Körperfossilien“ sowie einige Hinweise verdanke ich Manfred Stephan.
Anmerkungen
1 „This implies that the elpistostegid morphology was not a brief transitional stage, but a stable adaptive position in its own right.
2 „It is reminiscent of the lengthy coexistence of non-volant but feathered and ‘winged’ theropod dinosaurs with volant stem-group birds during the Mesozoic.“
3 „If their first appearance as body fossils reflects the time when they first colonized environments with preservation potential, as seems likely, the elpistostegids evidently arrived in advance of the tetrapods. The reason was presumably ecological but cannot be determined at present.“
4 Vgl. Carroll (1992, 49): „It has long been clear that none of the early tetrapod groups are ideal ancestors for any of the rest. All exhibit a mosaic of primitive and derived characters that indicate significant periods of independent evolution since the initial divergence of each group.“
5 Der Benker Sandstein ist nach Haubold & Klein (2000, 60) in die höchste Ladin-Stufe einzustufen; der erste Fährten-Horizont liegt noch unter dem Niveau der Bleiglanzbank nahe der Basis des Unteren Gipskeupers (= Basis Grabfeld-/Benk-Formation). Menning & Hendrich (2005, Tab. VII) stufen dieses Niveau mit ca. 231 Millionen Jahren ein.
6 Nach Menning & Hendrich (2005, Tab. VII) liegt die Karn-Obergrenze bei ca. 220 Millionen Jahren.
Literatur
Ahlberg PE & Clack JA (2006) A firm step from water to land. Nature 440, 747-749.
Benton MJ (2007) Paläontologie der Wirbeltiere. München.
Carroll RL (1992) The primary radiation of terrestrial vertebrates. Annu. Rev. Earth Planet. Sci. 20, 45-84.
Daeschler EB, Shubin NH & Jenkins FA (2006) A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763.
Genise JF, Melchor RN, Archangelsky M, Bala LO, Straneck R & de Valais S (2009) Application of neoichnological studies to behavioural and taphonomic interpretation of fossil bird-like tracks from lacustrine settings: The Late Triassic–Early Jurassic? Santo Domingo Formation, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 272, 143-161.
Haubold H (1984) Saurierfährten. Neue Brehm-Bücherei, 479. Wittenberg-Lutherstadt.
Haubold H & Klein H (2000) Die dinosauroiden Fährten Parachirotherium – Atreipus – Grallator aus dem unteren Mittelkeuper (Obere Trias: Ladin, Karn, ?Nor) in Franken. Hallesches Jb. Geowiss. B 22, 59-85.
Hu D, Hou L, Zhang L & Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 460-463.
Janvier P & Clément G (2010) Muddy tetrapod origins. Natur 463, 40-41.
Junker R (2005) Vom Fisch zum Vierbeiner – eine neue Sicht zu einem berühmten Übergang. Teil 3: Tetrapoden des Unterkarbons, unklare Selektionsdrücke und evolutionstheoretische Probleme. Stud. Int. J. 12, 11-18. (Vgl. auch Entstehung der Vierbeiner, https://www.genesisnet.info/index.php?Artikel=42843&Sprache=de&l=1)
Junker R (2006) Tiktaalik – ein erstklassiges Bindeglied? Stud. Int. J. 13, 88-91.
Menning M & Hendrich A (Hg, 2005) Erläuterungen zur Stratigraphischen Tabelle von Deutschland (ESTD) 2002. Newsl. Strat., 41 (1-3).
Niedzwiedzki G, Szrek P, Narkiewicz K, Narkiewicz M & Ahlberg PE (2010) Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43-48.
Schoch R (2007) Die Erfolgsgeschichte der Dinosaurier. In: Schoch R (Hg) Saurier. Ostfildern, 27-34.
Shubin N (2009) Der Fisch in uns. Eine Reise durch die 3,5 Milliarden Jahre alte Geschichte unseres Körpers. Frankfurt/M.
Shubin NH, Daeschler EB & Jenkins FA (2006) The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 440, 764-771.
Stephan M (2002) Der Mensch und die geologische Zeittafel. Holzgerlingen.
Witmer LM (2009) Feathered dinosaurs in a tangle. Nature 461, 601-602.
Autor dieser News: Reinhard Junker
© 2010, http://www.genesisnet.info/schoepfung_evolution/n141.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
15.04.09 Zur Entstehung von Kohle – es kann ganz schnell gehen
Kohle hat in der wirtschaftlichen Entwicklung der Bundesrepublik Deutschland eine bedeutende Rolle gespielt. Beschreibungen von Kohlevorkommen und deren Erklärungen fanden infolgedessen auch entsprechend Eingang in Lehrbücher. Joachim Scheven hat in verschiedenen Publikationen eine Reihe von geologischen Befunden zusammengetragen, die nicht mit gängigen Vorstellungen der Kohleentstehung in langen Zeiträumen in Einklang stehen. Ebenso hat er auch Modelle zur Entstehung von Kohleflözen vorgestellt (Scheven 1982, 1986, 1988). Auch wenn man nicht allen Äußerungen und Schlussfolgerungen des Autors folgt, so bleibt dennoch eine ganze Reihe von Befunden, die etablierte Erklärungen und Modelle in Frage stellen und herausfordern.
Im Folgenden sollen kurz einige Aspekte zur Chemie der Kohleentstehung, dem als Inkohlung bezeichneten Prozess, dargestellt werden.
Friedrich Bergius experimentierte im Rahmen seiner Habilitation (1913) mit Hochdruckverfahren. Ihm wurde 1931 zusammen mit Karl Bosch der Nobelpreis für Chemie verliehen für „ihre Verdienste um die Entdeckung und Entwicklung der chemischen Hochdruckverfahren“. Bergius (1928) fasste seine Untersuchungen zur Inkohlung organischen Materials in einem Artikel zusammen und stellte dabei folgende Punkte heraus:
- Physikalisch-chemische Prozesse der Inkohlung können im Labor unter definierten Bedingungen modellhaft nachgestellt werden.
- Der Reaktionsverlauf und die chemische Beschaffenheit des Endprodukts sind unabhängig von den Ausgangsmaterialien.
- Die Produkte verändern sich nach einer bestimmten Reaktionszeit nicht mehr nachweisbar („Endkohle“).
- Die Analyse der kohleartigen Produkte weist große Ähnlichkeiten zu natürlichen Kohlen auf.
Er hatte mit seinen Mitarbeitern experimentell gezeigt, dass unter entsprechend hohen Drücken und hoher Temperatur organisches Material innerhalb weniger Stunden in kohleartiges Material umgewandelt werden kann. Die von ihm als Endkohle bezeichnete Substanz weist dabei die Qualität von Steinkohle auf.
Unter der Leitung vom Markus Antonietti wurden am Max-Planck-Institut für Kolloid- und Grenzflächenforschung in Potsdam Untersuchungen zur Erzeugung von Nanostrukturen aus Kohlenstoff durchgeführt (Yu et al. 2004, Cui et al. 2006). Dabei handelte es sich im Grunde genommen um eine Wiederholung der Experimente von Bergius vom Beginn des 20. Jahrhunderts. Dessen Ergebnisse wurden dabei bestätigt. In einem Hochdruckautoklaven wurden verschiedene Pflanzenmaterialien innerhalb weniger Stunden in einem Prozess – den die Autoren als „hydrothermale Carbonisierung“ bezeichnen – in kohleartige Substanz umgewandelt. Abhängig von der Reaktionsdauer konnten Antonietti und seine Mitarbeiter torf- bis steinkohleartige Substanzen erzeugen (Titirici et al. 2007a, b).
Damit ist wiederholt in Laborexperimenten, die man durchaus als Modelle zur Entstehung von Kohle betrachten kann, gezeigt worden, dass für die Inkohlung nicht die Zeit, sondern vor allem der Druck-Temperaturbereich entscheidend ist. Bei hohem Druck und entsprechenden Temperaturen kann organisches Material innerhalb von Stunden in Kohle umgewandelt werden. Wie schnell sich die Kohlevorkommen in der Vergangenheit gebildet haben, ist damit nicht gezeigt. Dazu müsste man nach Hinweisen auf die Druck- und Temperaturbedingungen suchen. Man kann aber aufgrund chemischer Erfahrungen feststellen, dass sich Kohle innerhalb sehr kurzer Zeit bilden kann, wenn entsprechende Randbedingungen – Druck und Temperatur – vorliegen.
Literatur
Bergius F (1913) Anwendung hoher Drucke bei chemischen Vorgängen und die Nachbildung des Entstehungsprozesses der Steinkohle (Habilitationsschrift). Knapp, Halle a.d. Saale.
Bergius F (1928) Beiträge zur Theorie der Kohleentstehung. Naturwissenschaften 16, 1-10.
Cui XJ, Antonietti M, Yu SH (2006) Structural effects of iron oxide nanoparticles and iron ions on the hydrothermal carbonization of starch and rice carbohydrates. Small 2, 756-759.
Scheven J (19822) Daten zur Evolutionslehre im Biologieunterricht. Neuhausen-Stuttgart.
Scheven J (1986) Karbonstudien. Neuhausen-Stuttgart.
Scheven J (1988) Mega-Sukzessionen und Klimax im Tertiär. Neuhausen Stuttgart.
Titirici M-M, Thomas A, Antonietti M (2007) Back in the black: hydrothermal carbonization of plant material as an efficient chemical process to treat the CO2 problem? New. J. Chem. 31, 787-789.
Titirici M-M, Thomas A, Yu S-H, Müller J-O, Antonietti M (2007) A direct synthesis of mesoporous carbon with bicontinuous pore morphology from crude plant material by hydrothermal carbonization. Chem. Mater. 19, 4205-4212.
Yu SH, Cui XJ, Li LL, Li K, Yu B, Antonietti M, Colfen H (2004) From starch to metal/carbon hybrid nanostructures: Hydrothermal metal-catalyzed carbonization. Advanced Materials 16, 1636-1640.
Online-Veröffentlichungen der Max-Planck Gesellschaft (Zugriff am 2. 4. 2009): http://www.mpg.de/bilderBerichteDokumente/dokumentation/pressemitteilungen/
2006/pressemitteilung200607121/index.html
Film: Kohle aus Biomasse: http://www.mpg.de/bilderBerichteDokumente/multimedial/filmeWissenschaft/2006/
07/KochKohle/film3.mov
Autor dieser News: Harald Binder
© 2009, http://www.genesisnet.info/schoepfung_evolution/n128.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.04.06 Tiktaalik – ein erstklassiges Bindeglied?
Ein neues Bindeglied schickt sich an, Berühmtheit zu erlangen – Tiktaalik roseae, ein Fisch mit einigen Merkmalen, die typisch für Vierbeiner (Tetrapoden) sind. Anfang April wurde diese neue Fischgattung in zwei ausführlichen Artikeln in Nature beschrieben (Daeschler et al. 2006, Shubin et al. 2006). Überreste mehrerer Exemplare wurden in der kanadischen Arktis auf der Insel Ellesmere gefunden; geologisch gehören die Fundschichten zum untersten Oberdevon (Unterfrasne). Erhalten sind Schädel, Halsbereich, Schultergürtel, Brustflossen, Rippen und ein größerer Teil der Wirbelsäule. Die Tiere waren ca. 1,2-2,7 m groß, hatten einen flachen Körper und einen krokodilähnlichen Kopf und waren mit ihren scharfen Zähnen vermutlich gefährliche Räuber. Erik Ahlberg und Jennifer Clack, zwei ausgewiesene Spezialisten für frühe Tetrapoden, halten es für möglich, dass der neue Fund eine ähnliche Bedeutung erlangen könnte wie der berühmte „Urvogel“ Archaeopteryx (Ahlberg & Clack 2006, 747), und Neil Shubin kreierte anlässlich dieses Fundes den Begriff „fishopod“. Der Gattungsname Tiktaalik entstammt der Sprache der Inuit (Eskimos) und bedeutet „großer Fisch aus seichtem Gewässer“ in Anlehnung an den mutmaßlichen Lebensraum (s. u.); den Artnamen roseae verdankt der Fund einem anonymen Sponsor.
Was macht diesen Fund so interessant? Tiktaalik weist eine ausgeprägte Kombination aus eindeutig fischtypischen Merkmalen und typischen Tetrapodenmerkmalen auf. Fischartig sind die großen Kiemenhöhlen, die ausgeprägten Flossenstrahlen, die Schuppenhaut, der Unterkiefer und der Gaumen. Dagegen sind das verkürzte Schädeldach, der flache Schädel mit nach oben gerichteten Augen, die Ohrregion, die Beweglichkeit der Halsregion, das Fehlen von Kiemendeckeln, der relativ kräftige Brustkorb mit überlappenden Rippen, der flache Körperbau und der Besitz eines Handgelenks Merkmale, die eher zu Tetrapoden hinweisen. Die Forscher schließen aus der Anatomie der Brustflossen, dass sie ähnlich gebeugt und gestreckt werden konnten wie die Gliedmaßen der Landwirbeltiere. Damit war wahrscheinlich ein Kriechen auf dem Grund des Gewässers möglich, vermutlich auch ein Sich-Hochstemmen im Uferbereich und nach Meinung der Wissenschaftler auch kurze Landgänge. Kein Zweifel: Tiktaalik besitzt ein Merkmalsmosaik, das gut in einen Übergangsbereich zwischen Fischen und Vierbeinern passt – hier geht ein Punkt an die Evolutionstheorie.
Was nicht so gut passt. Dass Merkmale von Tiktaalik besonders hervorgehoben werden, die als „Übergangsmerkmale“ gedeutet werden können, ist legitim. Dennoch zeigt die Brustflosse insgesamt doch eher einen fischartigen Charakter (Ahlberg & Clack 2006, 748). Ein Vergleich mit anderen Formen aus dem Übergangsbereich Fische – Vierbeiner macht dies deutlich. Bisher galt Panderichthys (Abb. 1) als tetrapodenähnlichster Fisch; dessen Flossen sind aber deutlich anders gebaut, ihrerseits aber nicht gut als Vorläufer für Tetrapodenextremitäten geeignet. Die Unterschiede zwischen Tiktaalik und gefingerten Gattungen wie Acanthostega sind erheblich (Abb. 2). Ahlberg & Clack (2006, 748) weisen darauf hin, dass der Erwerb von Fingern, von Tiktaalik ausgehend, eine erhebliche Umorganisation („developmental repatterning“) erfordern würde. Der achtfingrige oberdevonische Tetrapode Acanthostega (Abb. 3) war höchstwahrscheinlich ausschließlich wasserlebend; seine Extremitäten waren relativ unbeweglich. Insgesamt eignet sich diese Gattung daher nicht als vermittelnde Form zwischen Tiktaalik und landlebenden Tetrapoden. Man kann es so sagen: Tiktaalik hatte deutlich andere „Übergangsmerkmale“ auf dem Weg zum Landleben als Acanthostega. Der Weg aufs Land konnte nicht über beide Formen zugleich führen, es sei denn, er wurde mindestens zweimal unabhängig durchlaufen, womit dann aber die Vierbeinigkeit nicht mehr als Schlüsselmerkmal (d. h. als Hinweis auf gemeinsame Vorfahren) gelten könnte, sondern konvergent entstanden wäre. Schon länger ist klar, dass auch das berühmte Ichthyostega (Abb. 4), vor noch nicht langer Zeit die Ikone für den Übergang vom Wasser- zum Landleben, deutlich von einer vermittelnden Position entfernt ist (Clack 2002, Ahlberg et al. 2005; zusammenfassend Junker 2004, 2006).

Abb. 1: Panderichthys aus dem unteren Oberdevon. Nach Clack JA (2002) Gaining Ground. The Origin and Evolution of Tetrapods. Bloomington, IN.
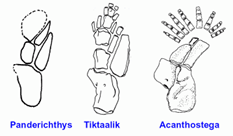
Abb. 2: Vorderextremitäten von Panderichthys, Tiktaalik und Acanthostega. Nach Clack JA (2002) Gaining Ground. Bloomington and Indianapolis, sowie Shubin NH, Daeschler EB & Jenkins FA (2006) The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 440, 764-771.

Abb. 3: Acanthostega. Nach Clack JA (2002) Gaining Ground. The Origin and Evolution of Tetrapods. Bloomington, IN.

Abb. 4: Ichthyostega aus dem Oberdevon mit 7 Fingern an der Hinterextremität
Quelle: Clack JA (2002) Gaining Ground. The Origin and Evolution of Tetrapods. Bloomington, IN.
Nicht nur der Bau der Flossen, sondern auch der Schädelbau passt insgesamt nicht in eine evolutive Reihe von tetrapodenähnlichen Fischen hin zu frühen Tetrapoden. Auch in dieser Hinsicht würde sich Acanthostega auf einer anderen „Schiene“ bewegen, wenn man die relevanten fossilen Gattungen in evolutionäre Linien einfügen wollte (Abb. 5).
Abb. 5: Schädel von Panderichthys, Tiktaalik und Acanthostega.
Nach Clack JA (2002) Gaining Ground. Bloomington and Indianapolis, sowie Daeschler EB, Shubin NH & Jenkins FA (2006) A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763.
Lebensraum. Die Deutung von Tiktaalik als evolutionäre Übergangsform muss mit einigen Unbekannten leben, da die hinteren Teile des Tieres fossil nicht überliefert sind. Über den Bau des Beckens, der Hinterextremitäten und des Schwanzes verraten die Funde leider nichts. Daher kann auch die Lebensweise nicht sicher rekonstruiert werden. (Man sollte in diesem Zusammenhang nicht vergessen, dass lange gemutmaßt wurde, dass die Quastenflosser ihre kräftigen Flossen zur Bewegung auf dem Grund genutzt haben könnten, bis Filmaufnahmen des „lebenden Fossils“ Latimeria diese Vermutung widerlegten.) Wie alle anderen oberdevonischen Vierbeiner und vierbeinerähnlichen Gattungen ist auch Tiktaalik zusammen mit Fischen eingebettet worden (Daeschler et al. 2006, 758). Vermutlich war er ein Beutelauerer im Uferbereich; die lange Schnauze war zum Schnappen nach Beute geeignet, und die beweglichen Extremitäten und der relativ bewegliche Halsbereich erlaubten in diesem Lebensraum eine gute Beweglichkeit. Junker (2004) diskutiert für andere Gattungen, dass es in Uferbereichen eine Vielfalt ökologischer Nischen gegeben haben könnte, „darunter möglicherweise auch (damalige) Lebensräume, die es in dieser Form heute nicht mehr gibt. Dieser Vielfalt könnte eine anatomische Formenfülle mit unterschiedlichen Konstruktions-Mosaiken des Kiemenapparats, der Extremitäten, der Wirbelsäule, des Schwanzes usw. entsprechen.“ In diesem Sinne kann möglicherweise auch Tiktaalik interpretiert werden, als Form, die für einen bestimmten (nicht mehr genau rekonstruierbaren Lebensraum) optimal konstruiert war.
Übergang bisher schlecht belegt? Etwas verwundert stellt man fest, dass Autoren und Kommentatoren Tiktaalik als langgesuchtes Bindeglied zwischen wasser- und landlebenden Wirbeltieren begrüßen. Hieß es nicht schon seit Jahrzehnten, die wesentlichen fossilen Belege für diesen Übergang seien gefunden worden? Nun aber schreiben Ahlberg & Clack, bis vor kurzem sei die morphologische Lücke „frustrierend weit“ geblieben – trotz Panderichthys, Acanthostega und Ichthyostega und anderen overdevonischen Tetrapoden-Gattungen. Ähnlich meinen auch Daeschler et al. (2006, 757), dass die Verwandtschaft zwischen Tetrapoden und Fleischflossern (Sarcopterygier) zwar gut begründet, der Ursprung der wichtigen Tetrapodenmerkmale jedoch in Dunkeln verblieben sei. Und Pennisi (2006) stellt in ihrem Kommentar in Science fest, dass die bislang bekannten Fossilien entweder vornehmlich fischartig oder tetrapodenartig waren statt wirklich intermediär zu sein. Offenbar erlaubt erst die verbesserte Datenlage, den bisherigen Stand des Wissens weniger geschönt darzustellen.
Schlussfolgerungen. Zusammenfassend kann man sagen, dass Tiktaalik das Spektrum von Fischen mit tetrapodenartigen Merkmalen erweitert und in diesem Sinne einerseits einen Baustein für evolutionäre Übergangshypothesen darstellt. Die morphologische Lücke zwischen manchen Formen wird mit dem neuen Fund verkleinert. Andererseits wird nicht die Lücke zwischen den bisher nächststehenden fossilen Gattungen verkleinert (das wäre zwischen Panderichthys und Acanthostega), sondern im Übergangsbereich Wasser – Land eine neue Mosaikform hinzugefügt. Das Merkmalsmosaik von Tiktaalik passt gut zu einem im Uferbereich lauernden Räuber. Ahlberg & Clack (2006, 748) schließen ihren Kommentar mit der Feststellung, dass fast nichts über den Schritt zwischen Tiktaalik und und den frühesten Tetrapoden bekannt sei. Bei diesem Übergang habe die Anatomie die „dramatischsten Änderungen“ erfahren. Außerdem weisen sie auf die markante Lücke zu den karbonischen landlebenden Tetrapoden hin (dazu siehe zusammenfassend Junker 2005).
Es sei noch angemerkt, dass die in Junker (2005) anhand der Fachliteratur zusammengestellten evolutionstheoretischen Probleme mit dem neuen Fund nicht verkleinert werden. Interessierte Leser seien auf diese Publikation oder auf Clack (2002) verwiesen.
Ein ausführlicherer Artikel ist für die Herbstausgabe von Studium Integrale Journal (http://www.wort-und-wissen.de/sij) geplant.
Literatur
Ahlberg PE & Clack JA (2006) A firm step from water to land. Nature 440, 747-749.
Ahlberg PE, Clack JA & Blom H (2005) The axial skeleton of the Devonian tetrapod Ichthyostega. Nature 437, 137-140.
Clack JA (2002) Gaining Ground. The origin and evolution of Tetrapods. Bloomington and Indianapolis.
Daeschler EB, Shubin NH & Jenkins FA (2006) A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763.
Junker R (2004) Vom Fisch zum Vierbeiner – eine neue Sicht zu einem berühmten Übergang. Teil 2: Ichthyostega, Acanthostega und andere Tetrapoden des höheren Oberdevons. Stud. Int. J. 11, 59-66.
Junker R (2005) Vom Fisch zum Vierbeiner – eine neue Sicht zu einem berühmten Übergang. Teil 3: Tetrapoden des Unterkarbons, unklare Selektionsdrücke und evolutionstheoretische Probleme. Stud. Int. J. 12, 11-17. (Siehe auch Entstehung der Vierbeiner)
Junker R (2006) Neue Rekonstruktion bestätigt: Ichthyostega ist kein Bindeglied. Stud. Int. J. 13, 35-36 (im Druck).
Pennisi E (2006) Fossil shows an early fish (almost) out of water. Science 312, 33.
Shubin NH, Daeschler EB & Jenkins FA (2006) The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 440, 764-771.
Autor dieser News: Reinhard Junker
© 2006, http://www.genesisnet.info/schoepfung_evolution/n63.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.01.05 Neuer Artikel über die „kambrische Explosion“ des Lebens
Die Fossilüberlieferung vielzelliger Tiere beginnt mit einer fast schlagartig auftauchenden breiten Vielfalt verschiedenster Baupläne im Unterkambrium (zum Überblick über die geologischen Systeme siehe Abb. 1).

Abb. 1: Vereinfachte geologische Säule. Am Rande befinden sich Altersangaben in Millionen Jahren gemäß radiometrischer Datierungen.
Dieses Phänomen wird heute meist als „kambrische Explosion“ bezeichnet. Es wird auch vom „Urknall der Paläontologie“ gesprochen. Lebewesen aus allen bekannten Tierstämmen, die Hartteile besitzen, sind im Kambrium (meist bereits im Unterkambrium) vertreten. Dazu gehören z. B. Schwämme, Hohltiere, Ringelwürmer, Armfüßer, Gliederfüßer, Weichtiere, Stachelhäuter und auch Chordatiere (darunter als erste Wirbeltiere auch kieferlose Fische). Diese Tierstämme sind zudem von Beginn ihres fossilen Nachweises in der Regel in verschiedene, deutlich abgrenzbare Untergruppen (Klassen) aufgespalten und geographisch meist weit verbreitet. In tieferen Schichten des obersten Präkambriums wurden dagegen nur sehr wenige Vielzeller gefunden, so z. B. einige Hohltiere oder Schwämme (s. u.). Im Artikel „Kambrische Explosion“ (https://www.genesisnet.info/index.php?Artikel=42842&Sprache=de&l=1) wird ein Überblick zur kambrischen Explosion gegeben und es werden Vorschläge zur Erklärung des plötzlichen Auftauchens der Tierwelt kritisch diskutiert. Im Experten-Teil „Kambrische Explosion“ (https://www.genesisnet.info/index.php?Artikel=42842&Sprache=de&l=2) werden außerdem einige diskutierte Vorläufer der kambrischen Tierwelt vorgestellt und Ergebnisse von Untersuchungen mit „molekularen Uhren“ diskutiert. Gut geeignete Vorläuferformen sind nicht bekannt. In einer 2004 erschienenen Monographie stellt der Paläontologe James Valentine, Experte auf diesem Gebiet, fest, dass von keinem einzigen Stamm die Vorläufer bekannt sind, ebenso ist der Weg der Entstehung aller Klassen der Wirbellosen unbekannt.
Autor dieser News: Reinhard Junker
© 2005, http://www.genesisnet.info/schoepfung_evolution/n31.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/