
Paläontologie: Fossilien des Erdmittelalters: Dinosaurier und Vögel im Besonderen
06.12.23 Alte Vogelspuren in Gondwana
Kürzlich wurden 27 Vogel-Fußspuren in Schichten der Unterkreide Australiens gefunden, die unsere Kenntnisse über das Vorkommen von Vögeln auf der Südhalbkugel erheblich erweitern. Bisher waren fossile Überreste von Vögeln in Jura und Kreide fast nur auf der Nordhalbkugel gefunden worden. Dies lässt auf umfangreiche fossil nicht überlieferte Lebensräume schließen, was in einem geologischen Langzeitrahmen schwer verständlich ist.
Unsere Kenntnisse über fossile Vögel in Sedimenten des Jura und der Kreide stammen fast ausschließlich von Funden auf der Nordhalbkugel, dem damaligen Nordkontinent Laurasia. Dagegen sind Funde auf dem Südkontinent Gondwana ausgesprochen selten; nur wenige Federn sowie einige Knochen und Fußspuren waren bisher in Australien und Brasilien gefunden worden. Dabei ist zu bedenken: Nach dem plattentektonischen Konzept lagen alle heutigen Kontinente früher deutlich näher beieinander. Dabei bildeten Nordamerika mit Grönland und dem größten Teil Eurasiens den Kontinent Laurasia, während Südamerika, Afrika, Indien, Australien und die Antarktis zum Südkontinent Gondwana gehörten.
Vor diesem Hintergrund sind kürzlich gemachte Funde von 27 einzelnen Vogel-Fußspuren in der Unterkreide Australiens bemerkenswert (Martin et al. 2023; s. Abb. 1). Die Spuren aus der auf 120–128 Millionen radiometrische Jahre (MrJ) datierten Wonthaggi-Formation zeigen drei dünne lange Zehen, die deutlich voneinander abgewinkelt sind, scharfe Krallen und in wenigen Fällen auch Spuren eines Hallux (1. Zehe). Diese Merkmale sind vogeltypisch; andere Verursacher kommen nach Einschätzung der Forscher nicht in Frage (Martin et al. 2023, 16). Die unterschiedliche Größe der Abdrücke und unterschiedliche Winkel zwischen den Zehen lassen erkennen, dass die Spuren von verschiedenen Arten stammen. Die größten Fußabdrücke sind knapp 12 Zentimeter lang und ca. 14 Zentimeter breit, was ungewöhnlich groß ist für Vögel, die aus Kreideschichten bekannt sind. Es sind zwar keine Fährten unter den Spuren, doch ihre engen Abstände sprechen für eine Geselligkeit der Vögel.

Abb. 1: Versteinerte Fußabdrücke urzeitlicher Vögel (links) und zeichnerische Rekonstruktion. Balken: 5 mm. (© Martin et al./ PLOS ONE, CC-BY 4.0)
Martin et al. (2023) schließen, dass es auf Gondwana im damals polaren Australien vor 120 MrJ eine größere Urvogel-Vielfalt gab als bisher abgenommen. Da in der auf 106 MrJ datierten Eumerella-Formation Australiens bereits zwei Vogel-Fußspuren entdeckt worden waren, gehen die Forscher davon aus, dass Vögel mindestens 15 bis 20 Millionen Jahre lang in der Gegend lebten. Die Fußspuren wurden in verschiedenen, insgesamt ca. neun Meter mächtigen Gesteinsschichten eines Überschwemmungsgebietes gefunden, was darauf hinweisen könnte, dass die Vögel nur saisonal in den Frühlings- oder Sommermonaten nach der Schneeschmelze dort gelebt haben (Martin et al. 2023, 20). Möglicherweise seien Urvögel und vogelähnliche Dinosaurier auf dem einstigen Südkontinent Gondwana häufiger vorgekommen als bislang angenommen. Die Autoren schreiben: „Die morphologische Ähnlichkeit einiger Vogelspuren aus der Wonthaggi-Formation mit denen, die aus Ostasien gemeldet wurden, könnte auch die Ausbreitung der frühkreidezeitlichen flugfähigen Vögel aus Laurasia in andere Teile Gondwanas widerspiegeln, bevor sie nach Australien gelangten.“
Kommentar
Es stellt sich die Frage, warum die fossilen Zeugnisse von Vögeln im Jura und in der Kreide auf der Nordhalbkugel (Laurasia) und der Südhalbkugel (Gondwana) so extrem unterschiedlich häufig sind. „Im Gegensatz zu Gondwana sind Vogelspuren aus der frühen und späten Kreidezeit in den ehemaligen laurasischen Landmassen wie Nordamerika und Asien reichlich und vielfältig vorhanden, auch in den Schichten der frühen Kreidezeit“ (Martin et al. 2023, 19). Man sollte erwarten, dass sich die Vögel aufgrund ihrer Fortbewegungsmöglichkeiten schnell weltweit verbreiten konnten. Man muss folglich davon ausgehen, dass es in großem Umfang und über mehrere zehn MrJ fossil nicht überlieferte Lebensräume gab. Die Autoren halten eine Verzerrung bei der fossilen Erhaltung („preservation bias“) für eine mögliche Erklärung. In einem geologischen Langzeitmodell ist dies jedoch schwer zu verstehen. Sollten die durch die Jura- und Kreideschichten repräsentierten Zeiträume aber in Wirklichkeit viel kürzer sein, wäre das Fehlen einer reichhaltigeren Fossilüberlieferung vielleicht leichter verstehbar. Wahrscheinlich fehlen – unabhängig vom zugrunde gelegten geologischen Zeitmodell – zu viele paläontologische Daten, um die enormen Unterschiede in der Fossilüberlieferung der Vögel in Laurasia und Gondwana verstehen zu können. Dies mahnt zur Vorsicht bei der Deutung auf der Grundlage weniger Fossilien – insbesondere, wenn es um weitreichende Herkunftsmodelle geht.
Literatur
Martin AJ, Lowery M, et al. (2023) Earliest known Gondwanan bird tracks: Wonthaggi Formation (Early Cretaceous), Victoria, Australia. PLoS ONE 18(11): e0293308. https://doi.org/10.1371/journal.pone.0293308.
Autor dieser News: Reinhard Junker
© 2023, http://www.genesisnet.info/schoepfung_evolution/n326.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
13.11.23 Ein neues Bindeglied zwischen Dinosauriern und Vögeln?
Erneut wurde ein Fossil entdeckt, dessen Merkmalskombination zwischen Dinosauriern und Vögeln vermitteln soll – Fujianvenator prodigiosus. Die neue Art hat relativ vogelähnliche Vorderextremitäten, jedoch ungewöhnlich lange Beine. Dieses auffällige Merkmal war von „Urvögeln“ bisher weder bekannt noch erwartet worden; und es passt auch nicht in bisherige evolutionäre Vorstellungen. Die Beschreiber deuten die Merkmalskombination als „evolutionären Mosaizismus“ (evolutionär bedingte mosaikartige Verteilung von Merkmalen). Damit wird jedoch nichts erklärt und das dahinterstehende Problem für evolutionäre Erklärungen wird verschleiert.
Als evolutionäres „Bindeglied zu den Vögeln“ stellt wissenschaft.de die neu entdeckte langbeinige Dinosaurier-Art Fujianvenator prodigiosus vor (Bernard 2023; Xu et al. 2023). Welche Anforderungen sollte eine Art erfüllen, damit sie als Bindeglied in einer evolutiven Abfolge interpretiert werden kann? Als eine Minimalbedingung kann genannt werden: Die Merkmalskombination eines Bindeglieds sollte einigermaßen widerspruchsfrei zwischen zwei Arten passen, zwischen denen die betreffenden verbindende Art vermittelt. Und wenn die Baupläne des Lebens auf evolutivem Wege entstanden sind, sollte man zahlreiche solche passenden Bindeglieder finden.
Die fasanengroße neue Art, deren Name „bizarrer Jäger aus Fujian“ bedeutet und die als Bindeglied angesehen wird, wurde in Schichten des Oberjura entdeckt und hat demnach ein ähnliches Alter wie der berühmte Urvogel Archaeopteryx. Das Fossil ist relativ gut erhalten; es fehlen aber Kopf, Hals und das Ende des Schwanzes.
Rekonstruktionen von Fujianvenator zeigen ein prächtig entwickeltes Federkleid aus flächigen Federn. Das Fossil selbst weist jedoch gar keine Federn auf. Der Besitz von Federn wird nur angenommen, weil seine nächsten Verwandten der Anchiornithidae Federn besitzen (Dunham 2023). Als vogelartig wird das Fossil eingestuft, da sein Körperbau eine Mischung aus Merkmalen der Avialae (Vögel und nächste Verwandte) sowie zu den Dinosaurierfamilien der Troodontidae und Dromaeosauridae aufweist (Xu et al. 2023). Sein Becken besitzt Merkmale, die zur Diagnose von Anchiornis und der Troodontidae verwendet werden (zur Systematik s. Abb. 1).

Abb. 1: Stellung von Fujianvenator im Cladogramm nach Xu et al. (2023). (Wikimedia: CC BY-SA 4.0: Audrey.m.horn; Fred Wierum; UnexpectedDinoLesson; Luxquine; CC BY 4.0: PaleoNeolitic; CC BY-SA 3.0: Matt Martyniuk; El fosilmaníaco; Cladogramm nach: https://en.wikipedia.org/w/index.php?title=Fujianvenator&oldid=1176509176, CC BY-SA 4.0)
Die Vorderextremitäten von Fujianvenator sind im Wesentlichen wie das Skelett eines Vogelflügels gebaut, allerdings mit drei Krallen an den Fingern, die bei heutigen Vögeln ebenso wie bei den meisten fossilen Formen fehlen. Eine Ausnahme stellen die Kletterkrallen bei Jungvögeln des Hoatzins dar (s. Abb. 2): „[…] Krallen charakterisierten Archaeopteryx […]. Seltsamerweise haben die Jungtiere des Hoatzins (Opisthocomus hoazin) ähnliche Krallen an ihren Flügeln, verlieren sie aber im Erwachsenenalter“ (Naeem & Post 2019). Auch die Proportionen der Handknochen von Fujianvenator ähneln denen von Archaeopteryx. Aufgrund der Merkmale des gesamten Skeletts ist Fujianvenator dennoch wahrscheinlich nicht flugfähig. So hat Fujianvenator ein relativ kleines Schulterblatt, was gegen Flugfähigkeit spricht, da ein vergrößertes Schulterblatt mit den Muskeln verbunden ist, die für den Kraftflug erforderlich sind. Details der Mittelhandknochen deuten darauf hin, dass Fujianvenator flexible Greiffinger besaß, die beim Ergreifen von Beutetieren hilfreich gewesen sein könnten, während bei verwandten Vogelarten die Mittelhandknochen eine einzige unbewegliche Einheit bilden, an denen die Flugfedern befestigt sind (Xu et al. 2023, 341). Nimmt man alle diese Merkmale zusammen, sind die Rekonstruktionen von Fujianvenator mit einem Federkleid irreführend. Die Einstufung als Bindeglied steht schon von daher auf schwachen Beinen.

Abb. 2: Jungtier des Hoatzin mit Krallen. (Amazon-Images / Alamy Stock Photo)
Apropos Beine. Diesbezüglich ist Fujianvenator außergewöhnlich. Denn das Tier besaß sehr lange Beine und ähnelte daher heutigen Lauf- oder Watvögeln. Das Schienbein ist etwa doppelt so lang wie der Oberschenkelknochen. „Betrachtet man […] nur die Hinterbeine, so liegt Fujianvenator weit entfernt von allen anderen Theropoden, einschließlich anderer jurassischer Avialae“, stellen Xu et al. (2023, 341) fest. Ob die Merkmale der Beine eher auf eine Lebensweise im Sumpf oder auf schnelles Laufen hinweisen, ist mangels schlechter Erhaltung der Zehen nicht entscheidbar (Xu et al. 2023, 342; Coleman 2023). Weitere Fossilien aus der gleichen Fundstelle deuten aber darauf hin, dass Fujianvenator in einem sumpfigen Ökosystem lebte. Damit unterscheidet er sich von anderen Formen, die als Vorfahren der Vögel diskutiert werden und die an ein Leben auf Bäumen und in der Luft angepasst waren. Diese ökologische Nische war bisher bei den Avialae unbekannt.
Fujianvenator wird von den Forschern zu den Anchiornithidae gestellt (Xu et al. 2023), die innerhalb der Avialae eine Schwestergruppe zu Archaeopteryx und allen anderen Vogelarten darstellen (s. o. Abb. 1). In evolutionäre Interpretation befindet sich Fujianvenator also auf einem Seitenast.
Kommentar
Min Wang, einer der Bearbeiter, wird mit den Worten zitiert: „Auf die Frage nach einem Wort, das Fujianvenator beschreiben würde, würde ich sagen ‚bizarr‘. Fujianvenator ist weit davon entfernt, modernen Vögeln zu ähneln“ (zit. in Coleman 2023). Einmal mehr wurde mit Fujianvenator ein Fossil entdeckt, dessen Merkmalskombination evolutionär nicht erwartet wurde. Offensichtlich ist Fujianvenator kein Bindeglied hin zu modernen Vögeln, dafür ist er zu „bizarr“ – konkret: Die langen Beine und der mutmaßliche Lebensraum passen nicht für eine Interpretation als Bindeglied. Fujianvenator erweitert somit unsere Formenkenntnis über Dinosaurier und Vögel, doch er füllt keine bisher vorhandene evolutionäre Lücke. Vielmehr führt das Fossil in evolutionärer Perspektive zu einer zusätzlichen Lücke im evolutionären Stammbaum – nämlich von den bereits bekannten Anchiornithidae hin zu Fujianvenator mit seinem einzigartigen Merkmalsmosaik.
Fujianvenator steht hingegen – wie viele andere Gattungen auch – dafür, dass Merkmale frei kombinierbar sind. Davon kann aber nur in einem Schöpfungsmodell ausgegangen werden. Xu et al. (2023, 340) sprechen von einem „evolutionären Mosaizismus“: „Die einzigartige Kombination postkranialer (Körperskelett-)Merkmale, die Fujianvenator mit frühen Paraves gemeinsam hat, zeigt, wie stark die Phylogenie [Stammesgeschichte] der Avialier durch evolutionären Mosaizismus beeinflusst wurde“ (zu „Paraves“, inklusive Deinonychosauria, und „Avialae“ siehe oben Abb. 1). Doch das ist nur ein Schlagwort, das nichts erklärt und das evolutionäre Problem verschleiert. Wie kann „Mosaizismus“ – mosaikartige Verteilung von Merkmalen – etwas beeinflussen? Es ist die freie Kombinierbarkeit von Merkmalen, die zu einer mosaikartigen Verteilung von Merkmalen verschiedener Arten oder größerer Gruppen führt und gerade nicht bekannte evolutionäre Prozesse; daher ist dies evolutionär nicht zu erwarten. „Mosaizismus“ bedeutet bei einer „Übersetzung“ in ein Cladogramm bzw. in einen Stammbaum, dass Konvergenzen angenommen werden müssen, also das mehrfach unabhängige Auftreten gleicher Merkmale. Durch einen evolutionären Prozess, der nicht zielorientiert verläuft, sind Konvergenzen aufgrund der fehlenden Steuerung aber gerade nicht zu erwarten (vgl. Junker 2016).
Quellen
Bernard E (2023) Langbeiniger Dinosaurier als Bindeglied zu den Vögeln, vom 06.09.2023,https://www.wissenschaft.de/erde-umwelt/langbeiniger-dinosaurier-als-bindeglied-zu-den-voegeln/
Coleman L (2023) ‘Weird’ dinosaur prompts rethink of bird evolution. Nature News, doi: 10.1038/d41586-023-02757-5.
Dunham W (2023) ‘Bizarre’ long-legged bird-like dinosaur has scientists enthralled, vom 06.09.2023https://www.reuters.com/science/bizarre-long-legged-bird-like-dinosaur-has-scientists-enthralled-2023-09-06/.
Naeem S & Post K (2019) How wings lost their claws. Science 364, 746–748, doi: 10.1126/science.2019.364.6442.twis.
Junker R (2016) Evolution „erklärt“ Sachverhalte und ihr Gegenteil. Stud. Integr. J. 23, 4–12.
Xu L, Wang M et al. (2023) A new avialan theropod from an emerging Jurassic terrestrial fauna. Nature 621, 336–343, doi:10.1038/s41586-023-06513-7.
Autor dieser News: Reinhard Junker
© 2023, http://www.genesisnet.info/schoepfung_evolution/n324.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
31.03.23 Cratonavis: Mosaik aus T. rex und Vogel?
Vom Umgang mit unerwarteten Vogel-Fossilfunden
Ein jüngst entdecktes Fossil überrascht mit einer ungewöhnlichen Merkmalskombination: Ein Kopf, der in seiner Form Tyrannosaurus rex ähnelt, kombiniert mit einem Vogelrumpf. Der Fund wird zwar als weiteres Puzzlestück der Vogel-Evolution bezeichnet, steht wegen ausgeprägter Konvergenzen tatsächlich aber quer zu stammesgeschichtlichen Hypothesen. Wie gehen die Forscher damit um?
Einführung
In populären Darstellungen wird oft behauptet, Fossilfunde bestätigten laufend die Vorstellung von einer Evolution der Lebewesen und neue Funde füllten Lücken im Stammbaum. Dass ein Fossil irgendeine Lücke füllt, ist trivial, wenn man voraussetzt, dass es einen Stammbaum überhaupt gibt. Die Frage ist hingegen: Füllen neuentdeckte Fossilien bisher vorhandene Lücken? Das ist in sehr vielen Fällen – vielleicht sogar in der Mehrzahl – durchaus nicht der Fall. Nicht umsonst erhalten neue Fossilfunde regelmäßig das Etikett „überraschend“, oder die Merkmale oder Merkmalsmosaike neuer Funde werden als „sonderbar“ oder „bizzar“ beschrieben. Das heißt, die betreffenden Merkmalskonstellationen waren evolutionstheoretisch nicht vorhergesagt worden und sind unerwartet. Natürlich widerlegen sie Evolutionstheorien als solche nicht, aber sie bestätigen sie auch nicht. Der vermutete historische Ablauf wird umfangreicher und komplexer und die Erklärungsleistung des Konzeptes damit geringer. Schließlich kann man immer den bisher bevorzugten Stammbaum ändern und an neue Funde anpassen – allerdings mit dem „Preis“, dass immer mehr Konvergenzen* angenommen werden müssen, also das unabhängige Auftreten ähnlicher Merkmale. Nicht selten kommt es vor, dass Merkmale, deren Ähnlichkeit zunächst als abstammungsbedingt interpretiert wurde (Homologien), später als Konvergenzen neu interpretiert werden müssen. Eine logische Schlussfolgerung dieser Situation ist, dass Bauplanähnlichkeiten kein sicherer Indikator für Evolution sein können.
*Konvergenzen (= gleichartige Ausprägung von Strukturen abstammungsmäßig nicht verwandter Organismen, die evolutionstheoretisch als unabhängig entstanden interpretiert werden)
Cratonavis
Ein aktuelles Beispiel für diese Situation ist ein kürzlich gemachter Fossilfund eines knapp habichtgroßen Vogels, der etwas schrill als „Urvogel mit Dino-Kopf“ (Manz 2023) oder als „seltsamer primitiver Vogel mit einem T. rex-Kopf“ (Puiu 2023) bezeichnet wurde. Das vollständig erhaltene Fossil wurde von Forschern der Chinese Academy of Sciences untersucht und als neue Art Cratonavis zhui beschrieben; sie haben es der neu aufgestellten Gattung Cratonavis zugeordnet (Abb. 1).

Abb. 1: Künstlerische Darstellung von Cratonavis zhui. (Bild: Chuang Zhao, CC BY-SA 3.0)
Cratonavis wurde in den Jiufotang-Schichten Nordchinas entdeckt, die auf 120 Millionen radiometrische Jahre (MrJ) datiert werden (Li et al. 2023). Sein Körper war vogelähnlich, der Kopf jedoch ausgesprochen untypisch für Vögel und dinosaurierartig. Insgesamt war der Kopf dem des Tyrannosaurus rex recht ähnlich hinsichtlichreptilientypischen Schläfenfenstern (diapside* Ausprägung), Gaumenregion sowie unbeweglichem Oberkiefer. Cratonavis konnte daher seinen Oberschnabel nicht unabhängig von Hirnschale und Unterkiefer bewegen, anders als die meisten heute lebenden Vögel.
*diapsid (= mit zwei Schläfenfenstern des Schädeldachs ausgestattet)
Zusammen mit den Gattungen Jinguofortis und Chongmingia wird Cratonavis in die erst vor wenigen Jahren aufgestellte Familie Jinguofortisidae gestellt. Diese Familie wird in den Bereich zwischen Archaeopteryx und der Vogelgruppe der Ornithothoraces (Gruppe aus ausgestorbenen Gegenvögeln (Enantiornithes) und Ornithuromorpha*) eingeordnet (s. Abb. 2).
*Ornithuromorpha (= Vögel mit Fächerschwanz, zu denen auch die heutigen Vögel gehören)
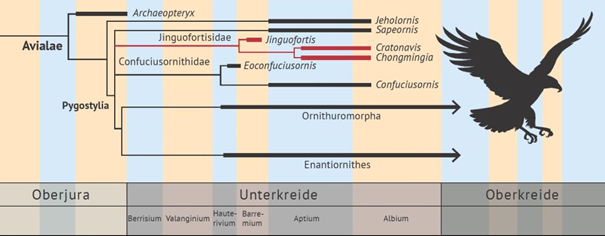
Abb. 2: Einfaches Cladogramm der Vögel mit der erst vor kurzem aufgestellten Familie Jinguofortisidae, zu der der neue Fund Cratonavis gestellt wird. (Ausschnitt eines umfangreichen Cladogramms nach Li et al. 2023)
In der Meldung von scinexx.de (Manz 2023) wird behauptet, dass ausgehend von zweibeinigen Raub-Dinosauriern (Theropoden) die Formen, die zu den Vögeln führten, „Schritt für Schritt mehr den heutigen Vögeln ähnelten“. Diese Behauptung findet sich auch in der Fachliteratur (z. B. Brusatte et al. 2014). Der neue Fund scheint das auf den ersten Blick zu bestätigen: Dino-Kopf und Vogelkörper. Eine detaillierte Analyse der Merkmalsverteilungen der Theropoden-Familien zeigt jedoch, dass ein „schrittweiser“ Übergang nicht den Fossilbefunden entspricht, denn die vogelartigen Merkmale bei Dinosauriern sind sehr unsystematisch unter den verschiedenen Gruppen der Theropoden verteilt, was sich in einer auffällig großen Zahl von Konvergenzen äußert (Junker 2019, Tab. 2 und 3, S. 46). Viele der untersuchten Vogelmerkmale, die bei Theropoden vorkommen, sind gar nicht bei denjenigen Formen ausgebildet, die als unmittelbare Vogelvorläufer angesehen werden. Stattdessen müssten sie – unter Voraussetzung von Evolution – konvergent entstanden sein und zwar zu einem Zeitpunkt, als es längst schon Vögel gab. Und die meisten anderen untersuchten Merkmale gelten als allgemeine und verbreitet vorkommende Theropodenmerkmale und sind daher ebenfalls nicht gut geeignet, eine schrittweise Entstehung von typischen Vogelmerkmalen bei Theropoden zu belegen.
Der neue Fund Cratonavis bestätigt dieses Bild von großen Unstimmigkeiten: Es handelt sich zwar um eine ausgeprägte Mosaikform, aber sie passt nicht in eine lineare Reihe, die „Schritt für Schritt“ von Dinosauriern zu Vögeln führt. Denn der „Dino-Kopf“ steht gleichsam quer dazu, weil der etwa 30 MrJ ältere „Urvogel“ Archaeopteryx einen deutlich vogeltypischeren Schädel hatte (Martin 1985; Elzanowski & Wellnhofer 1995; 1996; Wellnhofer 2009), während sein Rumpf umgekehrt weniger vogeltypisch war als der von Cratonavis. Letzterer besaß ein Pygostyl*, das Archaeopteryx fehlte, der stattdessen einen lange Schwanzwirbelsäule hatte (s. Abb. 3).
*Pygostyl (= Durch die Verschmelzung mehrerer Wirbel gebildeter Knochen am Ende der Wirbelsäule von Vögeln)

Abb. 3: Neuere Rekonstruktion von Archaeopteryx. Sie berücksichtigt die in jüngeren Studien gezogene Schlussfolgerung, dass mindestens ein Teil des Gefieders mit hoher Wahrscheinlichkeit schwarz war. (Nobu Tamaura, CC BY-SA 3.0)
Dazu kommt, dass Cratonavis einzigartige Merkmale (sog. Autapomorphien) besaß, die nicht in eine Evolutionsreihe hin zu den heutigen Vögeln passen: Die Schulterblätter und die Mittelfußknochen waren ungewöhnlich stark verlängert, so dass auch der vogeltypisch abstehende Hallux (großer Zeh) ziemlich lang war. Diese Ausprägungen werden als konvergente Bildungen angesehen; man kennt verlängerte Schulterblätter sonst bei nicht näher verwandten Ornithuromorphen* und verlängerte Mittelfußknochen beim Dromaeosauriden Balaur aus der Oberkreide (Li et al. 2023, 25, 26). Cratonavis könnte seine langen Zehen benutzt haben, um wie die heutigen Raubvögel zu jagen. Und die Ausbildung eines stark verlängerten Schulterblatts könnte das Fehlen eines verknöcherten Brustbeinkiels kompensiert haben, da es eine zusätzliche Fläche für die Befestigung des Brustmuskels (Pectoralis-Muskel) bietet (Li et al. 2023, 26).
Das komplette Merkmalsmosaik verkleinert insgesamt nicht eine vorhandene Lücke, sondern erfordert evolutionstheoretisch einen zusätzlichen Ast im hypothetischen Stammbaum (Familie Jinguofortisidae; s. o. Abb. 2).
Jinguofortis
Cratonavis wird zusammen mit der 2016 erstmals beschriebenen Gattung Jinguofortis und der Gattung Chongmingia in die neu aufgestellte Familie Jinguofortisidae gestellt. Auch Jinguofortis überraschte die Forscher mit einer ungewöhnlichen Merkmalskonstellation von theropoden- und vogeltypischen Merkmalen (Wang et al. 2016; 2018). Einerseits besaß diese Gattung ein Pygostyl und stark reduzierte Finger, andererseits Krallen an den Fingern der Flügel, ein bumerangförmiges, vermutlich starres Gabelbein und ein verschmolzener Schultergürtel (Schulterblatt und Rabenbein). Letzteres Merkmal erscheint zwar ungünstig für das Fliegen, weil es die Flexibilität für den Schlagflug einschränkt, dennoch waren die breiten, kurzen Flügel von Jinguofortis typisch für Vögel, die gut zwischen Bäumen manövrieren können. Vielleicht war eine bisher unbekannte Art des Fliegens verwirklicht. Der verschmolzene Schultergürtel (Schulterblatt und Rabenbein) ähnelt zwar der Situation bei einigen Theropoden-Dinosauriern, trotzdem eignet sich dieses Merkmal nicht als Beleg für eine stammesgeschichtliche Verbindung von Dinosauriern und Vögeln, weil beim über 20 MrJ älteren Archaeopteryx diese beiden Knochen nicht verschmolzen waren. Daher wird eine unabhängige (konvergente) Entstehung angenommen.
Wang et al. (2018, 10708) bemerken, dass Jinguofortis die bekannte Diversität (Vielfalt) früher Pygostylier (Vögel mit Pygostyl) vergrößere, und vermuten, dass Entwicklungs-Plastizität eine wichtige Rolle spielte und die mutmaßliche Evolution mosaikartig verlief. Die Jinguofortisidae trügen zum verbreiteten Vorkommen von Mosaik-Evolution bei (Wang et al. 2018, 10710).
Li et al. (2023, 24) stellen fest, dass diese Situation durch Cratonavis noch komplizierter werde. Die Unstimmigkeiten in den Merkmalsverteilungen haben mit dieser Gattung weiter zugenommen.
Evolutionstheoretische Deutung und die Alternative
Li et al. (2023, 20, 26) interpretieren den neuen Fossilfund so, dass er den „Einfluss von Mosaik-Evolution und biologischer Experimentierung“ im Flugverhalten widerspiegle. Die Vögel, die sich nahe am mutmaßlichen evolutionären Ursprung befinden, zeigten unterschiedliche Merkmale, mit denen sie den vermeintlich unterentwickelten Flugapparat kompensieren konnten, von denen einige bei heutigen Vögeln unbekannt seien. „Mosaik-Evolution“ und „Experimentierung“ sind jedoch Fremdkörper in einem evolutionären Szenario. Denn Experimente sind absichtsvolle Unternehmungen zur Klärung von konkreten Fragestellungen; die gewählten Begriffe verschleiern einen evolutionstheoretisch unerwarteten Befund. Die Merkmalsverteilung passt nicht in ein hierarchisches eingeschachteltes System; daher müssen in großem Maße Konvergenzen angenommen werden, was als „Mosaik-Evolution“ bezeichnet wird. Aber warum und auf welchem Wege gelangt ein natürlicher, zukunftsblinder evolutionärer Prozess vielfach unabhängig zu ähnlichen Konstruktionen? Und was soll es bedeuten, dass Mosaik-Evolution einen „Einfluss“ gehabt habe?
Noch weiter geht Puiu (2023) in seinem Newsbeitrag. Die Evolution verlaufe auf einem verschlungenen Pfad, auf dem sie oft einen Schritt vorwärts, aber zwei Schritte zurück machen müsse. Der Dino-Vogel-Mischmasch sei nicht völlig unerwartet. Es gebe immer mehr Belege dafür, dass der Übergang von den Theropoden zu den Vögeln weder glatt noch nahtlos war, sondern durch zahlreiche Wiederholungen und evolutionäre Experimente erfolgte, die zu einer Reihe primitiver Vögel mit mosaikartigen Merkmalen führten.
Genau das würde man aber wegen der Richtungslosigkeit evolutionärer Mechanismen nicht erwarten, weil diese Situation mit zahlreichen Konvergenzen einhergeht – ein Befund, der früheren evolutionstheoretischen Erwartungen diametral entgegensteht. Doch nun wird der Bock zum Gärtner gemacht: Unerwartete Befunde werden kurzerhand als Erwartungen umgedeutet – ohne jede Begründung. Man könnte auch sagen: Weil man schon öfter nicht passende Funde hatte, erwartet man solche auch zukünftig, bezeichnet diese dann aber neuerdings als Bestätigungen der eigenen Perspektive.
Es gibt allerdings eine konsistente alternative Deutung: die freie Verwendbarkeit und Kombinierbarkeit von Merkmalen. Diese Freiheit besitzt nur ein Schöpfer. Evolution ist dagegen an einen natürlichen Prozess und an natürliche Mechanismen gebunden. Ein evolutionärer Prozess ohne eingebaute Ziele lässt aber – selbst wenn er überhaupt funktionieren würde – keine ähnlichen komplexen Merkmale erwarten, die mehrfach und unabhängig voneinander auftreten.
Quellen
Brusatte SL, Lloyd GT, Wang SC & Norell MA (2014) Gradual assembly of avian body plan culminated in rapid rates of evolution across the dinosaur-bird transition. Curr. Biol. 24, 2386–2392.
Elzanowski A & Wellnhofer P (1996) Cranial morphology of Archaeopteryx: evidence from the seventh skeleton. J. Vert. Paleont. 16, 81–94.
Elzanowski A & Wellnhofer P (1995) The skull of Archaeopteryx and the origin of birds. Archaeopteryx 13, 41–46.
Junker R (2019) Sind Vögel Dinosaurier? Eine kritische Analyse fossiler Befunde. Internetartikel. https://www.wort-und-wissen.org/wp-content/uploads/b-19-4_dinos-voegel.pdf
Li Z, Wang M, Stidham TA & Zhou Z (2023) Decoupling the skull and skeleton in a Cretaceous bird with unique appendicular morphologies. Nat. Ecol. Evol, doi: 10.1038/s41559-022-01921-w.
Manz A (2023) Urvogel mit Dino-Kopf entdeckt. https://www.scinexx.de/news/biowissen/urvogel-mit-dino-kopf-entdeckt/.
Martin LD (1985) The relationship of Archaeopteryx to other birds. In: Hecht MK, Ostrom JH, Viohl G & Wellnhofer P (Hg) The beginnings of birds. Eichstätt: Freunde Jura-Museums, pp. 177–183.
Puiu T (2023) This weird primitive bird with a T. rex head has scientists puzzled. https://www.zmescience.com/science/news-science/this-weird-primitive-bird-with-a-t-rex-head-has-scientists-puzzled/.
Wang M, Wang X, Wang Y & Zhou Z (2016) A new basal bird from China with implications for morphological diversity in early birds. Sci Rep. 6:19700, doi: 10.1038/srep19700.
Wang M, Stidham TA & Zhou Z (2018) A new clade of basal Early Cretaceous pygostylian birds and developmental plasticity of the avian shoulder girdle. Proc. Natl. Acad. Sci. 115, 10708–10713.
Wellnhofer P (2009) Archaeopteryx. The icon of evolution. München: Pfeil.
Autor dieser News: Reinhard Junker
© 2023, http://www.genesisnet.info/schoepfung_evolution/n312.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
23.11.22 Flugsaurier mit Federn?
Begriffsverwirrung führt zu verwirrenden Aussagen
Bei einigen Flugsaurierfossilien wurden haarartige, büschelige und fiederige Strukturen auf der Haut entdeckt, die als „Federn“ interpretiert werden. Demnach sollen Federn schon lange vor dem Auftreten von Vögeln entstanden sein. Die betreffenden Körperanhänge sind jedoch sehr klein. Sie als „Federn“ zu bezeichnen, ist evolutionstheoretisch motiviert und durch den Aufbau der Strukturen nicht gerechtfertigt. Die Bezeichnung „Feder“ ist daher irreführend; das führt zu entsprechend irrigen Schlussfolgerungen. Tatsächlich waren Flugsaurier nach gegenwärtigem Kenntnisstand nicht befiedert. Zudem spricht die Verteilung der Ausprägungen von Hautstrukturen für eine freie Kombinierbarkeit von Merkmalen, die einem Schöpfer zur Verfügung steht, nicht aber einer Evolution, die an bestimmte Mechanismen gebunden ist.
Flugsaurier gehören zu den seltsamsten und ausgefallensten Geschöpfen. Ihre Körperproportionen sind ungewöhnlich: Ein großer Kopf, teils mit extravaganter Ornamentierung und oft mit spitzer Schnauze, gepaart mit einem langen Hals und einem kleinen Körper und kurzen Beinen. Ihre Flugfähigkeit verdanken sie einer Flughaut, die vor allem am extrem langen vierten Finger und am Arm sowie an der kurzen Körperseite aufgespannt war. Schon seit der Entdeckung der ersten Flugsaurier ist bekannt, dass die die Körperhaut und teilweise auch die Flughaut mit haarartigen Auswüchsen, sog. Pyknofasern, bedeckt war. Pyknofasern sind kurz, weisen einen in der Mitte verlaufenden Kanal auf und sind nicht in die Haut eingesenkt wie die tief verwurzelten Haare der Säugetiere (Anonymus 2018, 301). Vermutlich dienten sie der Wärmeisolierung.
Federn bei spatzengroßen Flugsauriern?
Die Pyknofasern erhielten in den letzten Jahren neue Aufmerksamkeit. Yang et al. (2019) berichteten von vier verschiedenen Typen haarartiger (oder federartiger?) Körperbedeckung bei zwei Exemplaren der Flugsaurier-Familie der Anurognathidae, deren Alter mit 160 Millionen radiometrischen Jahren (MrJ) bestimmt wurde. Die Anurognathidae gehören zu den kleinsten Flugsauriern und waren nur spatzengroß. Bei ihrer Körperbedeckung handelte sich zum einen um die schon lange bekannten einfachen Fasern, zum anderen wurden andere Typen von Anhängen entdeckt, die bisher bei Flugsauriern unbekannt waren. Es handelt sich 1. um pinselförmig gebündelte Fasern, 2. um Anhänge, die auf halber Höhe büschelig werden, und 3. um daunenförmig verzweigte Anhänge an den Flügelhäuten. Solche Strukturen sind auch bei Vögeln oder bei Dinosauriern bekannt, die als Vogelvorläufer gelten. Darüber hinaus wurden im Inneren der Fasern und in der Haut Melanosomen in verschiedenen Formen entdeckt. Diese Strukturen enthalten den Farbstoff Melanin und bestimmen bei heutigen Vögeln die Farbe des Gefieders. Die Autoren interpretieren diese Befunde insgesamt als Belege dafür, dass bereits Flugsaurier Federn besaßen, und vermuten einen gemeinsamen Ursprung von Federn beim gemeinsamen Vorfahren der Dinosaurier und Flugsaurier vor etwa 250 MrJ. Eine unabhängige Entstehung der „Federn“ wäre zwar auch denkbar, wurde von den Autoren jedoch wegen der Ähnlichkeiten mit Integumentstrukturen (= Strukturen auf der Körperhülle) von Dinosauriern als unwahrscheinlich eingestuft. Die Deutung der Strukturen als Körperanhänge erfuhr zwar Kritik (Unwin & Martill 2020), diese wurde aber von den Autoren Punkt für Punkt zurückgewiesen (Yang et al. 2020).
Tupandactylus
Nun wurde jüngst von einem weiteren Typ einer Körperbedeckung bei einem anderen, sehr viel größeren Flugsaurier berichtet. Es handelt sich um den Fund eines unvollständigen Schädels von Tupandactylus imperator aus der Unterkreide Brasiliens (113 MrJ). Tupandactylus besaß eine Flügelspannweite von 3–5 Metern. Auffällig ist ein von zwei Knochenspornen aufgespannter, mehrere Dezimeter langer Hautkamm auf dem Kopf, der wahrscheinlich eine Art Schau-Element darstellte. An dessen Basis fanden sich Überreste von teils fädigen, teils flauschigen, verzweigten Strukturen. Eine elektronenmikroskopische Untersuchung offenbarte, dass von einem zentralen, schwer erkennbaren Schaft, der sich zur Spitze hin verjüngte, über fast die gesamte Länge kurze gerade Fasern abgingen (Abb. 1).

Abb. 1: a Unvollständiger Schädel mit erhaltenem Weichteilkamm; b-f Detail der mit dem hinteren Teil des Schädels verbundenen Integumentstrukturen; b Monofilamente; c verzweigte Federn; d Detail der gebogenen verzweigten Feder in c; e, f gerade verzweigte Feder (e) mit Detail (f). Die weiße Pfeilspitze in e zeigt den basalen Calamus an (der Calamus ist der eingesenkte Teil des Federschafts). Balken: 50 mm (a), 5 mm (b), 2 mm (c), 0,25 mm (d-f). (Aus CINCOTTA et al. 2022, CC BY 4.0)
Die Autoren bezeichnen diese Strukturen daher als „Federn“, die dem Stadium 3b des evolutionären Feder-Entstehungsmodells von Prum (1999) gleichen, weil hier Federäste regelmäßig von einem zentralen Schaft abzweigen (vgl. Abb. 2).
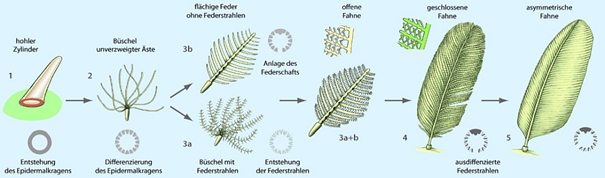
Abb. 2: Evolutionäres Feder-Entstehungsmodell nach PRUM (1999). Relevant für die Diskussion um den besonderen Federtyp von Tupandactylus ist Stadium 3b (bei CINCOTTA et al. als IIIa benannt) mit fiederiger Verzweigung. (Nach PRUM & BRUSH 2003 und PERRICHOT et al. 2008; 4-5 von Emily WILLOUGHBY, https://emilywilloughby.com, mit freundlicher Genehmigung)
Entsprechend lauteten die Pressemeldungen, dass „Flugsaurier mit Federn“ entdeckt worden seien. Auch hier wurden Melanosomen entdeckt. Dabei befinden sich in den Fasern, in den verzweigten Anhängen und in der Haut verschiedene Formen von Melanosomen. Da die Melanosomen mit der Farbmusterung zusammenhängen, wird aus diesem Befund geschlossen, dass verschiedene Regionen des Hautkamms von Tupandactylus unterschiedlich gefärbt waren, was dessen Signalwirkung verstärkt haben dürfte. Es könnte eine fleckige Verteilung von Farben auf der Oberfläche des Hautkamms gegeben haben (Benton 2022; vgl. Abb. 3).

Abb. 3: Rekonstruktion des Kopfes von Tupandactylus. (Julio Lacerda, CC BY 4.0)
Cincotta et al. (2022) kommen wie Yang et al. (2019) zum Schluss, dass federartige Strukturen bereits vorhanden waren, bevor sich die Linien der Flugsaurier und der Dinosaurier trennten – vor etwa 250 MrJ. Dass zumindest die fädigen und büscheligen Anhänge auf einen gemeinsamen Vorfahren beider Sauriergruppen zurückgehen, wurde durch eine Stammbaumanalyse mit 84-prozentiger Wahrscheinlichkeit (unter Annahme einer gemeinsamen Evolutionsgeschichte der Lebewesen) unterstützt. Komplexere Strukturen sollen aber in beiden Gruppen unabhängig entstanden sein (Cincotta et al. 2022). Auch diese Autoren vermuten, dass die „Federn“ der Wärmeisolation dienten und vielleicht auch als Elemente der visuellen Kommunikation fungierten.
Diskussion
Wer denkt bei „Federn“ nicht zuerst an die flächigen Vogelfedern, die Teil eines komplexen Flugapparats bilden? Meldungen wie „Federn bei Flugsauriern“ wirken daher irreführend. Die Strukturen, die Cincotta et al. (2022) als Federn identifizierten, sind sehr klein, insbesondere im Verhältnis zu dem gewaltigen Kopfkamm des Tieres, an dessen Rückseite sie gefunden wurden. Der zentrale Schaft besaß einen Durchmesser von etwa 60 μm; die abzweigenden Fasern waren etwa 0,1–0,2 mm lang. Die Verankerung im Körper ist unklar. Cincotta et al. verweisen darauf, dass ein Calamus (der eingesenkte Teil des Schafts) zu sehen sein könnte (s. o. Abb. 1e, weißer Pfeil). Die Art der Verankerung wäre ein wichtiges Indiz für eine Vergleichbarkeit mit Federn (es sei an die oberflächlichen Verankerungen von Pyknofasern erinnert; s. o.).
Die Anhänge der von Yang et al. (2019) beschriebenen vier Typen von Körperanhängen sind ähnlich klein. Ohne die evolutionäre Sichtweise, dass Vögel von Dinosauriern abstammen, würde man auf der Basis des morphologischen Befundes an sich, also aufgrund der Formen dieser Gebilde, wohl kaum auf die Idee kommen, man habe es mit „Federn“ zu tun. Nun aber ähneln die verzweigten Fasern solchen, die beim Dromaeosauriden Sinornithosaurus millenii gefunden wurden (Abb. 4). Die Dromaeosauriden wiederum werden in einen weiteren Abstammungszusammenhang mit Vögeln gestellt (obwohl sie in deutlich jüngeren Schichten gefunden wurden als Formen mit echten, flächigen, flugtauglichen Federn). Auf diese indirekte Weise wird somit der theoretische Zusammenhang zu „Federn“ bei Flugsauriern hergestellt.

Abb. 4: Links: Integumentstrukturen von Sinornithosaurus millenii mit gefiederter Verzweigung, ähnlich Typ 3b nach PRUMs Feder-Entstehungsmodell. Rechts: Vollständiges Sinornithosaurus-Exemplar „Dave“. (Links: Aus XU et al. 2001; mit freundlicher Genehmigung; Fossil: Jonathan Chen, CC BY-SA 4.0)
Man kann hier durchaus von Begriffsverwirrung sprechen. Sie entsteht dadurch, dass „Federn“ letztlich nicht vorrangig aufgrund der morphologischen Merkmale definiert werden, sondern aufgrund vermuteter evolutionärer Zusammenhänge. Das Vorhandensein verschieden geformter Melanosomen und winziger verzweigter Fasern auf der Haut wird als ausreichend betrachtet, um von „Federn“ zu sprechen (Cincotta et al. 2022, 6871). Das erscheint reichlich fragwürdig. Die Autoren schließen allerdings die Hypothese nicht vollständig aus, dass die fadenförmigen Integumentstrukturen der Flugsaurier eine dritte Art von Wirbeltier-Integumentauswüchsen (neben Haaren und Federn) darstellen, die in der Lage sind, eine auf Melanin basierende Färbung zu vermitteln und zu variieren. Sie halten diese Deutung (vermutlich wegen des evolutionären Denkrahmens) aber für unwahrscheinlich.
Eine Folge der Begriffsverwirrung ist die verwirrende Aussage, Flugsaurier hätten Federn besessen. Man wird hier unwillkürlich denken, das habe etwas mit ihrer Flugfähigkeit zu tun. Aber wozu sollen Federn dienen bei einem Spezialisten, der mit einer ausgefeilten Flughaut exzellent fliegen kann (vgl. Pittman et al. 2021)? Zu einer Flugfähigkeit tragen diese Gebilde offensichtlich nichts bei.
Aufgrund der Verteilung der „Federn“ des Typs 3b am Körper müsste man – wie von Cincotta et al. auch angemerkt – von einer konvergenten Entstehung bei Flugsauriern und Dromaeosauriden ausgehen. Selbst wenn man also evolutionstheoretisch argumentiert, könnte gerade das interessante Stadium 3b, dem die Integumentstrukturen von Tupandactylus ähneln, nicht als Vorläufer von echten Federn interpretiert werden.
Der Befund an sich legt eher nahe, dass die dichte und auch verzweigte Behaarung eine flugsaurierspezifische Form der Wärmeisolierung war und eventuell Signalwirkung ermöglichte. Ein evolutionärer Zusammenhang drängt sich nicht auf. Im Gegenteil: Wenn „Federn“ vom Typ 3b konvergent auftreten, sind sie kein verlässliches Argument für eine gemeinsame Abstammung von Theropoden-Dinosauriern und Vögeln. Wahrscheinlich sind sie hingegen eines von vielen Merkmalen, die wie aus einem Baukastensystem von einem Schöpfer frei kombiniert wurden.
Anmerkung
1 Über die Anwesenheit verschieden geformter Melanosomen schreiben sie: „This further supports the hypothesis that the branched integumentary structures in pterosaurs are feathers.“
Literatur
Anonymus (2018) Fur and fossils. Nature 564, 301–302.
Benton MJ (2022) A colourful view of the origin of dinosaur feathers. Nature 604, 630–631.
Cincotta A, Nicolai M et al. (2022) Pterosaur melanosomes support signalling functions for early feathers. Nature 604, 684–688.
Pittman M, Barlowa LA , Kaye TG & Habib MB (2021) Pterosaurs evolved a muscular wing–body junction providing multifaceted flight performance benefits: Advanced aerodynamic smoothing, sophisticated wing root control, and wing force generation. PNAS 118, e2107631118.
Prum RO (1999) Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291–306.
Perrichot V, Marion L, Neraudeau D, Vullo R & Tafforeau P (2008) The early evolution of feathers: fossil evidence from Cretaceous amber of France. Proc. R. Soc. 275B, 1197-1202.
Prum RO & Brush AH (2003) Zuerst kam die Feder. Spektr. Wiss. 10/03, 32-41.
Unwin DM & Martill DM (2020) No protofeathers on pterosaurs. Nat. Ecol. Evol. 4, 1590–1591.
Xu X, Zhou ZH & Prum RO (2001) Branched integumental structures in Sinornithosaurus and the origin of feathers. Nature 410, 200–204.
Yang X, Jiang B et al. (2019) Pterosaur integumentary structures with complex feather-like branching. Nat. Ecol. Evol. 3, 24–30.
Yang Z, Jiang B et al. (2020) Reply to: No protofeathers on pterosaurs. Nat. Ecol. Evol. 4, 1592–1593.
Autor dieser News: Reinhard Junker
© 2022, http://www.genesisnet.info/schoepfung_evolution/n308.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
25.06.22 Stummelarme bei T. rex
„Aber Tyrannosaurus – warum sind deine Arme so schrecklich kurz?“- „Damit meine Kumpels nicht aus Versehen reinbeißen.“
Wissenschaft ist oft anstrengend, kann aber auch manchmal erheiternd sein. Jedenfalls hatte ein aktueller Vorschlag zur Lösung eines paläontologischen Rätsels auf mich diese Wirkung. Über den betreffenden Wissenschaftler will ich mich aber keineswegs lustig machen – im Gegenteil: Seine Kreativität in der Suche nach der Antwort auf die ur-biologische Frage „Wozu“ finde ich bewundernswert. Die Wozu-Frage, also die Frage nach dem Zweck einer biologischen Struktur, ist eine Haupttriebfeder biologischer Forschung. Diesbezüglich stellen die Lebewesen die Wissenschaftler häufig vor Herausforderungen. Zur Klärung der Bedeutung einer biologischen Struktur ist die Lebensweise von zentraler Bedeutung. Allerdings ist diese bei Lebewesen, die wir nur fossil kennen, oft schwer herauszufinden.
Bei den heutigen Lebewesen waren manche Evolutionsbiologen in der Vergangenheit zu schnell dabei, die Frage „wozu ist das gut?“ mit einem „zu gar nichts mehr“ zu beantworten. Nicht mehr zweckmäßig seien viele Strukturen von Lebewesen, weil sie im Laufe der angenommenen Evolution ihre Funktion verloren hätten. Das wohl berühmteste Beispiel ist der Wurmfortsatz (Appendix) des menschlichen Blinddarms, der keinen Nutzen zu haben schien – bis genauere Untersuchungen dies widerlegten (Bollinger et al. 2007; Smith et al. 2013; Ullrich 2013). Überhaupt ist die Annahme, es gebe funktionslose Organe, nicht bestätigt worden. Der Neugier der Wissenschaftler sei Dank.
Die Biologie lebt also gewissermaßen davon, Wozu-Fragen zu beantworten. Das ist ihr Spezifikum. Dahinter steht – meist unausgesprochen – die Auffassung, dass in der Biologie nichts dem Zufall überlassen ist, jedenfalls nichts Wesentliches. Andernfalls würde sich die Frage nach der Funktion eigentlich erübrigen. Da sich die Lebewesen die Zwecke ihrer Organe aber kaum selber ausgedacht haben, steht im Hintergrund der Frage „Wozu“ letztlich immer auch die Frage nach einem zielgerichtet denkenden und handelnden Schöpfer.
Ein schönes Beispiel dafür, dass biologische Forschung sich keinesfalls damit zufrieden gibt, nur zu beschreiben, was man vorfindet, ist einer der bekanntesten Dinosaurier: kein geringerer als der „König der Herrscherechsen“ – Tyrannosaurus rex. Dieser beeindruckende König scheint jedoch einen ausgeprägten und unübersehbaren Makel zu haben. Seine Vorderextremitäten sind allzu klein und sehen am mächtigen Körper geradezu kurios, ja fast lächerlich aus (s. Abb. 1). Sie reichten nicht einmal bis zum Maul. Die kurzen Arme sind deshalb „seit langem Gegenstand von Verwunderung, Spekulationen und sogar Spott von Seiten der Paläontologen und der Öffentlichkeit gewesen“ (Padian 2022, 63). Und natürlich begnügen sich die Biologen nicht mit der Feststellung: „Es ist halt so.“ Selbstverständlich wollen sie wissen, warum das so ist. „Biologischen Theorien zufolge müssen die verkürzten Ärmchen dem T. rex irgendwelche Vorteile verschafft haben“, kommentiert das Online-Wissenschaftsmagazin scinexx (Podbregar 2022). Aber welche Funktion hatten diese derart kurzen Arme?
Abb. 1: Rekonstruktion von T. rex (Wikimedia: © Nobu Tamura, CC BY 2.5)
Genau das ist bis heute ein Rätsel, an dem sich die Paläontologen bisher vergeblich die Zähne ausgebissen haben. Alle bislang vorgeschlagenen Antworten sind nicht überzeugend, wie Padian (2022) in einem aktuellen Beitrag erläutert. Dienten die klauenbewehrten Arme dem Festhalten der Beute? Kaum, denn die Schnauze verbiss sich wohl längst tief in der Beute, bevor die kurzem Ärmchen diese überhaupt zu fassen bekamen. Eine Bedeutung bei der Paarung ist ebenfalls ausgeschlossen, da die Ansatzpunkte der beiden Arme viel zu eng zusammen liegen, um den über einen Meter breiten Brustkorb eines Weibchens umfassen zu können. Zudem konnte T. rex seine Arme gar nicht zur Seite bewegen. Auch vorgeschlagene Funktionen in der Kommunikation oder als Signalstruktur sind unwahrscheinlich; auch dafür sind die Arme einfach zu unscheinbar.
Im Rahmen phylogenetischer Hypothesen über Abstammungsverhältnisse muss zudem angenommen werden, dass die Vorderextremitäten im Laufe der Evolution kürzer geworden sind – dies gilt auch bei zwei anderen Dinosaurierlinien, den Abelisauriden und den Carcharodontosauriden (Padian 2022, 63; Abb. 2). „Es ist schwer zu erklären, warum es für die Abstammungslinie von Vorteil sein soll, kleinere Arme zu entwickeln, anstatt größere beizubehalten“, stellt Padian (2022, 67) fest. Für alle vorgeschlagenen Funktionen wären größere Arme deutlich effektiver gewesen (S. 63).

Abb. 2: Carnotaurus (Links) aus der Gruppe der Abelisauriden sowie Tyrannotitan aus der Gruppe der Carcharodontosauriden (Mitte) besaßen ebenso wie Tyrannosaurus (rechts) extrem verkürzte Arme, standen aber zueinander in keinem verwandtschaftlichen Verhältnis. (Wikimedia: Packa & Hic et nunc, CC BY-SA 3.0; Tecnópolis Argentina, CC BY 2.0; Billlion, CC BY-SA 3.0)
Was also kann die kleinen Arme von Tyrannosaurus erklären? Padian sieht eine neue heiße Spur, die mit der Lebensweise zusammenhängt: Es gibt nämlich paläontologische Hinweise darauf, dass mehrere Tyrannosaurier gemeinsam an einer Beute fraßen. Möglicherweise jagten sie auch im Rudel. Wenn sich nun mehrere Tiere gleichzeitig über einen Kadaver hermachten und dabei eng beieinander standen, könnte es bei einem solchen Getümmel leicht passieren, dass versehentlich die Arme eines Artgenossen erwischt wurden. Mit kurzen Armen waren die Tiere vor dieser unter Umständen tödlichen Gefahr geschützt. Die Kürze der Arme an sich hätte demnach keine direkte Funktion, würde aber indirekt vor Schaden bewahren.
Padian sieht Möglichkeiten, diese ausgefallene Hypothese anhand von paläontologischen Befunden zu überprüfen. So sind Bissspuren am Schädel und anderen Teilen des Skeletts von Tyrannosauriern und anderen Raubdinosauriern bekannt. Es wäre zu klären, ob Bissspuren bei T. rex weniger häufig auftreten. Das wäre eine indirekte Bestätigung seiner Hypothese. Padian diskutiert weitere Testmöglichkeiten, die aber insgesamt kein besonders klares Bild ergeben und teilweise auch nicht besonders zielgenau in Bezug auf die vorgeschlagene Hypothese sind. Jedenfalls hält Padian es für wichtig, beim Verständnis des Körperbaus – hier also der kleinen Ärmchen – auch soziale Aspekte, Fressverhalten und andere ökologische Faktoren zu berücksichtigen.
Diskussion
Die vorsichtige Argumentation von Padian und die ausführliche Diskussion von Möglichkeiten, seine Hypothese zu testen, sind durchaus lobenswert. Die Lösung des Rätsels der kurzen Ärmchen des „königlichen Herrschers“ kann einen dennoch zum Schmunzeln bringen. Man stelle sich die Szenerie einer gierig fressenden Tyrannosaurus-Horde vor und wie sie Gefahr laufen, sich dabei gegenseitig schwer zu verletzen. Wenn das kein starker Selektionsdruck ist, die Arme schnellstmöglich zurückzubilden! Dass man dabei wichtige Funktionen der Vorderextremitäten verliert, muss man eben in Kauf nehmen. Aber im Ernst: Ein solches Szenario wirft doch einige schwerwiegende Fragen auf. Der mutmaßliche Nutzen der starken Verkürzung – bzw. das Fehlen eines Schadens – ist ja erst dann gegeben, wenn die Arme schon kurz sind. Wie aber verlief der Weg dahin? Die Rückbildung der Vorderextremitäten wäre hinsichtlich anderer Funktionen zunächst ein Selektionsnachteil; was soll sie also selektiv gefördert haben?
Vielleicht ist beim gegenwärtigen Stand des Wissens die Feststellung angemessener, dass wir Tyrannosaurus einfach nicht verstehen. Es sind vermutlich noch zu viele unbekannte Aspekte im Spiel. Das gilt auch dann, wenn man die Perspektive wechselt und annimmt, der Grundtyp der Tyrannosauriden sei geschaffen worden. Die Frage „Wozu?“ stellt sich dann genauso. Allerdings erübrigen sich in dieser Sichtweise die Fragen nach Selektionsdrücken für die Rückbildung der Arme, weil ein Schöpfer von vorneherein die Gefahr gegenseitiger Verletzung mit einkalkulieren und von Beginn an kurze Arme schaffen kann.
Literatur
Bollinger RR, Barbas RS, Bush EL, Lin SS & Parker W (2007) Biofilms in the large bowel suggest an apparent function of the human vermiform appendix. J. Theor. Biol. 249, 826–831.
Padian K (2022) Why tyrannosaurid forelimbs were so short: An integrative hypothesis. Acta Palaeont. Polon. 67, 63–76.
Podbregar N (2022) Warum hatten Tyrannosaurier so kurze Arme? (4. April 2022) https://www.scinexx.de/news/biowissen/warum-hatten-tyrannosaurier-so-kurze-arme/.
Smith HF et al. (2013) Multiple independent appearances of the cecal appendix in mammalian evolution and an investigation of related ecological and anatomical factors. C. R. Palevol., doi:10.1016/j.crpv.2012.12.001.
Ullrich H (2013) Der Wurmfortsatz: Vom Nichtsnutz zum Mysterium. Stud. Integr. J. 20, 111–115.
Autor dieser News: Reinhard Junker
© 2022, http://www.genesisnet.info/schoepfung_evolution/n301.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.06.21 Ambopteryx: Dinosaurier mit Fledermausflügeln?
Eine Merkmalskombination aus Flughäuten und Federn – gleichsam ein Mix aus Fledermaus und Vogel – hätte man früher bei realen Lebewesen im Rahmen von Evolutionstheorien ausgeschlossen. Die beiden fossilen Arten Yi qi und Ambopteryx longibrachium weisen aber eine solche Kombination auf – allerdings nicht mit echten, flächigen Federn. Beide Arten werden zur Dinosaurier-Gruppe der vermutlich in Bäumen lebenden Scansoriopterygidae („Kletterflügel“) gestellt. Ihre Existenz wirft für evolutionstheoretische Modelle einige schwerwiegende Fragen auf.

Abb. 1 Rekonstruktion von von Ambopteryx longibrachium auf der Grundlage bekannter fossiler Elemente. (Wikimedia: Audrey.m.horn, CC BY-SA 4.0)
Als Kriterium für Wissenschaftlichkeit gilt weithin Falsifizierbarkeit. Das heißt: Es sollten Befunde denkbar sein, die der aufgestellten Hypothese widersprechen und ihre Änderung oder Verwerfung erfordern. Allerdings funktioniert dieses Kriterium bei naturhistorischen Hypothesen kaum und wird in der Praxis auch nicht so gehandhabt (vgl. Junker 2021a). Das gilt auch für Evolutionshypothesen, wie das Beispiel der Hypothesen zur Entstehung des Vogelflugs eindrucksvoll zeigt (Junker 2021b). Es können zudem viele Beispiele von Vorhersagen im Rahmen von Evolutionstheorien genannt werden, die nicht eingetreten sind, ohne dass Evolution als Rahmenparadigma angetastet wurde. Innerhalb dieses Deutungsrahmens gibt es zwar häufig Änderungen im Detail, aber der Rahmen selbst steht nicht zur Disposition.
Zu den von der Standard-Evolutionstheorie (StET) gleichsam verbotenen Befunden gehört laut Beyer (o. J.) das „unabhängige Auftreten spezifischer Ähnlichkeiten. … Daher dürfen mehrteilige und komplizierte Merkmalskomplexe … wie die Vogelfeder, der Wirbeltierknochen oder die Wirbeltierextremität nicht mehrfach und voneinander unabhängig entstanden sein. Falls man derlei findet (z. B. die erwähnte ‚Fledermaus mit Vogelfedern‘), wäre dies ein extrem starkes Falsifikationskriterium für die StET.“
Wenn es so wäre, wären die beiden zur Dinosauriergruppe der baumbewohnenden Scansoriopterygidae („Kletterflügel“) gestellten Gattungen Yi und Ambopteryx ein schwerwiegendes Problem für die Standard-Evolutionstheorie. Im Jahr 2015 war Yi qi („merkwürdiger Flügel“) beschrieben worden (Xu et al. 2015). Yi besaß einen bei anderen Dinosauriern bislang unbekannten langen stabförmigen Knochen, der vom Handgelenk ausging und an dem vermutlich eine Flughaut aufgespannt war, die wahrscheinlich einen Gleitflug ermöglichte. Wie bei anderen Scansoriopterygiden war der dritte Finger verlängert. Darüber hinaus wurden aber auch „Federn“ nachgewiesen – allerdings keine flächigen flugtauglichen Federn, sondern dünne, büschelige, pinselartige Fasern, die kaum eine aerodynamische Bedeutung haben konnten. Die Bezeichnung „Feder“ ist fragwürdig und tendenziös, denn sie ist nicht durch den morphologischen Befund begründet, sondern motiviert durch evolutionstheoretische Hypothesen zur Federentstehung ausgehend von haarartigen Körperbedeckungen bei Dinosauriern. So gesehen könnte also in Bezug auf eine Falsifikation einer Entstehung durch Evolution auch in dieser Hinsicht Entwarnung gegeben werden.
Neuer Fund. Nun wurde eine weitere Gattung mit dieser ungewöhnlichen Merkmalskombination von Flughaut, langem stabförmigem Knochen, langem dritten Finger und haarartigen „Federn“ entdeckt – Ambopteryx longibrachium (Wang et al. 2019), wie Yi in Schichten des Oberjura geborgen (163 Millionen radiometrische Jahre und damit älter als der „Urvogel“ Archaeopteryx). Die „Federn“ sind dicht um den Kopf, den Hals und die Schulterregionen erhalten. Der Gattungsname bedeutet „zweifach geflügelt“. Um den stabförmigen Knochen ist membranartiges Gewebe erkennbar. Die auffallenden Ähnlichkeiten mit Yi rechtfertigen die Zuordnung zu den Scansoriopterygiden. Ambopteryx weist aber auch deutliche Unterschiede zu Yi auf. Beispielsweise ist der leicht gebogene und sich verjüngende stabförmige Knochen anders als bei Yi kürzer als der Oberarmknochen. Die gerade Elle ist kürzer als der Oberarmknochen und hat fast den doppelten Durchmesser im Vergleich zur Speiche, was in starkem Kontrast zum geringen Unterschied zwischen Elle und Speiche bei anderen Scansoriopterygiden steht. Bemerkenswert ist der Besitz einer im Vergleich zu Yi kürzeren Schwanzwirbelsäule mit einem Pygostyl; das sind verschmolzene Schwanzwirbel, an denen bei Vögeln kräftige Muskulatur und fächerförmig angeordnete Schwanzfedern ansetzen. Das Pygostyl ist bei den Scansoriopterygiden als Konvergenz (unabhängig entstandenes Merkmal) zu werten und kann nicht als Hinweis auf eine nähere Verwandtschaft mit den Vögeln interpretiert werden.
Leider erlaubt die Qualität der Erhaltung von Ambopteryx und Yi nicht, die Beschaffenheit des membranösen Gewebes zu rekonstruieren, so dass eine Beurteilung seiner genauen Funktion nicht möglich ist; es ist nach wie vor nicht gesichert, dass die Membran eine Gleitflugfunktion besaß. Diskutiert wird auch eine Unterstützung einer anderen Art der Fortbewegung oder die Funktion als Schmuckelement.
Insgesamt unterscheiden sich die Scansoriopterygiden deutlich von anderen Dinosauriergruppen (Wang et al. 2019, 258). Evolutionstheoretisch müssen sie als spezielle, von anderen Dinosauriern und Vögeln unabhängige Linie interpretiert werden. Anders als die Vögel besaßen die Scansoriopterygiden ungewöhnlich kurze Mittelhandknochen; die Verlängerung der Vorderextremität wird in erster Linie durch die Verlängerung von Oberarmknochen und Elle erreicht, während bei den Vögeln Handwurzel- und Mittelhandknochen zum Carpometacarpus verschmolzen und verlängert sind. Wang et al. (2019, 259) fassen zusammen: „Das gemeinsame Auftreten von kurzen Mittelhandknochen mit membranösen Flügeln – im Gegensatz zu langen Mittelhandknochen und gefiederten Flügeln – zeigt, wie diese beiden deutlich unterschiedlichen Flugapparate die Gesamtstruktur der Vordergliedmaßen beeinflussen können. Membranflügel sind bei den Theropoden der Kreidezeit unbekannt, was darauf hindeutet, dass die Kombination eines vogelähnlichen Körperbaus (z. B. lange Vordergliedmaßen und ein kurzer Schwanz) mit Membranflügeln, die sich bei den Scansoriopterygidae entwickelt hat, eine bisher unbekannte Anpassung von Wirbeltieren mit Flugverhalten darstellt.“
Nach einer neuen Untersuchung der für den Flug relevanten Körperteile von Scansoriopterygiden mittels laserinduzierter Fluoreszenz war ihre Flugmuskulatur schwach, das Brustbein klein und der deltopectorale Kamm am Oberarmknochen, ein Ansatzpunkt für Oberarmmuskeln, nur schwach entwickelt (Dececchi et al. 2020). Daher seien aktiver Flugstart und dauerhafter aktiver Flug praktisch unmöglich gewesen und nur ein suboptimaler Gleitflug denkbar, so Dececchi et al. (2020, 12).
„Experimentierfreudige Evolution“. Eine Merkmalskombination wie bei Yi und Ambopteryx ist evolutionstheoretisch unerwartet. Der „Spiegel“ bringt es populär auf den Punkt: „Ambopteryx longibrachium wirkt wie ein Mix aus Fledermaus, Vogel und Mini-Raubsaurier – als hätte man das Tier aus Teilen verschiedener Puzzles montiert.“1 Nur gibt es in der Evolution keinen Monteur. Die Merkmalskombination passt nicht zwanglos in einen Stammbaum, vielmehr müssen in großem Umfang Konvergenzen, also eine unabhängige Entstehung ähnlicher Merkmale angenommen werden. Das ist eigentlich ein Problem für evolutionäre Hypothesen; nicht umsonst versucht man bei der Konstruktion von Cladogrammen (Ähnlichkeitsbäumen) die Anzahl von Konvergenzen zu minimieren. Aber aus dieser Situation versucht man dennoch, für Evolutionstheorien Kapital zu schlagen: mit der Idee der „Experimentierung“. Die FAZ kommentiert: „An dem Saurier lässt sich ablesen, dass die Natur bei der Erfindung des Fliegens äußerst experimentierfreudig gewesen war und mehrere Lösungen ausprobierte.“2 In der Originalarbeit bei Wang et al. (2019, 256) hört sich das so an: „Die häutigen Flügel, die bei den Scansoriopterygiden durch verlängerte Vordergliedmaßen gestützt werden, stellen wahrscheinlich ein kurzlebiges Experimentieren mit Flugverhalten dar, und gefiederte Flügel wurden schließlich während der späteren Evolution der Paraves bevorzugt.“ Auch Dececchi et al. (2020) sprechen von „Experimentierung“.
Gemessen an den bekannten Evolutionsmechanismen ist das allerdings unhaltbar, weil Mutation, Selektion und andere Evolutionsfaktoren ungerichtet sind, wie gerade die Befürworter der Evolutionslehre immer wieder betonen. Ein Evolutionsprozess wird als natürlicher Vorgang betrachtet, der als solcher keine Ziele verfolgen kann. Warum die Scansoriopterygiden „kurzlebig“ waren und ausgestorben sind, ist in Wirklichkeit unbekannt. Sie hätten jedenfalls nicht entstehen können, wären sie nicht lebens- und konkurrenzfähig gewesen. Man kann davon ausgehen, dass sie ihre eigene ökologische Nische innehatten, die bisher allerdings nicht im Einzelnen aufgeklärt ist. Aus der Sicht der Schöpfungslehre gab es zwar auch keinen Experimentator, jedoch einen Schöpfer, der offenbar eine große Vielfalt an Art und Weisen des Fluges erfunden und verwirklicht hat. Die Frage nach dem Grund des Aussterbens bleibt aber auch hier ohne sichere Antwort.
Anmerkungen
1 https://www.spiegel.de/wissenschaft/natur/forscher-finden-bizarren-fledermausfluegel-saurier-a-1266124.html
2 https://www.faz.net/aktuell/wissen/archaeologie-altertum/flugsaurier-16177567/ambopteryx-longibrachium-im-16176843.html
Quellen
Beyer A (o. J.) Wissenschaft im Rahmen eines Schöpfungsparadigmas? – eine Replik auf ein Positionspapier von „Wort und Wissen“. www.ag-evolutionsbiologie.de/app/download/5785603750/creationscience.pdf
Dececchi TA, Roy A et al. (2020) Aerodynamics show membrane-winged theropods were a poor gliding dead-end. iScience 23(12), 101574, https://doi.org/10.1016/j.isci.2020.101574
Junker R (2021a) Methodologie der Naturgeschichtsforschung. In: Junker R & Widenmeyer M (Hrsg.) Schöpfung ohne Schöpfer? Eine Verteidigung des Design-Arguments in der Biologie. Studium Integrale. Holzgerlingen, S. 65–81.
Junker R (2021b) Erklärungen in der Naturgeschichte am Beispiel der Entstehung von Vogelfeder und Vogelflug. In: Junker R & Widenmeyer M (Hrsg.) Schöpfung ohne Schöpfer? Eine Verteidigung des Design-Arguments in der Biologie. Studium Integrale. Holzgerlingen, S. 83–107.
Wang M, O’Connor JK, Xu X & Zhou Z (2019) A new Jurassic scansoriopterygid and the loss of membranous wings in theropod dinosaurs. Nature 569, 256–259.
Xu X, Zheng X, Sullivan C, Wang X, Xing L, Wang Y, Zhang X, O’Connor JK, Zhang F & Pan Y (2015) A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings. Nature 521, 70-73, doi: 10.1038/nature14423.
Autor dieser News: Reinhard Junker
© 2021, http://www.genesisnet.info/schoepfung_evolution/n292.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
28.04.21 Brutverhalten bei einem Dinosaurier – ein Vogelmerkmal, aber keine evolutionäre Entwicklung
Vogelmerkmale sollen sich unter Dinosauriern schrittweise evolutiv „angesammelt“ haben. Zu diesen Merkmalen soll sich nun auch das Brüten von Gelegen gesellen. Doch für dieses Merkmal muss eine unabhängige Entstehung bei den Dinosauriern angenommen werden; es passt daher nicht als Vorläufermerkmal. Das gilt auch für viele andere Vogelmerkmale bei Dinosauriern.
Dass die Vögel die letzten Überlebenden der Dinosaurier aus der Gruppe der Theropoden („Wildtierfüße“) seien, hat sich als ziemlich fester Bestandteil von Evolutionstheorien etabliert. Darüber hinaus soll der Übergang von Theropoden-Dinosauriern zu Vögeln nachweislich schrittweise erfolgt sein. Die Merkmale, die charakteristisch für Vögel sind, seien im Wesentlichen in unterschiedlicher Häufigkeit bereits bei verschiedenen Dinosaurier-Gattungen aus mehreren Familien der Theropoden ausgebildet gewesen. Man könne daher heute keine Grenze mehr zwischen Dinosauriern und Vögeln ziehen (Brusatte 2017, 55).

Abb. 1: Skizze eines Oviraptor philoceratops. (Wikimedia: Jaime A. Headden (User:Qilong) – http://qilong.wordpress.com/2011/06/05/better-know-an-oviraptorid-the-first-egg-thief/, CC BY 3.0).
Zu den auch bei Dinosauriern vorkommenden Vogelmerkmalen wird seit Kurzem auch das Brüten des Geleges aufgenommen. Zwar waren schon länger Fossilien der Theropodenfamilie der Oviraptoridae („Eierräuber“) bekannt, bei denen fossile Skelette zusammen mit Gelegen gefunden wurden; bei den bisherigen Funden gab es aber Indizien dafür, dass das Elterntier beim Eierlegen oder beim bloßen Bewachen des Geleges überrascht und sedimentiert wurde, ohne dass ein Brutverhalten sicher nachweisbar war (Bi et al. 2020, 1). Nun aber berichtet eine Forschergruppe von einem bisher nicht benannten teilweise erhaltenen Oviraptoriden-Fossil aus der oberen Oberkreide (auf ca. 70 MrJ datiert), bei dem ein erwachsenes Skelett über einem Gelege mit 24 Eiern erhalten ist, von denen mindestens sieben embryonale Überreste enthalten. Die Lage des erwachsenen Tieres in Bezug auf das Gelege, die fortgeschrittenen Wachstumsstadien der Embryonen und die aus den fossilen Befunden (Verhältnisse der Sauerstoffisotope) erschlossenen hohen geschätzten Bebrütungstemperaturen unterstützen klar die Bruthypothese. „Da die Embryonen in späten Entwicklungsstadien starben, hatte das erwachsene Tier das Nest offensichtlich über einen längeren Zeitraum gepflegt und ist nicht beim Akt der Eiablage umgekommen, im Gegensatz zu dem, was bisher für andere Funde angenommen wurde“, schreiben die Autoren (Bi et al. 2020, 6). Bemerkenswert ist darüber hinaus, dass sich die Embryonen im Gelege in unterschiedlichen Entwicklungsstadien befanden, was auf ein asynchrones Schlüpfen hinweist, wie es sonst nur bei „hochentwickelten“ Vögeln bekannt ist.

Abb. 2: Ein Dinosaurier-Gelege. (Wikimedia: Steve Starer; Uploaded by FunkMonk, CC BY 2.0)
Ein weiteres Vogelmerkmal bei einem Dinosaurier-Vorläufer? Die Autoren deuten zunächst ihre Befunde als Unterstützung dafür, dass vogeltypische Merkmale bereits bei Dinosauriern auftreten (Bi et al. 2020, 6f.). Dennoch sei die Entdeckung des asynchronen Schlüpfens bei Oviraptorosauriern unerwartet, da dieses Verhalten selbst unter den Vogelgruppen mit heute lebenden Arten (sog. Kronengruppe), erst spät auftrat. Sie schließen daraus, dass sich dieses Merkmal bei einigen Vögeln der Kronengruppe und zumindest bei den Oviraptorosauriern unabhängig entwickelt habe. Das heißt aber nichts anderes, als dass das Brüten bei den Oviraptoriden gar nicht als Vogelvorläufermerkmal gedeutet werden kann, sondern unabhängig von den Vögeln vorkommt. Dafür spricht auch das relativ geringe geologische Alter; schließlich soll der „Urvogel“ Archaeopteryx mehr als doppelt so alt sein.
Das Indiz der Brutpflege bei Oviraptoriden kann somit nicht als Argument für einen Evolutionsweg von Dinosauriern zu Vögeln gewertet werden, sondern ist im Gegenteil für Evolutionsvorstellungen ein problematischer Befund: Konvergente Entstehungen von nicht-trivialen Ähnlichkeiten sind „unerwartet“.
Bi et al. (2020, 1) drücken das etwas verklausuliert aus, wenn sie in der Zusammenfassung schreiben: „Diese Befunde zeigen, dass die Evolution der Fortpflanzungsbiologie bei den Archosauriern der Vogelgruppe eher ein komplexer als ein linearer und schrittweiser Prozess war, und sie legen nahe, dass einige Aspekte der Fortpflanzung bei den nicht-vogelartigen Theropoden einzigartig für diese Dinosaurier waren.“ Die Autoren schreiben außerdem, dass „die unabhängige Evolution des asynchronen Schlüpfens bei einem nicht-vogelartigen Theropoden“ ein weiteres Beispiel für das „Vorherrschen von Homoplasien bei diesem großen evolutionären Übergang“ sei.
Mit dem Begriff „Homoplasien“ werden Konvergenzen und Parallelismen (unabhängige Entstehung) und Rückentwicklungen zusammengefasst, also Ähnlichkeiten, die nicht als Folge gemeinsamer Abstammung (Homologie) gedeutet werden können. Die Autoren schreiben sogar von einem „Vorherrschen“ der Homoplasien. Tatsächlich ist es der Regelfall, dass vogeltypische Merkmale bei Theropoden-Dinosauriern aufgrund ihrer unsystematischen Verteilung mehrfach unabhängig entstanden sein müssten, wenn sie auf evolutivem Wege entstanden wären. In vielen Fällen sind sie bei mutmaßlichen Vogel-Vorläufern gar nicht nachgewiesen, sondern nur in bestimmten Dinosaurier-Gruppen, die als ganze Gruppe in die Vogelverwandtschaft gestellt werden. Das gilt beispielsweise für verschiedene Federtypen, den Schnabel, das Gabelbein (Furkula), ein großes Brustbein, verknöcherte Brustbeinplatten, ein nach hinten gerichtetes Schambein, ein reduziertes Wadenbein oder für verschmolzene Schwanzwirbel (Junker 2019).
Literatur
Bi S, Amiot R et al. (2020) An oviraptorid preserved atop an embryo-bearing egg clutch sheds light on the reproductive biology of non-avialan theropod dinosaurs. Sci. Bull., https://doi.org/10.1016/j.scib.2020.12.018
Brusatte S (2017) Taking wing. Sci. Am. 316, 48–55.
Junker R (2019) Sind Vögel Dinosaurier? Eine kritische Analyse fossiler Befunde. Internetartikel. https://www.wort-und-wissen.org/wp-content/uploads/b-19-4_dinos-voegel.pdf
Autor dieser News: Reinhard Junker
© 2021, http://www.genesisnet.info/schoepfung_evolution/n291.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
20.01.20 Asfaltovenator vialidadi – ein Dinosaurier so seltsam wie sein Name
Erneut weist eine Fossilform einen unerwarteten Merkmalsmix auf. Asfaltovenator vialidadi wird an die Basis der Dinosaurier-Gruppe der Tetanurae gestellt, besitzt aber auch eine Reihe bunt gemischter abgeleiteter Merkmale, die sonst typisch für verschiedene Gruppen sind. Die Merkmale dieser und anderer Gattungen sind so unsystematisch verteilt, dass extrem viele Konvergenzen bei einer evolutionären Deutung angenommen werden müssen. Als Folge davon können Abstammungsverhältnisse nicht sicher ermittelt werden.
Erst kürzlich berichteten wir an dieser Stelle über einen Fossilfund, der zeitlich, d. h. in der geologischen Schichtenabfolge, im Bereich der Basis einer größeren Gruppe steht, dessen Merkmale aber insgesamt für diese Position nicht passen (s. Parmastega – neuer erster Vierbeiner?, https://www.genesisnet.info/index.php?News=273).
Solche Funde sind nicht selten. Immer wieder stellt sich heraus, dass basale evolutionäre Formen einerseits zwar sogenannte „primitive“ Merkmale besitzen, andere Merkmale jedoch nicht zu dieser Stellung passen. Von einem weiteren solchen Fund – Asfaltovenator vialidadi – berichten der Münchner Paläontologe Oliver Rauhut und der argentinische Paläontologie Diego Pol. Es handelt sich um einen gut erhaltenen Dinosaurier aus mitteljurassischen Schichten, der zur Gruppe der Tetanurae gestellt wird und zu deren geologisch ältesten Fossilfunden gehört. „Tetanurae“ bedeutet „starre Schwänze“, eine größere Gruppe zweibeiniger Dinosaurier, zu denen bekanntere Formen wir Tyrannosaurus rex, Allosaurus oder Velociraptor gehören; aus dieser Gruppe sollen auch die Vögel entstanden sein. Asfaltovenator war etwa 8 m lang; erhalten sind außer dem 80 cm langen Schädel die gesamte Wirbelsäule, vollständige Arme und Teile der Beine.

Abb. 1: Skelettrekonstruktion und Anatomie des Körperskeletts von Asfaltovenator vialidadi. (Wikimedia: Oliver W. M. Rauhut & Diego Pol based on a skeletal reconstruction of Allosaurus by Scott Hartman (Uncredited) – https://www.nature.com/articles/s41598-019-53672-7, CC BY 4.0)
Ungewöhnlich und für die Wissenschaftler überraschend ist die Kombination von Merkmalen, die bisher als apomorph (evolutionär abgeleitet, spezialisiert) für verschiedene Gruppen gehalten wurden. So vereinigt Asfaltovenator Merkmale der Megalosauroidea, der Allosauroidea und verschiedener einzelner Gattungen (Rauhut & Pol 2019, 6). Eine Gattung, die an die Basis einer Gruppe gestellt wird, sollte aus evolutionstheoretischer Sicht jedoch idealerweise nur primitive (ursprüngliche) Merkmale besitzen. Und von diesen beginnend sollten sich in verschiedenen Linien allmählich unterschiedliche abgeleitete Merkmale entwickeln, aus denen die Abstammungsverhältnisse erschlossen werden. Stattdessen findet sich bei Asfaltovenator eine zusammengewürfelte Mischung von abgeleiteten Merkmalen. In Beziehung zur Verteilung der Merkmale bei anderen Gattungen erfordert der Befund die Annahme extrem häufiger Konvergenzen und Parallelismen bei den verschiedenen Dinosauriergruppen (Rauhut & Pol 2019, 7). Das heißt: Viele Merkmale müssten sich mehrfach unabhängig in verschiedenen Linien entwickelt haben. Ihre Verteilung stellt sich ziemlich unsystematisch dar. Folge ist, dass Ähnlichkeiten keine sicheren Indizien hinsichtlich eines gemeinsamen Vorfahren sind. Das liegt daran, dass manche Ähnlichkeiten, die bisher als homolog (abstammungsbedingt) gehalten wurden, nun als konvergent (unabhängig entstanden) eingestuft werden müssen. Die Autoren stellen selber fest, dass diese Situation es schwierig mache, Abstammungszusammenhänge an der Basis größerer Linien zu ermitteln. Verlässliche Rückschlüsse auf die stammesgeschichtlichen Beziehungen der Hauptgruppen und ihrer Untergruppen seien derzeit nicht möglich. Und dies sei auch bei anderen Hauptgruppen der Wirbeltiere verbreitet der Fall.
Rauhut & Pol (2019) vermuten schnelle, fast explosive Evolutionsprozesse infolge eines vorangegangenen Aussterbeereignisses (des sog. „Toarcian Anoxic Event“) als Ursache dieser eigenartigen Merkmalsverteilung. Aufgrund der freigewordenen Lebensräume und damit einhergehender geringerer Selektionsdrücke habe es als Resultat eine experimentelle Phase der Evolution gegeben. Bemerkenswert ist auch, dass die in Abb. 4A ihrer Arbeit gezeigte chronostratigraphische Tabelle nicht nur das explosive Auftreten zeigt, sondern auch, dass Phylogenie (vermutete Abstammungsbeziehungen) und Stratigraphie (fossiles Auftreten in der Schichtenfolge) weithin nicht zusammenpassen.
Kommentar. Weder verursachen unbesetzte Nischen oder geringe Selektionsdrücke evolutive Neuheiten noch gibt es im Rahmen von Evolution „Experimente“. Solche Begriffe und Mutmaßungen kaschieren ein grundsätzliches evolutionstheoretisches Problem: Warum sollten auf der Basis ungerichteter Prozesse vielfach ähnliche Bauplanmodule entstehen, die unsystematisch auf verschiedene Linien verteilt sind? Genau solche Befunde werden evolutionstheoretisch nicht erwartet (deshalb sind sie für die Wissenschaftler überraschend). Es ist keine neue kausale Theorie der Evolution als Lösung in Sicht, die eine solche durch vielfache Konvergenzen geprägte Merkmalsverteilung erklären könnte.
Dagegen könnten solche Befundkonstellationen recht gut zum Konzept polyvalenter Grundtypen im Rahmen eines Schöpfungsmodells passen. Ausgehend von Tiergruppen mit einem großen Potenzial bereits angelegter Merkmalsausprägungen könnten schnelle Radiationen im Rahmen einer Neubesiedlung katastrophisch zerstörter Ökosysteme verständlich gemacht werden. Das wäre dann aber kein evolutiver Prozess im klassischen Sinne, sondern ein Ausschöpfen bereits vorhandener Anlagen, ausgelöst möglicherweise durch Umweltreize. Dafür gib es aus der heutigen Biologie empirisch nachvollziehbare Beispiele (vgl. Crompton 2019).
Literatur
Crompton N (2019) Mendel‘sche Artbildung und die Entstehung der Arten. Internetartikel, http://www.wort-und-wissen.de/artikel/sp/b-19-3_mendel.pdf
Rauhut OWM & Pol D (2019) Probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs. Sci. Rep. 9:18826; doi:10.1038/s41598-019-53672-7
Zitate aus Rauhut & Pol (2019); Hervorhebungen hinzugefügt.
„These results highlight the complex morphological evolution in the early radiation of tetanuran theropods, in which convergences and parallelisms were extremely common. This pattern seems to be a common feature in rapid radiation events of major clades of vertebrates and might explain the common difficulties to unravel phylogenetic relationships of important lineages at the base of major clades.“
„Asfaltovenator furthermore shows characters previously regarded as autapomorphies of different taxa …“
„The abundance and concentration of parallelisms and convergences during the early radiation of the clade explains the difficulties in the establishment of interrelationships of major clades of Tetanurae.“
Autor dieser News: Reinhard Junker
© 2020, http://www.genesisnet.info/schoepfung_evolution/n274.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
22.10.19 Sind Vögel Dinosaurier? Eine kritische Analyse fossiler Befunde
Eine Untergruppe der Dinosaurier, die Theropoden, gilt heute weitgehend unangefochten als Vorläufer der Vögel. Ein Großteil der vogeltypischen Merkmale soll Schritt für Schritt bereits innerhalb der sogenannten „Nicht-Vogel-Dinosaurier“ evolviert sein; es gebe einen durch Fossilien belegten kleinstufigen Weg von Dinosauriern zu Vögeln. Das wird in zahlreichen Fachartikeln behauptet. Eine detaillierte Analyse offenbart jedoch ein deutlich anderes Bild: Für zahlreiche vogeltypische Merkmale muss eine mehrfach konvergente Entstehung angenommen werden und gemäß aktueller Phylogenien waren viele vogeltypische Merkmale gar nicht an der Basis des mutmaßlichen Vogelstammbaums ausgebildet. Viele weitere Befunde zeigen: Der Weg von Dinosauriern zu Vögeln war – evolutionstheoretisch gesehen – alles andere als linear, und zahlreiche Befunde passen besser zu einem Schöpfungsmodell. Reinhard Junker hat dazu ein „Special Paper“ verfasst, dessen Zusammenfassung nachfolgend in modifizierter Form an dieser Stelle publiziert wird.
Vögel gelten heute in der Fachwelt als „lebende Dinosaurier“. In den letzten Jahrzehnten hat sich die mutmaßliche Abstammung der Vögel von Dinosauriern als einer der populärsten evolutionären Übergänge etabliert. In den letzten Jahren wurden zunehmend vogeltypische Merkmale bei Theropoden-Dinosauriern nachgewiesen. Es hat sich herausgestellt, dass ein Großteil von Vogelmerkmalen bei verschiedenen Dinosauriergruppen vertreten ist. Daher gilt ein schrittweiser Übergang von Dinosauriern zu Vögeln als fossil gut belegt. Nur wenige Merkmale eignen sich demnach, um zwischen Vögeln und Dinosauriern zu unterscheiden. Man könne daher heute keine Grenze mehr zwischen Dinosauriern und Vögeln ziehen.
In welchen Theropodengruppen kommen die betreffenden Merkmale vor und welche phylogenetische und welche stratigraphische Position nehmen die betreffenden Gattungen ein? Bewahrheitet es sich, dass die Vogelmerkmale Schritt für Schritt hinzugekommen sind? Untersucht werden folgende Merkmale bzw. Merkmalskomplexe: Federtypen und Flugfähigkeit, Zähne und Schnabel, Gehirn und Encephalisations-Quotient, Furkula, Gastralia (Bauchrippen), Rippenkorb, Brustbein, hohle Knochen, Luftsacksystem und Atmung, Becken und rückwärts orientiertes Schambein, Pygostyl, Wadenbein, Handgelenk sowie Eier, Gelege und Brutpflege.
Ergebnisse. Die Analyse zeigt, dass eine Reihe von vogeltypischen Merkmalen, die bei Theropoden-Dinosauriern vorkommen, mehrfach konvergent entstanden ist und in Phylogenien an der Basis der jeweiligen Gruppen mutmaßlich gar nicht ausgeprägt war. Bei nahezu allen untersuchten Merkmalen müssen aufgrund der mosaikartigen Verteilungen mehr oder weniger häufig Konvergenzen* angenommen werden. Dabei werden teilweise je nach zugrundgelegtem Merkmal verschiedene, sich widersprechende Verwandtschaftsverhältnisse nahegelegt. Dies äußert sich z. B. darin, dass eine Reihe von Gattungen gemessen an etablierten Phylogenien widersprüchliche Merkmalskombinationen aufweist, die eine phylogenetische Einordnung mehr oder weniger stark erschweren. Diese häufig mosaikartige Verteilung begünstigt auch die umstrittene Deutung, dass manche Gattungen, die als Vogelvorläufer interpretiert werden, sekundär flugunfähige Vögel sein könnten. Deren Merkmale würden in diesem Falle als Belege für einen schrittweisen Übergang von Dinosauriern zu Vögeln ausscheiden. Bei einigen Merkmalen sind weiterhin deutliche Sprünge zu verzeichnen.
*Konvergenzen (= mehrfach unabhängige Entstehung)
Aufgrund dieser Befunde ist die Behauptung eines schrittweisen evolutionären „Aufbaus“ des Vogelbauplans anhand des Fossilbefundes stark in Frage gestellt und wird bei einigen Merkmalen durch den Fossilbefund nicht unterstützt.
Darüber hinaus entsprechen diese und andere häufig anzutreffenden Befunde nicht evolutionstheoretischen Erwartungen und können als Anomalien angesehen werden:
- Auf der Basis der Merkmalskonstellationen bei frühen Vögeln und den ihnen nahestehenden Dinosauriern und ihrer stratigraphischen Positionen ergibt sich ein Bild aus unterschiedlichen Mosaiken und folglich eines Netzwerkes von Ähnlichkeitsbeziehungen.
- Es ist nicht klar, welche Gruppe an der Basis der Vögel steht.
- Das große Ausmaß an Konvergenzen hat zur Folge, dass die Cladogramme nicht stabil sind und neue Funde zu größeren Änderungen führen können.
- Die ältesten Vogelgruppen sind schon an der Basis stark differenziert und es gibt teilweise auch „moderne“ Merkmale bei den ältesten Vertretern einer Gruppe.
- Speziell bei Gefiedermerkmalen zeigt sich das größte Ausmaß an Vielfalt nahe der Basis der Vogelgruppen.
- Die meisten Theropoden-Dinosaurier-Gattungen, die vogelartige Merkmale besitzen, sind geologisch jünger als die geologisch ältesten Vögel.
Einige Befunde sind evolutionstheoretisch gut interpretierbar. Manche (aber bei weitem nicht alle) Mosaikformen könnten evolutionären Übergangsformen nahestehen. In einigen Gruppen können im Verlaufe der Kreide Tendenzen der Veränderung von Merkmalsausprägungen nachvollzogen werden. Auch die Tatsache, dass viele Vogelmerkmale bei verschiedenen Dinosaurier-Gruppen der Theropoden vorkommen, kann mit gewissen Einschränkungen (z. B. Konvergenzproblematik) evolutionär gedeutet werden.
„Evolutionäre Experimente.“ Der Mosaikcharakter der Merkmalsverteilungen und besonders die Vielfalt an früh etablierten Federtypen und Arten des Flugs (mit vierflügeligen Formen und teilweise unklaren Flugfähigkeiten) veranlasst viele Forscher, eine Art „evolutionäre Experimentierung“ anzunehmen. Diese Begrifflichkeit ist jedoch evolutionstheoretisch fragwürdig, da sie eine Zielorientierung impliziert, die einem Naturprozess jedoch nicht innewohnt. Die Rede von experimentellen Phasen in der Evolution kaschiert das evolutionstheoretische Problem einer schnell etablierten Vielfalt von unterschiedlichen Mosaiken.
Schöpfung. In den Ursprungsfragen der Biologie werden gewöhnlich nur Erklärungen diskutiert, die dem naturalistischen Paradigma verpflichtet sind. Für diese Festlegung gibt es weder eine naturwissenschaftliche noch eine philosophische Begründung, sondern es handelt sich um eine Konvention, die auch verlassen werden kann. Unter der Annahme einer Erschaffung flexibler, anpassbarer Grundtypen lassen sich die evolutionstheoretisch problematischen Befunde erklären:
- as explosive Auftreten der Formenvielfalt spiegelt die Existenz von Grundtypen wider, die in fertig ausgeprägter und vielfältiger Form erschaffen wurden.
- Die zahlreichen, unterschiedlichsten Mosaikformen sind Ausdruck vielfältiger Kombinationen von Merkmalen, deren Ausprägung durch die jeweilige Lebensweise und nicht durch eine vorausgegangene Evolution erklärt wird.
- Die Schwierigkeiten, die sich für evolutionstheoretische Modellierungen der Entstehungsweise der verschiedenen Mosaikformen ergeben, erübrigen sich, wenn die Merkmale frei und entsprechend der Anforderungen für bestimmte Lebensweisen und Lebensräume kombiniert werden können.
- Die diskutierte Problematik einer „Experimentierung“ entfällt. Es gibt keine „Experimente“, sondern eine anfängliche Vielfalt von Formen, die ursprünglich in mancher Hinsicht am größten war (besonders bei Federtypen und Flugformen).
Der komplette Artikel (93 Seiten, PDF) kann hier heruntergeladen werden:
http://www.wort-und-wissen.de/artikel/sp/b-19-4_dinos-voegel.pdf
Autor dieser News: Reinhard Junker
© 2019, http://www.genesisnet.info/schoepfung_evolution/n271.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
17.05.19 Neues Vogelfossil: Wird der „Urvogel“ Archaeopteryx als „ältester Vogel“ abgelöst?
Ein neu beschriebener Vogel aus dem Oberjura – etwa gleichalt wie der berühmte „Urvogel“ Archaeopteryx – weist einige moderne Merkmale auf. Muss nun eine schnellere Evolution des Vogelfluges angenommen werden?
Erneut macht ein Urvogel-Fund aus den Solnhofener Plattenkalken (Altmühltal, Frankenalb) von sich reden. Nachdem erst Ende letzten Jahres A. albersdoerferi als neue Archaeopteryx-Art beschrieben worden war (Kundrát et al. 2019), wird nun von einer neuen Gattung – Alcmonavis – berichtet, deren fossile Reste in denselben Oberjura-Schichten geborgen wurden, denen auch eines der Archaeopteryx-Fossilien entstammt (Rauhut et al. 2019). Benannt wurde Alcmonavis nach seinem Fundort: Alcmona ist der keltische Name der Altmühl. Der Artname poeschli ist vom Entdecker Roland Pöschli abgeleitet.

Abb. 1: Das Fossil, das als Alcmonavis beschrieben worden ist. (Wikimedia: Oliver Rauhut – https://elifesciences.org/articles/43789, CC BY-SA 4.0)
Gefunden wurde allerdings nur der rechte Flügel, aber er ermöglicht einige interessante Beobachtungen. So sind zwar viele Merkmale ähnlich wie beim „Urvogel“, einige jedoch werden als „moderner“ eingestuft. Die Muskelansatzstellen am Flügel deuten nach Ansicht der Forscher darauf hin, dass Alcmonavis besser für den aktiven Flatterflug gerüstet war als Archaeopteryx. Der Ansatz des Brustmuskels, die Form der Speiche und ein robuster zweiter Finger gelten ebenfalls als „abgeleitete“ Merkmale, die von modernen Vögeln bekannt sind. Alcmonavis teilt somit mit Archaeopteryx den Rang als geologisch ältester Urvogel. Bemerkenswert ist auch, dass Alcmonavis größer war als alle bekannten Exemplare des Archaeopteryx, nämlich gut 10% größer als der größte und mehr als doppelt so groß als der kleinste fossil bekannte Archaeopteryx (Rauhut et al. 2019, 18).
Welche Schlussfolgerungen ziehen die Forscher aus dem neuen Fund? „Die Anpassungen des Urvogels zeigen, dass die Evolution des Fluges relativ schnell vorangeschritten sein muss“, wird Co-Autor Christian Foth auf wissenschaft.de zitiert (Albat 2019). Unter evolutionstheoretischen Vorgaben müsse der aktive Flug früher entstanden sein als bisher angenommen. Außerdem zeige sich, dass die Vielfalt der Vogelwelt im Oberjura größer war als bisher bekannt. Die Forscher werfen auch die Frage auf, ob der neue Fund Hinweise darauf gebe, wie der Vogelflug entstanden sein könnte – direkt oder über ein Gleitflugstadium. Der Fund scheine eher für die direkte Variante zu sprechen, doch es seien zur Klärung weitere Funde erforderlich.
Kommentar. Eine schnelle Entstehung des Vogelflugs ist evolutionstheoretisch nicht zu erwarten. Denn die Erfordernisse für den Flug sind immens (Junker 2016) und der Erwerb der Flugfähigkeit müsste auf der Basis bekannter Evolutionsmechanismen große Zeiträume beanspruchen und ließe viele Zwischenstufen in Bezug auf Flugapparat und Flugfähigkeit erwarten. Die meisten mutmaßlichen Zwischenformen stammen aber aus der geologisch jüngeren Kreide (ausführliche Diskussion zu diesen Formen bei Junker 2017). In der Schichtenfolge tauchen fossile Zeugnisse flugtüchtiger Formen mit dem neuen Fund noch abrupter auf. Einige älter datierte, zur Dinosauriergruppe der Troodontiden gestellte Gattungen aus dem Oberjura wie z. B. Anchiornis waren zwar offenbar voll gefiedert (vgl. Saitta et al. 2017; vgl. Junker 2018), doch deren Merkmalskombinationen erschweren eine Interpretation als Vorfahrenstadien und sprechen eher dafür, dass es sich aus evolutionstheoretischer Sicht um eine unabhängige Linie handelt. Der neue Fund vergrößert weiter die Vielfalt fossiler Formen, die schon bisher nur mit Annahme zahlreicher Konvergenzen in ein Baumschema gebracht werden kann, und eher netzförmige Beziehungen aufweist. Zum Modus des Flugerwerbs kann der neue Fund nichts aussagen, denn mit ihm verkleinern sich die enormen Probleme aller Flugentstehungshypothesen (vgl. Junker 2017, 59ff.) in keiner Weise.
Quellen
Albat D (2019) Gesellschaft für den Archaeopteryx. https://www.wissenschaft.de/erde-klima/gesellschaft-fuer-den-archaeopteryx/
Junker R (2016) Vogelfedern und Vogelflug. 1. Was eine Evolutionshypothese erklären müsste. Stud. Integr. J. 23, 75-82. http://www.wort-und-wissen.de/sij/sij232/sij232-2.html
Junker R (2017) Dino-Federvieh. Zum Ursprung von Vogelfeder und Vogelflug. http://www.wort-und-wissen.de/artikel/sp/b-17-1_feder-und-flug.pdf
Junker R (2018) Neuartiger Federtyp bei mutmaßlichem Dinosaurier. Neuartiger Federtyp bei mutmaßlichem Dinosaurier, https://www.genesisnet.info/index.php?News=257.
Kundrát M, Nudds J, Kear BP, Lüe J & Ahlberg P (2019) The first specimen of Archaeopteryx from the Upper Jurassic Mörnsheim Formation of Germany. Hist. Biol. 31, 3-63
Rauhut OWM, Tischlinger H & Foth C (2019) A non-archaeopterygid avialan theropod from the Late Jurassic of southern Germany. eLife 2019;8:e43789. doi: 10.7554/eLife.43789.
Saitta ET, Gelernter R & Vinther J (2017) Additional information on the primitive contour and wing feathering of paravian dinosaurs. Paleaontology, DOI: 10.1111/pala.12342
Autor dieser News: Reinhard Junker
© 2019, http://www.genesisnet.info/schoepfung_evolution/n267.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
22.02.19 Archaeopteryx albersdoerferi – ein weiterentwickelter Urvogel?
Die genauere Untersuchung des achten Exemplars des berühmten Urvogels Archaeopteryx erwies so viele Unterschiede zu den anderen Funden, dass eine neue Art Archaeopteryx albersdoerferi aufgestellt wurde. Sie erscheint „fortschrittlicher“ als Archaeopteryx lithographica, wird evolutiv jedoch als blinder Seitenzweig gedeutet und nicht als Übergangsform zu anderen Vögeln.
Der berühmte fränkische „Urvogel“ Archaeopteryx lithographica aus Schichten des Oberjura gilt als gut passende Übergangsform zwischen Dinosauriern und Vögeln und hat diesbezüglich geradezu ikonenhaften Charakter. Allerdings sind einige Merkmale z. B. des Schädels oder der Beine zu spezialisiert für eine Übergangsstellung (z. B. Shipman 1998, 116; Hou 2001, 7; Xu & Pol 2013, 331), weshalb Archaeopteryx eher als übergangsformnah oder als modellhaft für eine Übergangsform angesehen wird.

Abb. 1: Das „achte Exemplar von Archaeopteryx“ – beschrieben als neue Art Archaeopteryx albersdoerferi. (Wikimedia: H. Raab (User:Vesta), CC BY-SA 3.0)
Nun wurde das Exemplar 8 von Archaeopteryx, das im Gemeindegebiet von Daiting gefunden worden war, genauer untersucht (Kundrát et al. 2019). Dieses Exemplar wird um 400.000 radiometrische Jahre jünger datiert als die anderen Archaeopteryx-Funde. Und es weist eine Reihe von Merkmalsausprägungen auf, die als fortschrittlicher gewertet werden als die entsprechenden Ausprägungen bei den anderen Funden: Es sind weniger Zähne ausgebildet, die Schädelknochen sind stärker verschmolzen, die Furkula (Gabelbein, mutmaßlich verschmolzene Schlüsselbeine) besitzt eine größere Ansatzstelle für die Flugmuskeln, die Handgelenksknochen sind verstärkt und die Knochen sind mehr pneumatisiert (Ausbildung von Hohlräumen) und dadurch vermutlich leichter. Aufgrund dieser Unterschiede wurde Exemplar 8 nun in eine eigene Art Archaeopteryx albersdoerferi gestellt (nach dem Fossilienhändler Raimund Albersdörfer, der das Exemplar der Öffentlichkeit präsentierte). Die Merkmalsausprägungen von A. albersdoerferi könnten eine verbesserte Flugfähigkeit ermöglicht haben.
Auf den ersten Blick scheint A. albersdoerferi einen evolutiven Wandel in Richtung moderner Vögel zu belegen. Doch auch Kundrát et al. (2019) sehen die Gattung Archaeopteryx auf einem Seitenzweig, der letztlich nicht zu den heutigen Vögeln führte. Die Änderungen von A. albersdoerferi gegenüber Archaeopteryx lithographica seien als Konvergenzen zu werten, also als unabhängige gleichsinnige Änderungen, so die Forscher. Sie sprechen von einer „Mosaikevolution“ und mutmaßen, dass sich wiederholende „Erkundungen“ typisch seien für große Übergänge1 – eine fragwürdige Deutung im Rahmen der Evolutionstheorie, weil sie eine Zielorientierung beinhaltet („Erkundungen“). Werden dagegen evolutionstheoretische Voraussetzungen einer ungerichteten Entwicklung konsequent zugrunde gelegt, sind Konvergenzen grundsätzlich unwahrscheinlich und umso unwahrscheinlicher, je häufiger sie auftreten und je komplexer sie sind. Denn ohne Zielorientierung sind ähnliche neuartige komplexe Konstruktionen, die auf unabhängigen Wegen erreicht werden, nicht zu erwarten und auch nicht durch Selektionsdrücke oder Konstruktionszwänge zu erklären (vgl. Braun 2012). Die Option auf einen Flugerwerb (Selektionsdruck) erklärt schließlich nicht dessen Realisierung.
Befindet sich also Archaeopteryx in evolutionstheoretischer Interpretation auf einem blinden Seitenzweig, würde das auch für A. albersdoerferi gelten.
Es ist allerdings nicht ausgeschlossen, dass es sich bei manchen Archaeopteryx-Funden um Jungtiere handeln könnte. Vielleicht ist auch A. albersdoerferi ein ontogenetisches Stadium (z. B. ein Jugendstadium). Denkbar ist auch innerartliche Variation, die nicht evolutionäre Tendenzen widerspiegelt. Die Gattung Archaeopteryx kann auf der Grundlage ihres gesamten Merkmalskomplexes als variabler Grundtyp2 gewertet werden, der zur gestaltlichen Verschiedenartigkeit der mesozoischen Vögel beigetragen hat.
Literatur
Braun HB (2012) Warten auf einen neuen Einstein. Stud. Integr. J. 12, 12-19.
Hou L (2001) Mesozoic Birds of China. Institute of Vertebrate Paleontology and Paleoanthropology. Phoenix Valley Provincial Aviary of Taiwan.
Kundrát M, Nudds J, Kear BP, Lüe J & Ahlberg P (2019) The first specimen of Archaeopteryx from the Upper Jurassic Mörnsheim Formation of Germany. Hist. Biol. 31, 3-63.
Shipman P (1998) Taking Wing: Archaeopteryx and the evolution of bird flight. New York: Simon and Schuster.
Xu X & Pol D (2013) Archaeopteryx, paravian phylogenetic analyses, and the use of probability-based methods for palaeontological datasets. J. Syst. Paleol. 12, 323–334.
Anmerkungen
1 „These innovations appear to be convergent on those of more crownward avialans, suggesting that Bavarian archaeopterygids independently acquired increasingly bird-like traits over time. Such mosaic evolution and iterative exploration of adaptive space may be typical for major functional transitions like the origin of flight“ (Kundrát et al. 2019, 3).
2 Zu Grundtypen werden alle Biospezies zusammengefasst, die durch Kreuzungen direkt und indirekt miteinander verbunden sind, wobei die Mischlinge nicht fruchtbar sein müssen. So gehören z. B. alle Entenartigen oder Hundeartigen zu einem Grundtyp. Bei nur fossil bekannten Formen kann das Grundtypkriterium zwar nicht angewendet werden, doch geben heutige Grundtypen eine gute Orientierung, welche Vielfalt innerhalb eines Grundtyps ausgeprägt sein kann.
Autor dieser News: Reinhard Junker
© 2019, http://www.genesisnet.info/schoepfung_evolution/n266.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
17.03.18 Neuartiger Federtyp bei mutmaßlichem Dinosaurier
Die zu Dinosauriern gestellte befiederte Gattung Anchiornis („nahe bei Vögeln“) besaß einen bisher unbekannten Federtyp, weder daunenartig noch flächig, der diesem Tier wahrscheinlich ein zotteliges Aussehen verlieh. Die gesamte Merkmalskombination von Anchiornis ist evolutionstheoretisch unerwartet, passt jedoch in ein Schöpfungskonzept. Allerdings ist das Merkmalsmosaik dieses Tieres ohne genauere Kenntnis seiner Lebensweise und seines Lebensraums schwer zu verstehen, so dass Deutungen vorläufig bleiben müssen.
Fossilfunde sogenannter „Nicht-Vogel-Dinosaurier“ und fossiler Vögel waren in den letzten Jahren für zahlreiche Überraschungen gut. Bei einer ganzen Reihe oft nicht näher verwandter Dinosaurier-Arten wurden flaumig-haarige Körperbedeckungen nachgewiesen, die meist als „Protofedern“ bezeichnet werden (was allerdings tendenziös ist, da typische Kennzeichen von Federn fehlen). Eine Reihe von Dinosaurier-Arten besaß dagegen echte, flächige Federn, wobei die Meinungen darüber auseinander gehen, ob es sich dabei bei einigen von ihnen um sekundär flugunfähige Vögel handelt. Überraschend war dabei die Vielfalt von Feder- und Bauplantypen, die das heutige Spektrum deutlich übertrifft. So gab es auch vierflügelige Formen, z. T. wahrscheinlich Doppeldecker-Flieger, die – anders als heutige Vögel – auch an den Beinen kräftig befiedert waren. Und es wurden mehrere speziell ausgeprägte Federtypen nachgewiesen, die bei heutigen Arten ebenfalls nicht bekannt sind. So wurden Konturfedern mit körpernaher bandartiger Ausprägung („proximately ribbon-like pennaceous feathers“) entdeckt, wobei aufgrund der unsystematischen Verteilung von Arten mit diesem Federtyp evolutionstheoretisch eine mindestens vier Mal unabhängige Entstehung angenommen werden muss (Xu et al. 2010). Ein weiterer Federtyp sind verlängerte breite fadenförmige Federn („elongate broad filamentous feathers“; Xu et al. 2009); bei beiden Federtypen wird eine Schaufunktion vermutet. Erst jüngst wurde bei der neu beschriebenen Unterkreide-Gattung Cruralispennia aus der Gruppe der Gegenvögel ein weiterer neuer Federtyp beschrieben (Wang et al. 2017a). Diese Federn waren im körpernahen Bereich drahtartig und besaßen distal fädige Spitzen („proximally wire-like part with a short filamentous distal tip“). Eine Vielfalt von Federtypen war früh etabliert und diese trat recht abrupt auf. Und sie lässt sich nicht ohne Weiteres in ein evolutionstheoretisches Schema einfügen.

Abb. 1: Flügel von Anchiornis. (Wikimedia: ★Kumiko★, CC BY-SA 2.0)
Neuer Federtyp bei Anchiornis. Einen weiteren neuen Federtyp beschreiben nun Saitta et al. (2017) bei der Paraves-Gattung Anchiornis („nahe einem Vogel“), die im Jahr 2009 erstmals beschrieben wurde. Zu den Paraves werden Vögel und deren nach evolutionstheoretischen Vorstellungen nächstverwandte Dinosauriergruppen gerechnet. Anchiornis besaß an Armen, Beinen und am Schwanz kräftige flächige symmetrische Federn; ungewöhnlich daran war ihre Anordnung in mehreren Lagen (Longrich et al. 2012), wofür es kein Vorbild aus der heutigen Vogelwelt gibt. Kürzlich waren Flughäute nachgewiesen worden, die auf eine bessere Flugfähigkeit als zuvor angenommen hinweisen (Wang et al. 2017b). Am ehesten kann Anchiornis demnach als spezialisierter Gleitflieger interpretiert werden. Nun fanden Saitta et al. durch hochauflösende Fotographie heraus, dass neben den mit flächigen Federn bedeckten Bereichen einige Körperpartien mit einem bisher unbekannten Federtyp bedeckt waren, den sie als „zottelig“ („shaggy“) beschreiben: Von einer kurzen Achse gehen beidseitig mehrere lange Äste ab. Dieser Federtyp ist weder typisch daunenartig noch weist er in seiner Form typische Kennzeichen einer Konturfeder auf noch ist er eine intermediäre Ausprägung dieser beiden Federtypen. Diese Art von Befiederung dürfte für ein insgesamt krauses Aussehen gesorgt haben, war mutmaßlich geeignet für Thermoregulation und Wasserabweisung, aber aerodynamisch ungünstig und gegen Nässe nicht gut geeignet (s. Link zu Bild weiter unten). Diese Eigenschaften machen das Verständnis der Lebensweise dieser zunehmend ungewöhnlichen Gattung nicht leichter.
Die Federstrahlen der Flugfedern erwiesen sich als zumindest teilweise vorne offen, was die Flugtauglichkeit stark mindert. Diese Minderung könnte dadurch ausgeglichen worden sein, dass mehrere Lagen von Deckfedern ausgebildet waren. Das allerdings führt zu Gewichtsvergrößerung, was dem Fliegen wiederum abträglich ist. Die Ausstattung des Tieres mit Federn und deren Bau erscheint insgesamt rätselhaft.
Evolutionstheoretische Betrachtungen. Anchiornis besaß ein sehr spezielles Merkmalsmosaik, das insgesamt nicht als Übergangsform zwischen unbefiederten Dinosauriern und Vögeln passt. Weder die Art der Befiederung noch die ungewöhnlichen, „zotteligen“ Federn wurden evolutionstheoretisch vorhergesagt und passen auch nicht in die gängigen Federentstehungsmodelle. Interessanterweise bringen Saitta et al. (2017, 13, 14) wie schon andere Autoren bei anderen „Nicht-Vogel-Dinosauriern“ die Möglichkeit einer sekundären Flugunfähigkeit ins Spiel, also die Abstammung von flugfähigen Formen (Vögeln?).1 Aber manche Merkmale von Anchiornis wie die mehrlagige Federanordnung und der neu entdeckte Federtyp passen gar nicht gut als Zeugen einer Rückbildung. Das Merkmalsmosaik passt aufgrund der speziellen, nicht-intermediären Merkmale auch nicht zu einer Evolution ausgehend von ungefiederten Formen. Dazu kommt, dass ein Gleitflug (wie bei Anchiornis am ehesten anzunehmen) als Vorstufe zum Schlagflug mit vielen Problemen behaftet (Überblick dazu bei Junker 2017, Abschnitt 4.2).
Doch es könnte auch ganz anders sein: Anchiornis mit seinem speziellen Merkmalsmosaik war vermutlich für eine spezielle ökologische Nische und Lebensweise geschaffen. Wenn weder eine „Aufwärts-“ noch eine „Abwärtsevolution“ plausibel sind, ist diese Möglichkeit neben anderen guten Gründen im Spiel. Letztlich scheinen die Kenntnisse über die Lebensweise und den Lebensraum von Anchiornis zu bruchstückhaft und unsicher zu sein, um die komplette Biologie dieses Dinosauriers oder Vogels verstehen zu können, so dass alle Deutungen vorläufig bleiben müssen. Die anatomisch besondere und bisher einzigartige Ausprägung des „zotteligen“ Federtyps sperrt sich gegen eine Ableitung in Form der klassisch-linearen Evolutionsmodelle und kann im Sinne des Schöpfungsansatzes so interpretiert werden, dass es von Anfang an eine Merkmalsvielfalt der Federn gab.
Anmerkung
1 „If a consensus can be built to demonstrate that some non-avian dinosaurs were secondarily flightless, then caution must be had when using their feather morphologies to reconstruct feather macroevolution with regards to the appearance of fully modern feathers and flight. … When examining feather evolution in the future, the impact of secondary flightlessness on feather morphology also needs to be more carefully considered if support for secondary flightlessness in non-avian dinosaurs or basal avialans can be demonstrated based on multiple lines evidence, including quantitative biomechanical evidence“ (Saitta et al. 2017, 14).
Link zu Bild (Neue Rekonstruktion und neuer Federtyp):
http://www.bristol.ac.uk/media-library/sites/news/2017/november/dino-bird-article.jpg
Weitere Artikel über Anchiornis in Genesisnet
- Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137
- Archaeopteryx – Gleitflieger und Bindeglied?, https://www.genesisnet.info/index.php?News=193, auch mit Ausführungen über Anchiornis.
Literatur
Longrich NR, Vinther J, Meng Q, Li Q & Russell AP (2012) Primitive Wing Feather Arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Curr. Biol. 22, 2262-2267.
Junker R (2017): Dino-Federvieh. Zum Ursprung von Vogelfeder und Vogelflug. W+W Special Paper, http://www.wort-und-wissen.de/artikel/sp/b-17-1_feder-und-flug.pdf
Saitta ET, Gelernter R & Vinther J (2017) Additional information on the primitive contour and wing feathering of paravian dinosaurs. Paleaontology, DOI: 10.1111/pala.12342
Wang M, O’Connor JK, Pan Y & Zhou Z (2017a) A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nat. Comm. 8:14141, doi: 10.1038/ncomms14141
Wang X, Pittman M, Zheng X, Kaye TG, Falk AR, Hartman SA & Xu X (2017b) Basal paravian functional anatomy illuminated by high-detail body outline. Nat. Comm. 8:14576, doi: 10.1038/ncomms14576
Xu X, Zheng X & You H (2009) A new feather type in a nonavian theropod and the early evolution of feathers. Proc. Natl. Acad. Sci. 106, 832-834.
Xu X, Zheng X & You H (2010) Exceptional dinosaur fossils show ontogenetic development of early feathers. Nature 464, 1338-1341.
Autor dieser News: Reinhard Junker
© 2018, http://www.genesisnet.info/schoepfung_evolution/n257.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
21.02.18 Caihong – ein Konkurrent für den „Urvogel“?
Erstmals wurde ein gefiederter Dinosaurier entdeckt, der nicht nur älter datiert wird als der „Urvogel“ Archaeopteryx, sondern auch asymmetrische Flugfedern besaß. Damit hat Archaeopteryx den Status als ältestes Fossil auch in Bezug auf dieses Merkmal verloren. Die Merkmalskombination der neu beschriebenen Gattung Caihong ist dennoch evolutionstheoretisch – einmal mehr – unerwartet, füllt keine evolutionäre Lücke und vergrößert die im Oberjura überlieferte Vielfalt gefiederter Formen.
Die Serie interessanter Fossilfunde von gefiederten Formen – seien es Vögel oder Dinosaurier mit Federn – reißt nicht ab. Erneut wurde über einen Fund mit unerwarteten Mosaikmerkmalen berichtet. Bisher war der berühmte Archaeopteryx das stratigraphisch älteste Fossil, bei dem asymmetrische, flugtaugliche Federn nachgewiesen worden waren. Nachdem schon seit einigen Jahren ältere Formen mit symmetrischen Federn bekannt sind (Anchiornis, Eosinopteryx), wurde mit der Gattung Caihong nun erstmals eine Gattung entdeckt, die auch bezüglich der Asymmetrie flächiger Federn den bisherigen Altersrekord überbietet (Hu et al. 2018). Aus evolutionstheoretischer Sicht sollte man unter den Fossilien, die stratigraphisch älter sind als Archaeopteryx, Formen mit einfacheren Federtypen finden. Solche sind auch bekannt: Einfache haar- oder flaumartige Köperanhänge, die als Federn interpretiert werden, sind bei zahlreichen Theropoden-Dinosauriern überliefert – doch diese Gattungen sind fast durchweg jünger als die echt gefiederten Formen aus dem Oberjura und selten etwa gleich alt. Viele Gattungen mit Flaumbesatz sind sogar deutlich jünger (Unter- oder Oberkreide). Daher eignen sich alle diese Formen aufgrund ihrer stratigraphischen Position nicht als Vorstufen von Formen mit echter Fiederung.

Abb. 1: Rekonstruktion von Caihong. (Wikimedia: Lucas-Attwell, CC BY-SA 4.0)
Caihong juji – „Regenbogen mit großem Kamm“ – wurde in den Yanliao-Biota des unteren Oberjura Nordostchinas (Oxfordium, 161 Millionen radiometrische Jahre) gefunden und erhielt seinen Namen wegen mutmaßlich schillernder Federn und auffälligen Kammstrukturen (vermutlich Ornamente) auf den Tränenbeinen (Ossa lacrimales; diese liegen bei Theropoden-Dinosauriern direkt vor und über den Augenhöhlen). Es handelt sich um einen zweibeinigen Räuber mit scharfen Zähnen. Sein Wadenbein ist kurz – eine Besonderheit, die Caihong von verwandten Formen der Paraves (mit den Vögeln nächstverwandte Formen) unterscheidet. Mit ca. 40 cm Länge war das Tier etwa so groß wie eine Ente. Die Vorderextremitäten sind relativ zu den Hinterextremitäten kurz (deutlich kürzer als bei verwandten Formen), während das Längenverhältnis Unter-/Oberarm typisch für heutige Vögel ist (Hu et al. 2018, 2). Der längliche, schmale Schädel weist einige sogenannte autapomorphe (abgeleitete, spezielle) Merkmale auf wie z. B. einige Schädelöffnungen und die erwähnten Knochenkämme. Caihong unterscheidet sich somit insgesamt deutlich von nächstverwandten Formen.
Die Arm- und Fußfedern sowie Deckfedern und Schwanzfedern sind ungewöhnlich lang und länger als beim gleich alten Anchiornis. Die Schwanzfedern haben asymmetrische Federfahnen und bilden eine größere Schwanzfläche als die Federn von Archaeopteryx. Möglicherweise bildeten Federn am Daumen eine Art Alula (Daumenfittich), ein aerodynamisch wichtiges „modernes“ Merkmal. Die recht vielfältige Kombination von Federmerkmalen bei Caihong war bisher nicht bekannt und könnte auf einen besonderen Flugstil hinweisen. Die mögliche Ausbildung einer Alula würde aufgrund der unsystematischen Verteilung dieses Merkmals unter den Paraves eine konvergente (mehrfach unabhängige) Entstehung erfordern (Hu et al. 2018, 8).
In den Federresten wurden als Melanosomen interpretierte, unterschiedlich ausgeprägte Nanostrukturen gefunden. Sie gleichen solchen Strukturen in heutigen Federn, die dunkle und helle schillernde Farben hervorrufen; teilweise erinnern sie an Strukturen, wie sie bei hell schillernden Federn von Kolibris bekannt sind; es fehlen ihnen aber die Luftbläschen, mit denen die Melanosomen der Kolibri-Federn ausgestattet sind.1
Diskussion. Den ungewöhnlichen Merkmalsmix vom Caihong interpretieren Hu et al. (2018) als Folge einer schnellen Merkmalsevolution mit einer erheblichen Vielfalt in Signalgebung und Fortbewegungsstrategien in der Nähe des Ursprungs der Vögel.2 Die Evolution der Federn und besonderer Merkmale der Federn sei schneller verlaufen als die Evolution anatomischer Merkmale. Dabei handelt es sich um eine typische Formulierung zur Deutung eines Befundes, der evolutionstheoretisch unerwartet ist. Denn „schnelle Merkmalsevolution“ entspricht weder evolutionstheoretischen Vorstellungen noch sind dafür plausible Mechanismen bekannt. Der Befund ist: Formen mit unterschiedlichen Merkmalsmosaiken tauchen relativ plötzlich auf; es gibt überraschend viele verschiedene Gefiederausprägungen und anstelle einfacherer Federtypen im Vergleich zu Archaeopteryx mit dessen unbestritten „moderner“ Federform tauchen unter den Formen, die stratigraphisch älter sind als Archaeopteryx, neue Mosaike auf. Caihong füllt keine evolutionstheoretische Lücke, sondern vergrößert die Formenvielfalt der im Oberjura fossil überlieferten gefiederten Gattungen. Die äußert sich in recht großen Unsicherheiten der rekonstruierten Verwandtschaftsverhältnisse und je nach Untersuchung unterschiedlichen Cladogrammen der basalen Paraves.
Hu et al. (2018, 9) äußern die Idee, dass die schillernden Federn knöcherne Ornamente in der Signalgebung abgelöst haben könnten3, doch ist Caihong dafür kein passender Kronzeuge, da diese Gattung beide Optionen in sich vereinigt. Solche Knochenornamente sind sonst bei vielen Dinosauriern bekannt, nicht aber in der näheren Verwandtschaft von Caihong, was den Mosaikcharakter dieser Gattung weiter unterstreicht.
Anmerkungen
1„Although Caihong’s platelet-like structures are most similar in shape to hummingbird melanosomes of the included taxa (…), the interior of hummingbird melanosomes is pitted with air bubbles (…) while the fossilized structures show no evidence of hollowness“ (Hu et al. 2018, 9).
2 The new species indicates the presence of bony ornaments, feather colour and flight-related features consistent with proposed rapid character evolution and significant diversity in signaling and locomotor strategies near bird origins“ (Hu et al. 2018, 1).
3 Integumentary structures may largely replace the signalling role of bony features close to the origin of aerial locomotion“ (Hu et al. 2018, 9).
Literatur
Hu D, Clarke JA, Eliason CM, Qiu R, Li Q, Shawkey MD, Zhao C, D’Alba L, Jiang J & Xu X (2018) A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution. Nat. Comm. 9:217.
Autor dieser News: Reinhard Junker
© 2018, http://www.genesisnet.info/schoepfung_evolution/n256.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
18.01.18 „Dino-Duck“ Halszkaraptor – „rätselhaft und bizarr“
Erneut wurde ein Dinosaurier-Fossil gefunden, dessen Merkmale in kein systematisches Schema passen, sondern einen unerwarteten Mix repräsentieren. Überraschend ist das aber nur, wenn man von einer evolutionären Geschichte des Lebens ausgeht. Aus der Schöpfungsperspektive sind ungewöhnliche Merkmalskombinationen durchaus zu erwarten. Denn ein Schöpfer hat die Freiheit, Mosaikformen hervorzubringen, die nicht von Zwängen hypothetischer Abstammungsprozesse bestimmt werden.
Das Tier ist so seltsam und seine Merkmalskombination so sperrig wie sein Name klingt. Einer der Entdecker, der Paläontologe Andrea Cau (Bologna), beschreibt es als „rätselhaft und bizarr, mit einem völlig unerwarteten Mix aus außergewöhnlichen Merkmalen; die aufregendste Herausforderung für einen Paläontologen“.1 Die Rede ist von der zur Dinosaurier-Familie Dromaeosauridae gestellten Art Halszkaraptor escuillie, die nach der polnischen Paläontologin Halszka Osmólska und dem Fossiliensammler François Escuillié benannt ist; der Namensteil „raptor“ steht für „Räuber“. Das Tier war etwa so groß wie eine Ente und an eine Ente erinnern auch manche seiner Merkmale (s. u.). Da in der „Branche“ Fälschungen nicht ausgeschlossen sind, unterzogen die Forscher das Fossil einer umfassenden 3-D-Analyse und sind aufgrund der Ergebnisse von seiner Echtheit überzeugt. Das Fossil ist komplett in der Gesteinsmatrix eingebettet und vollständig intakt.

Abb. 1: Skelett von Halszkaraptor. (Wikimedia: Ghedoghedo, CC BY-SA 3.0)
Überraschend ist die ungewöhnliche Kombination von Merkmalen. Einerseits besaß das Tier einen langen Hals (50% der Länge von Schnauze bis Kreuzbein) und ans Schwimmen angepasste kurze Vorderextremitäten mit flachen, dünnwandigen Knochen und verlängerten Fingern. Andererseits hatte es lange Beine und war zweibeinig als Läufer unterwegs. Der Kopf war entenartig, aber der Kiefer war mit für einen Dinosaurier ungewöhnlich vielen spitzen Zähnen besetzt, die zum Greifen von Beute geeignet waren. Die Wissenschaftler beschreiben die zweibeinige Körperhaltung als analog der Situation heutiger Vögel.2 Die Forscher schließen auf eine amphibische Lebensweise, wobei der lange Hals zum Gründeln verwendet worden sein dürfte. Dafür spricht auch das Vorhandensein eines gut ausgebildeten Netzwerks von Kanälen (vermutlich mit Nerven und Blutgefäßen) im Zwischenkieferknochen, das dem Tier ein gutes Tastvermögen verliehen haben dürfte. Der Bau der Wirbelkörper lässt auf eine biegsame Wirbelsäule schließen und ist typisch für undulierende (wellenförmig sich fortbewegende) Schwimmer.

Abb. 2: Lebensrekonstruktion von Halszkaraptor. (Wikimedia: Tomopteryx, CC BY-SA 4.0)
Die neue Gattung wird zu den Dromaeosauriden gestellt, einer Gruppe der Maniraptoren („Handräuber“). Zu den Maniraptoren gehören die mit den Vögeln nächstverwandten Dinosaurier-Familien. Nach Cau et al. (2017, 397) handelt es sich bei Halszkaraptor um das erste Beispiel eines Dinosauriers mit „doppeltem Fortbewegungsmodul“; Bau und Ökologie weichen markant von anderen Maniraptoren ab.
In Pressemeldungen ist auch von Federn die Rede und Rekonstruktionen werden entsprechend gezeichnet (s. Link unten), doch sind in der Originalarbeit keine Informationen dazu zu finden. Vermutlich wird aufgrund der Position im Cladogramm auf eine flaumige Körperbedeckung geschlossen, die häufig irreführend als „Federkleid“ bezeichnet wird.
Warum ist ein solcher Merkmalsmix unerwartet? Cau et al. (2017) schreiben, dass die meisten Merkmale von Halszkaraptor bei den Maniraptoren nicht vorkommen, obwohl diese gemäß der Evolutionstheorie den Vögeln nahestehen. Sie teilen jedoch einige Merkmale mit Reptilien und Vögeln mit aquatischer oder semiaquatischer Ökologie, sind also typisch für am oder im Wasser lebende Tiere.3 Ein solches Merkmalsmosaik steht quer zu etablierten evolutionären Abstammungszusammenhängen. Natürlich kann auch Halszkaraptor wie jede andere Art in ein Cladogramm eingefügt werden; sie steht an der Basis der Dromaeosauriden, doch das ist nur bei Annahme vieler Konvergenzen möglich (mehrfache unabhängige Entstehung ähnlicher Merkmale).
Zudem steht das stratigraphische Alter dazu in klarem Widerspruch: Halszkaraptor stammt von der Fossillagerstätte Ukhaa Tolgod in der Wüste Gobi in der Mongolei aus der oberen Oberkreide. Viele Dromaeosauriden, aber auch viele abgeleitete Formen und Dutzende Vogelgattungen der Gegenvögel und der Ornithurae (also deren mutmaßliche Nachfahren) sind bereits aus Gesteinen der unteren Unterkreide überliefert und weisen damit ein deutlich höheres stratigraphisches Alter auf. Der berühmte Archaeopteryx stammt sogar aus noch älteren Schichten des Oberjura und einige eindeutig gefiederte Dino-Gattungen mit unklarer Flugfähigkeit werden noch älter eingestuft. Insgesamt ist verständlich, dass der Merkmalsmix und die stratigraphische Position von Halszkaraptor unerwartet ist, wenn man von einer evolutionären Ableitung der Vögel von den Dinosauriern ausgeht.
Betrachtet man die neue Gattung aus der Perspektive einer Schöpfungsanschauung, kann Folgendes gesagt werden: Ein Schöpfer hat die Freiheit, von ihm geschaffene Merkmale in einer für uns unerwarteten Weise zu kombinieren. Man kann daher durchaus erwarten, dass der theoretisch denkbare Formenraum möglicher Gestalten von Pflanzen und Tieren durch den Schöpfer vielseitig gefüllt werden kann, völlig unabhängig von irgendwelchen Zwängen, die sich aus hypothetischen Abstammungs- und Aufspaltungsprozessen ergeben. Aus dieser Sicht sind weder ungewöhnliche Merkmalsmosaike noch unpassende stratigraphische Positionen überraschend.
Wichtig ist: Halszkaraptor ist kein Einzelfall. Gerade im Formenkreis Dinosaurier – Vögel sind unterschiedlichste Merkmalsmosaike bekannt, was sich in einer geradezu ausufernden Anzahl von Konvergenzen in Cladogrammen bemerkbar macht, worauf viele Kenner des Sachverhalts immer wieder hinweisen; Halszkaraptor ist nur ein besonders markantes Beispiel. (Vergleiche auch Buntes Merkmalsmosaik: Ein „Schnabeltier“ unter den Raubdinosauriern)
Literatur
Cau A, Beyrand V, Voeten DFAE, Fernandez V, Tafforeau P, Stein K, Barsbold R, Tsogtbaatar K, Currie PJ & Godefroit P (2017) Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs. Nature 552, 395-399.
Bilder und Rekonstruktion:
http://www.huffingtonpost.ca/2017/12/07/newly-discovered-dinosaur-halszkaraptor-escuilliei-looks-like-a-mix-of-a-penguin-duck-and-swan_a_23300819/
http://cdn.sci-news.com/images/enlarge4/image_5516e-Halszkaraptor-escuilliei.jpg
Anmerkungen
1 „so enigmatic and bizarre, with a completely unexpected mix of strange features. It was the most exciting challenge for a paleontologist!” (https://news.nationalgeographic.com/2017/12/duck-dinosaur-amphibious-halszkaraptor-fossil-mongolia-science)
2 „… it developed postural adaptations convergent with short-tailed birds” (Cau et al. 2017, 395). „… a body posture that was more erect on land, which is analogous to the postures of modern birds“ (Cau et al. 2017, 397).
3 „… revealed a mosaic of features, most of them absent among non-avian maniraptorans but shared by reptilian and avian groups with aquatic or semiaquatic ecologies“ (Cau et al. 2017, 395).
Autor dieser News: Reinhard Junker
© 2018, http://www.genesisnet.info/schoepfung_evolution/n254.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
04.01.18 Zum Ursprung von Vogelfeder und Vogelflug
Vogelfedern sind die komplexesten Körperanhänge im Tierreich und der Vogelflug gehört zu den faszinierendsten Fähigkeiten von Tieren. Seit Mitte der 1990er-Jahre wurden zahlreiche Dinosaurier-Gattungen mit feder- oder flaumartigen Körperbedeckungen entdeckt. Und es wurden in dieser Zeit neue Modelle der Federentstehung entwickelt. Auf dieser Basis gilt die Entstehung von Vögeln ausgehend von Dinosauriern evolutionstheoretisch als gut begründet. Kürzlich veröffentlichte die Studiengemeinschaft Wort und Wissen ein „Special Paper“ von Reinhard Junker, in dem die evolutionstheoretischen Fragen rund um die Entstehung von Vogelfeder und Vogelflug kritisch diskutiert werden. Wir bringen nachfolgend eine Zusammenfassung dieser Arbeit.
Federn als synorganisierte Gebilde. Vogelfedern sind die komplexesten Körperanhänge im gesamten Tierreich. Für ihre Flugtauglichkeit werden höchste Ansprüche an Material, Bau und Steuerung gestellt, damit Leichtheit, Robustheit, Biegsamkeit und Beweglichkeit gewährleistet sind. Der Feinbau mittels Fasern und Faserbündeln aus beta-Keratin ist äußerst komplex und von der im Follikel befindlichen Spule bis zur Spitze unterschiedlich. Das Bogen-Hakenstrahlen-System ermöglicht kontrolliertes Öffnen und leichtes Schließen. Ein komplexes Geflecht aus Muskeln, Sehnen, Nervenbahnen, Sinneskörperchen und Fettgewebe ermöglicht eine abgestimmte Bewegung und straffe Verankerung der Federn. Zur Gewährleistung der Funktionalität der Federn müssen die verschiedenen Aspekte der Federfeinstruktur und des Federbaus fein aufeinander abgestimmt sein (Synorganisation). Die Federn selbst sind nur ein Teil der notwendigen Voraussetzungen für die Flugfähigkeit der Vögel. Auch zahlreichen anatomischen, physiologischen und verhaltensbiologischen Erfordernissen muss Rechnung getragen werden.
Entstehungsmodelle. Evolutionäre Modelle müssen die Details des Federbaus und des Flugsystems sowie ihrer Steuerung berücksichtigen. Das heute gängige Federentstehungsmodell von Prum (1999) und ähnliche Modelle orientieren sich an den ontogenetischen Abfolgen des Federwachstums. Mit 5-8 Stadien sind sie allerdings viel zu grob, um eine evolutionäre Entstehung realistisch modellieren zu können. Ein Großteil der histologischen und anatomischen Verhältnisse bleibt unbeachtet. Ein Abgleich der Modelle mit den einzelnen Erfordernissen für Federn und Flug erweist die Modelle als untauglich. Es ist daher im Wesentlichen ungeklärt, wie flugtaugliche Federn allein durch zukunftsblinde Variation und Selektion und andere rein natürliche Prozesse entstanden sein könnten.
Fossilien. Seit Mitte der 1990er-Jahre ist mehr als ein Dutzend fossiler Gattungen mit flaumartiger Körperbedeckung (sog. „Dino-Flaum“) gefunden worden, die zu verschiedenen Gruppen von Dinosauriern gehören, darunter zu solchen, die in die Nähe der Vögel gestellt werden (sog. „Nicht-Vogel-Dinosaurier“). Daher werden diese faserigen, z. T. büscheligen oder gering verzweigten Körperanhänge meistens als einfache Federn interpretiert und als Protofedern bezeichnet. Es ist aber nicht erkennbar, ob überhaupt Follikel (im Körper eingesenkter Teil der Federn) erhalten waren, und es ist nicht sicher nachweisbar, ob manche dieser Gebilde hohl waren. Die Bezeichnung „Feder“ ist daher fragwürdig. Aufgrund der sehr weiten systematischen Verteilung von „Dino-Flaum“ wird diskutiert, ob eine faserige bzw. haarartige Körperbedeckung ein allgemeines Kennzeichen der Dinosaurier sein könnte, womit es seinen Indiziencharakter als mögliche Federvorstufe einbüßen würde.
Bei einer Reihe von sog. Nicht-Vogel-Dinosauriern unterschiedlicher systematischer Zugehörigkeit wurden auch flächige Konturfedern nachgewiesen. Diese sind im Wesentlichen heutigen Federn vergleichbar. Es ist nicht auszuschließen, dass es sich dabei um sekundär flugunfähig gewordene Vögel handelt. Im Fossilbericht zeigt sich eine deutliche Kluft zwischen Gattungen mit Dino-Flaum und solchen mit Konturfedern.
Stratigraphische Abfolge. Nicht-Vogel-Dinosaurier mit Flaumbedeckung oder Konturfedern sind nicht nur im cladistischen System recht unsystematisch verteilt, sondern treten in den geologischen Stufen ohne klar erkennbare Ordnung auf. Die geologisch ältesten Gattungen mit Konturfedern sind etwa gleich alt wie die ältesten bekannten Gattungen mit Flaum. Viele Nicht-Vogel-Dinosaurier-Gattungen mit Flaum sind geologisch deutlich jünger als eine Reihe von Gattungen mit flächigen Federn und auch jünger als zahlreiche Vogel-Gattungen der Enantiornithes (Gegenvögel) und Ornithurae („Vogelschwänze“). Gemessen an der stratigraphischen Abfolge ist kein Trend einer Federevolution erkennbar.
Hypothesen zu Vogelflug. Bezüglich der Entstehung des Vogelflugs stehen sich die Cursorialtheorie (evolutiver Beginn des Fluges mit schnellen Läufern vom Boden aus) und die Arborealtheorie (Beginn von Bäumen aus) gegenüber. Die Begründungen für die konkurrierenden Modelle sind ziemlich vage und beruhen vor allem auf Gegenargumenten gegen das jeweils konkurrierende Modell. Die Einwände sind so zahlreich und grundlegend, dass beide Szenarien unglaubhaft sind. Neuere Ansätze wie Flugentstehung auf dem Weg über das „WAIR“-Verhalten („wing-assisted incline running“) oder ein „Sturzflug-Modell“ (Pouncing-Proavis-Modell) scheitern ebenfalls an zahlreichen Einwänden. Wie bei den Federentstehungsmodellen bleiben viele relevante Details unberücksichtigt, weshalb auch die Flugentstehungsmodelle unrealistisch sind.
Selektionsfaktoren. Bis auf wenige Ausnahmen sind die Wissenschaftler der Auffassung, dass Federn nicht von Anfang an auf Flugfähigkeit selektiert wurden, sondern zunächst eine andere Funktion erfüllt haben und später für Flugzwecke kooptiert wurden. Über eine mögliche Erstfunktion (und damit zusammenhängend anfängliche Selektionsdrücke) gibt es verschiedene Auffassungen, ein Konsens ist nicht in Sicht, und Fossilien ermöglichen dazu keine Klärung. Mit dem Umweg über eine andere Erstfunktion von Federn als die der Flugtauglichkeit wird zudem nur sehr wenig gewonnen, weil ein Großteil der für das Fliegen erforderlichen Einrichtungen für alle anderen diskutierten Zwecke nicht benötigt wird.
Methodische Fragen. Die Frage nach der Testbarkeit von Feder- und Flugentstehungstheorien wird in der Fachliteratur nur selten behandelt. Ein direkter (experimenteller) Test ist grundsätzlich nicht möglich, da es sich um historische Prozesse handelt, die anders als im experimentellen Bereich durch möglichst viele bestätigende Befunde (im Vergleich zu konkurrierenden Hypothesen) geprüft werden. Diskutiert werden folgende Kriterien: Änderungsschritte müssen realistisch, kontinuierlich und in der zeitlichen Abfolge passend sein, zu Cladogrammen passen, die funktionelle Ganzheit der Organismen muss berücksichtigt werden. Diese Testkriterien sind jedoch weich, in der Praxis meist vage. Vor allem fallen die vorgeschlagenen Modelle dem Kriterium der Kontinuität und der Berücksichtigung der funktionellen Ganzheit durch.
Schöpfung. Ein grundlegendes Kennzeichen von Wissenschaft ist Wahrheitsorientierung und somit Offenheit auch für grundsätzlich verschiedene Ansätze. Die Möglichkeit einer Schöpfung – einer willentlichen, zielorientierten Hervorbringung durch einen geistbegabten Schöpfer – kann nicht grundsätzlich ausgeschlossen werden. Die Festlegung auf den naturalistischen Ansatz, wonach nur Naturgesetze, Zufall und plausible Randbedingungen in Erklärungen zugelassen, ist wissenschaftstheoretisch nicht zu rechtfertigen und führt zu Zirkelschlüssen. Zahlreiche Befunde im Bau von Federn und in der Fossilüberlieferung sowie das nachhaltige Scheitern naturalistischer Entstehungshypothesen zur Entstehung von Vogelfeder und Vogelflug können als Indizien für einen Schöpfer gewertet werden.
Der Artikel (PDF, 9,2 MB) ist unter http://www.wort-und-wissen.de/artikel/sp/b-17-1_feder-und-flug.pdf frei verfügbar.
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2018, http://www.genesisnet.info/schoepfung_evolution/n253.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.11.17 Serikornis – Dinosaurier mit halbfertigen Federn?
Die kürzlich beschriebene Dinosaurier-Gattung Serikornis besaß sowohl flaumartig-büschelige als auch einfach gefiederte Körperanhänge. Die Deutung als Zwischenform, die zu Vögeln überleitet, ist dennoch problematisch. Das Merkmalsmosaik von Serikornis ist evolutionstheoretisch unerwartet und wesentliche Fragen zur Entstehung flugtauglicher Federn kann das Fossil nicht klären.
Die Serie interessanter Fossilfunde von Vögeln und Dinosauriern, die in die Nähe der Vögel gestellt werden, reißt nicht ab. In den letzten Jahren gab es dabei einige Überraschungen. Statt passender Bindeglieder wurden Formen entdeckt, die unerwartete Merkmalsmosaike aufwiesen wie z. B. Yi qi (mit einer fledermausartigen Flughaut) oder Chilesaurus, oder andere, deren Merkmale oder Fähigkeiten „moderner“ waren als bis dahin angenommen (z. B. bei Anchiornis oder Confuciusornis); siehe Links am Ende des Artikels.

Abb. 1: Rekonstruktion von Serikornis. (Wikimedia: Emily Willoughby (e.deinonychus@gmail.com, http://emilywilloughby.com), CC BY-SA 4.0)
Im Sommer dieses Jahres wurde eine neue fossile Gattung beschrieben, die in einem wichtigen Merkmal evolutionstheoretischen Erwartungen teilweise entspricht, wenn man einen evolutionären Übergang von Dinosauriern zu Vögeln annimmt (was der Auffassung der klaren Mehrheit der Paläontologen entspricht). Die Rede ist von einem nahezu vollständig erhaltenen Exemplar der Art Serikornis sungei aus der Gruppe der sognannten Paraves (Bilder hier: http://novataxa.blogspot.de/2017/08/serikornis.html). Die Bezeichnung „Paraves“ kann man mit „neben den Vögeln“ übersetzen. Diese Gruppe umfasst eine recht bunte Mischung von zweibeinig sich fortbewegenden Gattungen aus verschiedenen Familien, deren Verwandtschaftsbeziehungen oft unklar sind und entsprechend kontrovers diskutiert werden.1 Entdeckt wurde die neue Gattung in der oberjurassischen Tiaojishan-Formation (Liaoning-Provinz, China), in der schon andere Gattungen der Paraves gefunden wurden, darunter mit Anchiornis auch eine vierflügelige Gattung mit eindeutig flächigen Federn; nach neueren Erkenntnissen konnte Anchiornis wahrscheinlich recht gut fliegen (Wang et al. 2017). Serikornis war etwa 60 cm groß und damit so groß wie ein Fasan, vermutlich ein Waldbewohner, der wahrscheinlich auch auf niedriges Geäst klettern konnte (wofür die Existenz stark gebogener Klauen sprechen könnte) (Lefèvre et al. 2017, 11).
Das evolutionstheoretisch interessanteste Merkmal von Serikornis ist der Besitz von unterschiedlich gestalteten flaumigen oder fiederigen Körperanhängen, die den ganzen Körper, auch Beine und Füße bis zu den Fußphalangen bedeckten (Lefèvre et al. 2017). Neben Regionen mit flaumartigen, unverzweigten und dünnen, büscheligen Haaren (evolutionstheoretisch als „Protofedern“ bezeichnet, die man aber nicht unbedingt als „Federn“ bezeichnen muss) sind an den Armen, Beinen, auf dem Rücken und am Ende des Schwanzes auch flächig-fiederige Anhänge mit zentraler Achse (Schaft) überliefert, die man als einfach gebaute Federn interpretieren kann; Serikornis war also sozusagen vierflügelig. Die Vorderflügel waren mit mehreren Lagen relativ kurzer, schlanker, symmetrischer und wenig differenzierter federartiger Anhänge besetzt (eine ähnliche Situation wie bei Anchiornis, bei dem aber wie angemerkt echte Federn ausgebildet waren). Die Autoren erwähnen auch ein großes Propatagium (Flughaut); dessen Existenz scheint aber nur indirekt erschlossen und nicht fossil nachgewiesen zu sein (Lefèvre et al. 2017, 72). Schwung- und Deckfedern – wenn man von „Federn“ sprechen möchte – sind nicht unterscheidbar (anders als beim „Urvogel“ Archaeopteryx, bei der vierflügeligen Gattung Microraptor, bei Confuciusornis und bei modernen Vögeln).
Aufgrund der sehr guten Erhaltung und der erkennbaren feinen Details kann man nach Auffassung der Beschreiber ausschließen, dass Federstrahlen (Bogen- und Hakenstrahlen) ausgebildet waren. Das System aus Federstrahlen ist zwingend notwendig für die Flugfähigkeit; sonst kann der Luft nicht genügend Widerstand entgegengesetzt werden. Vermutlich waren die Federn zudem nicht leicht und nicht steif genug, um ausreichend Schub zu erzeugen.
Darüber, wie die Anhänge im Körper verankert waren, ist offenbar nichts bekannt; die Autoren äußern sich nicht dazu. Damit besteht über ein wichtiges Kennzeichen echter Federn Unklarheit. Ob man also von Federn oder von gefiederten Haaren sprechen will, ist Definitionssache; jedenfalls fehlen wesentliche Merkmale flugtauglicher Federn.3 Die flächigen Federn waren klein, schlank und symmetrisch (ein weiteres Kennzeichen für das Fehlen von Flugtauglichkeit).
Da die Körperanhänge also mit großer Wahrscheinlichkeit nicht für Flugzwecke genutzt werden konnten und auch die Fähigkeit zum Gleitflug wahrscheinlich nicht gegeben war – allenfalls könnten die Federn bei einem Sturzflug bremsend gewirkt haben –, stellt sich die Frage nach ihrer Funktion. Die Forscher diskutieren Isolierung (Thermoregulation) und Zurschaustellung („display“, für die Balz); erst später seien die „Federn“ für den Flug kooptiert worden.
Auch der Skelettbau zeigt keine Anpassung an einen Flug und spricht für ein Leben am Boden. Insbesondere waren die Vorderextremitäten viel zu kurz für eine Flugfunktion. Die für einen Flügelschlag erforderlichen Bewegungen waren aufgrund der geraden Form von Elle und Speiche kaum möglich; die Beinmerkmale sprechen für eine Fortbewegung als Läufer (Lefèvre et al. 2017, 10f.).
Diskussion
Kann Serikornis als Bindeglied zwischen „noch nicht“ flugfähigen befiederten Dinosauriern und Vögeln interpretiert werden? Lefèvre et al (2017, 8) sehen das – wenig überraschend – genau so. Dafür werden zwei Befunde ins Feld geführt: Die einfach gefiederten Anhänge und das stratigraphische Alter (die Position im geologischen System). Die Körperanhänge von Serikornis kommen im Vergleich zu den Körperanhängen von Gattungen mit „Dino-Flaum“ (flaumige, faserige oder büschelige haarartige Anhänge) echten Federn strukturell am nächsten. Und Serikornis gehört zu den geologisch ältesten Gattungen mit „Dino-Flaum“. Diese Punkte gehen an evolutionstheoretische Deutungen. Die Tatsachenaussage der Artikelüberschrift („China documents a transitional step“) ist aber nicht gerechtfertigt, wie die nachfolgenden Aspekte zeigen.
Selektionsdrücke. Serikornis ist nämlich in einem evolutionstheoretischen Rahmen schwer einzuordnen. Die Merkmalskombination aus einfachen Federn, befiederten Beinen, kurzen Vorderextremitäten und Merkmalen, die auf ein Bodenleben hinweisen, ist evolutionstheoretisch unerwartet.4 Denn für einen Bodenstart eines schnellen Läufers sind befiederte Beine nachteilig; für den Baumstart passen die kurzen Vorderextremitäten und das Leben auf dem Waldboden nicht. Problematisch sind die Selektionsdrücke, ohne die evolutionstheoretische Modellierungen kaum auskommen5, und zwar in zweierlei Hinsicht: 1. Welcher Selektionsdruck könnte die Art der Befiederung bei Serikornis begünstigt haben; 2. wie könnte es Richtung Flugfähigkeit weitergegangen sein? Für eine Schaufunktion (Balz) und Thermoregulation erscheinen die fiederigen Anhänge und ihre Art ihrer Anordnung auf der Körperoberfläche nicht notwendig zu sein; sexuelle Selektion als Erklärung ist ein Notbehelf. Selektion auf zukünftige Flugfähigkeit ist nicht möglich, da auf potentiellen zukünftigen Nutzen nicht ausgelesen werden kann. Serikornis ist aus diesen Gründen als Startpunkt für spätere Flugfähigkeit wenig überzeugend.
In einem nicht-evolutionären Deutungsrahmen entfällt die Frage nach Selektionsdrücken, die eine erstmalige Entstehung begünstigt haben könnten. Arten müssen einfach nur überlebensfähig sein, während die sich Frage nach der Überlebensfähigkeit hypothetischer Vor- oder Zwischenformen gar nicht stellt, weil diese nicht benötigt werden. Außerdem müssen nicht alle Merkmale unbedingt unter dem Zweckmäßigkeitsaspekt gesehen werden, es können auch Schönheit oder Phantasiereichtum eine Rolle spielen (hier wird wie oben angedeutet evolutionstheoretisch mit sexueller Selektion argumentiert).
Von Serikornis-„Federn“ zu flugtauglichen Federn. Der Schritt zu flugtauglichen Federn wäre von Serikornis ausgehend immer noch enorm. Die einfache Fiederung bringt – gemessen an den zahlreichen Anforderungen für Flugtauglichkeit – gegenüber einfachen oder büscheligen Haaren kaum etwas. Zur Flugtauglichkeit gehören das ausgefeilte Bogen-/Hakenstrahlen-System, eine geeignete Verankerung im Körper mit der Fähigkeit der Federbewegung (das ist bei Serikornis unbekannt, s. o.), vielfältige Steuerungsmechanismen und vieles mehr (vgl. Junker 2016 und „Wunderwerk Feder“, https://www.genesisnet.info/index.php?News=248).
Vielfalt von Oberjura-Formen. Aus dem Oberjura stammt bekanntlich auch Archaeopteryx, der „moderne“ Federn besaß (dessen Flugfähigkeit aber umstritten ist). Die Juraschichten, in denen er geborgen wurde, sind allerdings stratigraphisch relativ jünger als die Schichten der chinesischen Tiaojishan-Formation; Serikornis ist stratigraphisch älter (s. o.). Aus diesen Schichten und anderen etwa gleich alten Schichten Chinas stammt eine ganze Reihe verschiedener echt befiederter Formen wie z. B. der oben erwähnte Anchiornis, die kletternden Scansoriopterygiden (u. a. der mit einer besonderen Flughaut ausgestattete Yi qi), die Gattung Pedopenna mit langen symmetrischen Konturfedern, die an ganzer Länge am Mittelfuß ansetzen (bei dieser Gattung sind nur die Hinterbeine fossil überliefert (Xu & Zhang 2005) und weitere Gattungen; sie tauchen in ihrer Vielfalt recht plötzlich auf, und ihre Merkmalsmosaike können nicht ohne Weiteres in ein evolutionstheoretisches System gebracht werden. Entsprechend zeigt das von Lefèvre et al. veröffentlichte Cladogramm zahlreiche Konvergenzen bzw. Merkmalswidersprüche und es unterscheidet sich von zuvor publizierten Cladogrammen.
Vermutlich ist über die Lebensweise und die Ökologie von Serikornis wie auch von anderen fossilen Formen, die zu den Paraves gestellt werden, einfach zu wenig bekannt, um sich einen klaren Reim auf die Merkmalskombinationen dieser Gattungen machen zu können. In gewisser Hinsicht erinnert Serikornis an die flugunfähigen Kiwis, die ebenfalls einfach gefiederte Körperanhänge – Federn ohne Federstrahlen – haben, die als rückgebildet interpretiert werden. Es spricht zwar einiges dafür, dass Serikornis auf dem Waldboden lebte, doch die Beinbefiederung erscheint dafür eher hinderlich. Aber auch für ein Klettern sind Federn an den Beinen nicht förderlich. Wie hat Serikornis also gelebt? Hier muss wohl einiges offen bleiben.
Weitere thematisch verwandte Artikel auf Genesisnet
- Buntes Merkmalsmosaik: Ein „Schnabeltier“ unter den Raubdinosauriern, https://www.genesisnet.info/index.php?News=227
- Yi qi – „merkwürdiger Flügel“ eines (Dino-?)sauriers, https://www.genesisnet.info/index.php?News=225
- Mosaikform der Gegenvögel passt erneut nicht ins evolutionäre Bild, https://www.genesisnet.info/index.php?News=247
- Der älteste „echte“ Vogel: überraschend modern, https://www.genesisnet.info/index.php?News=226
- Alte Vögel mit moderner Flugkunst, https://www.genesisnet.info/index.php?News=233
- Urvogel Confuciusornis unerwartet „modern“, https://www.genesisnet.info/index.php?News=244
- Zhenyuanlong – befiederter Dinosaurier, flugunfähiger Vogel oder …?, https://www.genesisnet.info/index.php?News=230
- Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137
Anmerkungen
1 Man vergleiche beispielsweise die Cladogramme in Godefroit et al. (2013a; 2013b), Xu et al. (2011) und Hu et al. (2009).
2 Lefèvre et al. (2017, 7) schreiben: „… forelimb feathers are arranged in several rows in Serikornis, suggesting the presence of a large propatagium, … The development of a large propatagium in basal paravians is therefore supported by the presence of numerous layers of feathers along the forearm in Serikornis …“
3 Chuong et al. (2003) definieren Federn als komplexe Integumentanhänge, die hierarchisch verzweigt sind, aus Rachis, Federästen und Federstrahlen bestehen und aus einem Follikel heraus wachsen und besondere biochemische, morphologische und entwicklungsbiologische Eigenschaften besitzen.
4 Vgl. Pickrell (2017): „The distribution and type of feathers on its body are not consistent with the currently preferred scenario about the evolution of bird feathers and flight. That scenario assumes that long pennaceous feathers on arms and legs originated with arboreal four-winged gliders such as Microraptor.“
5 Evolution ohne Selektion wäre noch problematischer, weil in diesem Fall davon ausgegangen werden müsste, dass sich Voraussetzungen für die Flugfähigkeit latent zufällig entwickelt hätte, was angesichts der Fülle der dafür erforderlichen Voraussetzungen völlig unglaubhaft wäre.
Literatur
Chuong CM, Wu P, Zhang FC, Xu X, Yu M. Widelitz RB, Jiang TX & Hou L (2003) Adaptation to the sky: Defining the feather with integument fossils from Mesozoic China and experimental evidence from molecular laboratories. J. Exp. Zool. 298B, 42-56.
Godefroit P, Cau A, Hu DY, Escuillié F, Wu W & Dyke G (2013a) A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds. Nature 498, 359-362.
Godefroit P, Demuynck H, Dyke G, Hu D, Escuillié F & Claeys P (2013b) Reduced plumage and flight ability of a new Jurassic paravian theropod from China. Nat. Comm. 4:1394,1-6, doi:10.1038/ncomms2389.
Hu D, Hou L, Zhang L & Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 460-463.
Lefèvre U, Cau A, Cincotta A, Hu D, Chinsamy A Escuillié F& Godefroit P (2017) A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers. Sci. Nat. 104:74.
Pickrell J (2017) New Feathered Dinosaur Had Four Wings but Couldn’t Fly. https://news.nationalgeographic.com/2017/08/feathered-dinosaur-four-wings-species-serikornis-science/
Wang X, Pittman M, Zheng X, Kaye TG, Falk AR, Hartman SA & Xu X (2017) Basal paravian functional anatomy illuminated by high-detail body outline. Nat. Comm. 8:14576, doi: 10.1038/ncomms14576.
Xu X, You H, Du K & Han F (2011) An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 475, 465-470.
Xu X & Zhang F (2005) A new maniraptoran dinosaur from China with long feathers on the metatarsus. Naturwissenschaften 92, 173-177.
Autor dieser News: Reinhard Junker
© 2017, http://www.genesisnet.info/schoepfung_evolution/n251.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
22.03.17 Mosaikform der Gegenvögel passt erneut nicht ins evolutionäre Bild
Erneut wurde eine fossile Vogelgattung mit einem ungewöhnlichen Merkmalsmosaik entdeckt. Die zu den Gegenvögeln (Enantiornithes) gestellte Art Cruralispennia multidonta besitzt auch Merkmale, die typisch sind für die andere große Vogelgruppe aus der Kreide, den Ornithurae („Vogelschwänze“). Obwohl Cruralispennia eine abgeleitete Stellung unter den Gegenvögeln einnimmt und nicht als Übergangsform deutbar ist, gehört diese Gattung nach Archaeopteryx zu den ältesten Vögeln – eine „stratigraphisch-phylogenetische Diskrepanz“ (Wang et al. 2017).
Einleitung. In den letzten 30 Jahren wurden ungewöhnlich viele fossile Gattungen von Vögeln in den geologischen Schichten der Unterkreide entdeckt. Nach dem berühmten oberjurassischen Urvogel Archaeopteryx gehören diese Formen zu den stratigraphisch ältesten Gattungen. Die meisten von ihnen lassen sich zwei deutlich unterscheidbaren Gruppen zuordnen, den sogenannten Gegenvögeln (Enantiornithes) und den „Vogelschwänzen“ (Ornithurae). Letztere haben ihren Namen daher, dass sie wie heutige Vögel eine kurze Schwanzwirbelsäule mit einem Pygostyl besaßen (das sind mehrere verschmolzene Schwanzwirbel), an dem ein Fächerschwanz ansetzt. Dagegen besaßen Archaeopteryx und einige andere Gattungen von „Urvögeln“ eine lange Schwanzwirbelsäule und entsprechend einen Fiederschwanz.

Abb. 1: Fossil von Cruralispennia multidonta. Beschriftung in Übersetzung: „Cruralispennia multidonta Holotypus (IVPP V21711). (a) Photographie; (b) Strichzeichnung. ca, Schwanzwirbel; cv, Halswirbel; il, Darmbein; is, Sitzbein; lad, linkes Alularglied; lco, linkes Korakoid; lde, linkes Dentarium; lfe, linker Oberschenkelknochen; lhu, linker Oberarmknochen; lmd, linkes Hauptglied; lpd, linkes Tretglied; lra, linker Radius; lta, linker Tarsometatarsus; lti, linker Tibiotarsus; lul, linke Ulna; pu, Schambein; py, Pygostyle; qu, Quadrate; rco, rechtes Coracoid; rfe, rechtes Femur; rhu, rechter Humerus; rmd, rechter Hauptfinger; rpd, rechter Fußfinger; rra, rechter Radius; rsc, rechtes Schulterblatt; rta, rechter Tarsometatarsus; rti, rechter Tibiotarsus; rul, rechte Ulna; sk, Schädel; st, Sternum; sy, Synsacrum; tv, Brustwirbel. Die weißen Kreise (nummeriert 1-5) und der Kasten zeigen die Lage der Federn bzw. der histologischen Proben an. Maßstabsbalken, 10 mm.“ (Wikimedia: Wang, Minet al. (2017) A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nature Communications. 8: 14141. doi:10.1038/ncomms14141., CC BY 4.0)
Wie so oft zeigt sich auch bei den Vögeln der Unterkreide, dass mit der Zunahme von Funden mögliche Abstammungsverhältnisse nicht klarer werden, sondern immer schwieriger zu rekonstruieren sind. Die Anzahl der evolutionstheoretisch anzunehmenden Konvergenzen von wichtigen Merkmalen nimmt zu, so dass sich Merkmalswidersprüche häufen. Das heißt es muss mehrfache Entstehung weitgehend baugleicher Merkmale angenommen werden – evolutionstheoretisch problematisch.
Der neue Fund. Ein jüngst beschriebener Fund aus der Huajiying-Formation Nordwestchinas (Unterkreide, 131 Millionen radiometrische Jahre), der aufgrund zahlreicher anatomischer Merkmale zu den Gegenvögeln gestellt wurde (Wang et al. 2017, 6), trägt weiter zu diesen Merkmalswidersprüchen bei. Die Huajiying-Formation ist die geologisch älteste Schicht, in der fossile Vögel geborgen wurden, nur übertroffen vom fränkischen Oberjura mit seinen Archaeopteryx-Fossilien. Der neue Fund wurde aufgrund seiner ungewöhnlich befiederten Beine Cruralispennia genannt (crus = Schenkel, pennae = Federn). Außerdem handelt es sich trotz seines geologischen Alters um eine abgeleitete („höherentwickelte“) Form. Das heißt: Sie steht gemäß phylogenetischer Analysen nicht im Bereich der Basis der Gruppe, sondern tief eingeschachtelt weiter oben im Dendrogramm (Ähnlichkeitsbaum). Nach Auffassung der Bearbeiter ist dies „unerwartet“ (Wang et al. 2017, 1), weil evolutionär an der Basis einer Gruppe natürlich mit „primitiven“ Formen gerechnet wird. Die Autoren sprechen von einer „stratigraphisch-phylogenetischen Diskrepanz“ (Wang et al. 2017, 7).
Besonders überraschend ist der Besitz eines pflugförmigen Pygostyls, dessen Form für Ornithurae typisch ist (Gegenvögel haben sonst ein längliches Pygostyl).1 Wang et al. nehmen an, dass es konvergent zu den Ornithurae entstanden ist, was die homoplastische Natur der frühen Vogelevolution aufzeige. (Homoplasie ist ein Sammelbegriff für Konvergenzen, Parallelismus und Rückentwicklungen.) Ebenfalls bisher nicht bekannt unter den Gegenvögeln der Unterkreide war die Ausbildung eines extrem schlanken Coracoids (Rabenbein) bei Cruralispennia; bisher war diese Form des Coracoids nur bei Formen aus der Oberkreide bekannt. Übergangsformen zwischen Formen mit Fiederschwanz und den Pygostylia sind nicht bekannt (Wang et al. 2017, 8).2
Ungewöhnlich sind auch die bereits erwähnten Federn an den Schenkeln, die im körpernahen Bereich drahtartig sind und distal fädige Spitzen besitzen. Diese Federform war bisher unbekannt und erweitert das ohnehin bereits reichhaltige Spektrum von Federformen bei frühen Vögeln, das die Vielfalt heutiger Federtypen noch übersteigt. Wang et al. bezeichnen sie als als „PWFDTs“ – „proximally wire-like part with a short filamentous distal tip“. Ansonsten ist fast das ganze Skelett von verkohlten Federresten umgeben, neben haarartigen Körperfedern sind auch flächige asymmetrische Schwungfedern erhalten.
Auffällig ist schließlich der Besitz von 14 Zähnen auf dem Dentale, was den Artnamen C. multidonta motiviert hat. Fast alle Gegenvögel besitzen weniger Zähne (einzige Ausnahme ist Eopengornis mit 12-14 Zähnen). Interdentalplatten (dreieckige Zwischenzahnwände), wie sie bei Archaeopteryx und theropoden Dinosauriern und z. T. auch bei anderen Sauriern bekannt sind, wurden nicht nachgewiesen. Histologische Befunde an den Knochen deuten darauf hin, dass Cruralispennia schnell wuchs und vermutlich binnen eines Jahres ausgewachsen war; für die Gegenvögel ein weiteres ungewöhnliches, als abgeleitet interpretiertes Merkmal.
Der neue Fund trägt zum Bild eines plötzlichen fossilen Erscheinens einer erheblichen Formenvielfalt in der Unterkreide bei. Auch unter den Ornithurae (ebenfalls Unterkreide) findet sich eine abgeleitete Gattung – Archaeornithura – ausgerechnet unter den geologisch ältesten Formen (vgl. Wang et al. 2015; Der älteste „echte“ Vogel: überraschend modern, https://www.genesisnet.info/index.php?News=226).
Literatur
Wang M (2015) et al. The oldest record of ornithuromorpha from the early cretaceous of China. Nat. Comm. 6:6987, doi: 10.1038/ncomms7987
Wang M, O’Connor JK, Pan Y & Zhou Z (2017) A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nat. Comm. 8:14141, doi: 10.1038/ncomms14141
Anmerkungen
1„Although detailed features are obscured by compression and two-dimensional preservation, the general shape of the pygostyle of Cruralispennia is indistinguishable from that of ornithuromorphs, particularly with regards to the upturned distal end“ (Wang et al. 2017, 8).
2 „Unfortunately, little is known about this dramatic transition, and it is unclear when this feature evolved.“
Autor dieser News: Reinhard Junker
© 2017, http://www.genesisnet.info/schoepfung_evolution/n247.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.03.17 Anfangs explosiv – dann nur noch Fine-Tuning: Vielfalt der Vogelschnäbel
Eine umfangreiche Analyse mittels 3D-Scans von Schnäbeln von etwa 2000 heutigen Vogelarten legt den Schluss nahe, dass am Anfang der fossilen Überlieferung der heutigen Vogelordnungen (Neornithes) sehr rasch eine enorme Vielfalt von Schnabelformen etabliert war. Danach erfolgte im Wesentlichen – von wenigen Ausnahmen abgesehen – eine Art Fine-Tuning. Dieser Befund ermöglicht die Deutung im Rahmen eines Schöpfungsansatzes mit programmierter Variabilität.
Die heute existierenden Vogelordnungen (Neornithes) erscheinen im Fossilbericht plötzlich in großer Vielfalt in den ältesten Schichten des Tertiärs. Wie bei vielen anderen Tier- und Pflanzengruppen ist von einer explosiven Entfaltung oder Radiation die Rede. In den letzten Jahren wurde zwar immer wieder über Funde von Vertretern der Neornithes auch in der Oberkreide berichtet, doch ändern diese nichts daran, dass die Vielfalt der Vogelbaupläne – gemessen an den Fossilfunden – nach der berühmten Kreide-Tertiär-Grenze abrupt und in einem geologisch kurzen Zeitraum zunimmt. Ein gemeinsamer Vorfahr ist nicht bekannt und der Fossilbericht gibt über die Anfänge der Neornithes fast keine Auskunft (Zhou 2004, 456, 467).1 Vergleichend-biologische Studien der heutigen Vogelordnungen bestätigen dieses Bild: Cladogramme (Ähnlichkeitsbäume) lassen sich auf der Ebene der Ordnungen kaum auflösen (vgl. z. B. Thomas 2015). Sie gleichen daher in den unteren Teilen eher einem Busch als einen Baum.
Eine solche vergleichende Studie führen Cooney et al. (2017) anhand von 3D-Scans von Schnäbeln von 2028 heutigen Vogelarten aus 194 Familien durch, die im Natural History Museum Tring (England) und im Manchester Museum archiviert sind und einen Großteil des Systems der Vögel abdecken. Das charakteristische Merkmal des Vogelschnabels ist für seine faszinierende Vielfalt von Gestaltungen bekannt. Die unterschiedlichen Schnabelformen ermöglichen den Vögeln, sehr verschiedene Nahrungsquellen zu nutzen. Durch die Kombination der Ergebnisse der Untersuchungen der Schnabelformen mit neueren DNA-basierten Cladogrammen ergab sich, dass die Vielfalt der Schnäbel sehr schnell in der mutmaßlichen evolutionären Geschichte aufgetreten ist, also innerhalb kurzer Zeit stark zunahm. Evolutionstheoretisch folgt daraus, dass ein extrem schneller evolutiver Wandel angenommen werden muss – die Autoren sprechen von „früher schneller Quanten-Evolution“ (Cooney et al. 2017, 345), das bedeutet eine anfänglich sehr schnelle Divergenz (Aufspaltung und Verschiedenwerden) in neue Formen und Funktionen. Von wenigen Ausnahmen abgesehen änderten sich die Schnabelformen nach der Anfangsdivergenz nur noch relativ geringfügig in der Art eines Fine-Tunings und durch ökologische Anpassungen (vgl. Bhullar 2017).
Fossilfunde wurden in die Untersuchung nicht einbezogen. Bhullar (2017) weist aber darauf hin, dass der Fossilbericht den Befund von Cooney et al. unterstützt, dass eine große Vielfalt von Schnabelformen nach der Kreide-Tertiär-Grenze rasch präsent war, ja dass es anfangs noch weitere Schnabelformen gab, die heute nicht mehr vertreten sind. Die Gründe für dieses Muster mutmaßlicher evolutionärer Sprünge und anschließender Stabilität seien nicht bekannt.
Mikroevolution, Makroevolution und Schöpfung. Die Autoren stellen fest, dass es einen qualitativen Unterschied zwischen Mikroevolution (z. B. Fine-Tuning) und Makroevolution (neuartige Baupläne oder Bauplan-Elemente) gibt. Sie schreiben: „Auf welche Weise mikroevolutive Prozesse addiert werden, um die Ausweitung der gestaltlichen Vielfalt über viel größere evolutionäre Zeiträume hinweg zu formen, ist unklar“ (Cooney et al. 2017, 344). Und weiter: „Untersuchungen an kleineren (evolutiven) Zweigen haben Einsichten in die Rolle der natürlichen Auslese als verändernde Kraft ermöglicht, sie können aber nicht den Prozess erhellen, der die Vielfalt und Diskontinuitäten von Radiationen über längere evolutionäre Zeiträume formen“ (ebd.).2 Das entspricht vielen ähnlich lautenden Einschätzungen zum Unterschied von Mikro- und Makroevolution (vgl. Entstehung evolutionärer Neuheiten – ungelöst!, https://www.genesisnet.info/index.php?News=243).
Das Muster der Entfaltung der Schnabelformen kann bei Zugrundelegung eines Schöpfungsansatzes mit programmierter Variabilität gut erklärt und eingeordnet werden. Dass innerhalb einer Formengruppe eine große Vielfalt relativ plötzlich auftaucht und Cladogramme an der Basis kaum auflösbar sind bzw. anfangs sehr enge Verzweigungen beinhalten, könnte als Indiz dafür gewertet werden, dass es gar keine Abstammung der größeren Einheiten gab, sondern dass diese in fertiger Form durch Schöpfungsakte ins Dasein kamen und sich anschließend ein anfangs bereits angelegtes Potential an Ausprägungs- und Spezialisierungsmöglichkeiten entfaltet hat.
Für die biblische Schöpfungslehre bleibt jedoch die Frage offen, warum zum einen Vögel heutiger Grundtypen fossil weitgehend erst ab dem unteren Tertiär überliefert sind, aber auch, weshalb Vögel, die im älteren geologischen System der Kreide fossil überliefert sind, andere Baupläne besitzen als die tertiären und heute lebenden Vögel.
Anmerkungen
1 „[R]elatively little is known about the origin and the immediate ancestor of all extant avian groups“ (Zhou 2004, 456). „[Y]et there is still no fossil evidence indicating the origin of modern groups in the Mesozoic“ (Zhou 2004, 467).
2 „[B]ut how microevolutionary processes scale up to shape the expansion of phenotypic diversity over much longer evolutionary timescales is unclear“ (Cooney et al. 2017, 344).
„Studies of small clades have provided insights into the role of natural selection as a diversifying force, but cannot illuminate the processes that shape the diversity and discontinuities of radiations over longer evolutionary time frames“ (ebd.).
Literatur
Bhullar BAS (2017) Catastrophe triggers diversification. Nature 542, 304-305.
Cooney CR, Bright JA et al. (2017) Mega-evolutionary dynamics of the adaptive radiation of birds. Nature 542, 344-347.
Zhou Z (2004) The origin and early evolution of birds: discoveries, disputes and perspectives from the fossil record. Naturwissenschaften 91, 455-471.
Thomas GH (2015) An avian explosion. Nature 526, 516-517.
Autor dieser News: Reinhard Junker
© 2017, http://www.genesisnet.info/schoepfung_evolution/n245.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.02.17 Urvogel Confuciusornis unerwartet „modern“
Der „Konfuzius-Vogel“ – Confuciusornis sanctus – gehört aufgrund zahlreicher fossiler Funde zu den am besten bekannten Vögeln der Unterkreide und nach Archaeopteryx auch zu den stratigraphisch ältesten. Neue Untersuchungstechniken brachten neue Einsichten in die Weichteilanatomie des Flugapparats. Confuciusornis war demnach ein besserer Flieger als bisher gedacht – angesichts der zahlreichen Anforderungen an aktiven Flug ist das letztlich nicht überraschend.

Abb. 1: Confuciusornis sanctus, Naturhistorisches Museum Wien. (Wikimedia: Tommy from Arad, Uploaded by FunkMonk, CC BY 2.0)
Die Vogelgattung Confuciusornis gehört nach dem berühmten oberjurassichen „Urvogel“ Archaeopteryx zu den stratigraphisch* ältesten Vögeln. Im Jahr 1995 wurden gut erhaltene fossile Exemplare dieser Gattung in großer Zahl in Süßwasserablagerungen der unteren Kreide im Nordosten Chinas gefunden. Während manche Merkmale (z. B. Becken, Finger, Schädel mit zwei Schläfenfenstern) als „ursprünglich“ eingestuft werden, besaß Confuciusornis – anders als Archaeopteryx und viele andere „Urvögel“ – einen Hornschnabel und eine Reihe weiterer „fortschrittlicher“ Merkmale. So hatte er keinen langen, reptilienähnlichen Schwanz, sondern wie heutige Vögel ein Pygostyl (mehrere verschmolzene Schwanzwirbel), an dem die Schwanzfedern fächerförmig ansitzen. Bei einigen Exemplaren ist auf dem Brustbein ein niedriger Kiel in der hinteren Hälfte zu erkennen; das Rabenbein ist ähnlich wie bei heutigen Vögeln lang und strebenförmig. Wie bei heutigen Vögeln dürften Ellbogen- und Handwurzelgelenk mechanisch gekoppelt gewesen sein, was ebenso wie das gut ausgebildete Federkleid mit asymmetrischen Flugfedern für eine Fähigkeit zu aktivem Flug spricht.
*stratigraphisch (= die geologische Schichtenfolge betreffend)
Die Lebensweise und die Art der Flugfähigkeit von Confuciusornis wurden dennoch kontrovers diskutiert (Wang et al. 2011, 1229). Peters & Ji (1998) hielten den Konfuzius-Vogel wegen der großen Schwingen für einen ausdauernden Segler; ob Confuciusornis klettern konnte, war bisher ebenfalls umstritten (z. B. Feduccia 2001, 143; Chiappe et al. 1999, 1). Nudds & Dyke (2010) hatten eine Studie veröffentlicht, wonach die Schäfte der Federn von Confuciusornis zu schwach für eine Tauglichkeit zu aktivem Flug gewesen seien, doch ihre Schlussfolgerungen wurden wegen methodischer Fehler zurückgewiesen (Paul 2010; Longrich et al. 2012; Lingham-Soliar 2015).
Neue Einsichten zu diesen Fragen und weitere Erkenntnisse über die Flug- und Kletterfähigkeit brachten nun Untersuchungen mit Laser-unterstützter Fluoreszenzmikroskopie (Falk et al. 2016). Damit konnten bisher unbekannte Details der Weichteilanatomie (Bänder, Muskeln, Sehnen) gewonnen werden. Sie zeigen, dass Confuciusornis „moderner“ gebaut war als bisher angenommen. Die Autoren stellen fest, dass Confuciusornis „eine Reihe relativ moderner Weichgewebe-Strukturen besaß, die fortschrittlicher waren als erwartet werden konnte“ (Falk et al. 2016, 6).1 Außerdem konnten gut entwickelte und widerstandsfähige Flughäute (Pro- und Postpatagium an Innen- und Außenseite der Flügel) nachgewiesen werden, was laut Falk et al. enormen Auftrieb ermöglicht haben müsse und zusammen mit den robusten Federschäften für die Fähigkeit zum aktiven Flug spreche. Die Autoren schließen unter evolutionstheoretischen Voraussetzungen, dass das Flughautsystem früh entstand und ein gemeinsames Merkmal (Synapomorphie) aller Vögel sein könnte (Falk et al. 2016, 8).2
Auch zur Flügelform konnten neue Erkenntnisse gewonnen werden; diese sei einzigartig gewesen, folgern Falk et al. aus dem erhaltenen Federkleid. Die Breite des Flügels unterstreiche einen anderen Flugstil als bisher angenommen und sei typisch für Vögel, die in einem dicht bewachsenen Lebensraum vorkommen und daher ein hohes Maß an Manövrierbarkeit benötigen, oder auch für Segler. Sie schließen aus der Flügelstreckung (aspect ratio), dass Confuciusornis in seinem Lebensraum eher Manövrierbarkeit als Geschwindigkeit benötigte.3
Netzförmige Schuppen auf den Beinen, kräftige Fingerpolster und stark gebogene Klauen an den Zehen unterstützen nach Falk et al. nachhaltig die Rekonstruktion als baumlebender Vogel. Die netzförmigen Schuppen sind mehr abgerundet als schildförmig, was die Flexibilität der Zehen verbessert.
Aus den neuen Befunden schließen die Forscher insgesamt, dass Confuciusornis Kurzstrecken gut fliegen konnte und dass dieser Urvogel viele relativ „fortschrittliche“ („advanced“) vogeltypische anatomische Merkmale besaß. Insgesamt war der Konfuzius-Vogel also keinesfalls primitiv, weshalb die Autoren evolutionstheoretisch mutmaßen, dass seine modernen Merkmale viel früher als bisher angenommen entstanden sein müssten; ältere Gesteinsschichten könnten vielleicht entsprechende Antworten über Vorstufen bringen.4
Erst kürzlich waren bei einem anderen Vogel aus Schichten der Unterkreide ebenfalls Weichteilmerkmale identifiziert worden, die als überraschend modern charakterisiert wurden und für eine sehr gute Flugfähigkeit sprechen (Navalón et al. 2015), so dass auch in diesem Fall eine Revision bezüglich der Flugfähigkeit vorgenommen wurde: Die Gegenvögel, zu welchen der Fund gerechnet wird, werden nun als gute Flieger angesehen. (Vgl. Alte Vögel mit moderner Flugkunst, https://www.genesisnet.info/index.php?News=233)
Diese Befunde und Interpretationen sind letztlich aber nicht überraschend. Der Besitz asymmetrischer Federn wie bei Confuciusornis und anderen Vögeln aus Kreideschichten macht nur Sinn, wenn die betreffenden Arten flugfähig wären; dafür aber benötigen sie zugleich auch eine ausgefeilte passende Weichteilanatomie. Flugfähigkeit erfordert einen Komplex an Merkmalen und Fähigkeiten – vom geeigneten „Baummaterial“ für die Federn bis zum Verhalten mit den dafür erforderlichen Gehirnleistungen und Rückkopplungsmechanismen. Eine schrittweise Entstehung einer Grundausstattung ist hier kaum denkbar. Die neuen Daten zu Confuciusornis bestätigen diese Sicht.
Literatur
Chiappe LM, Ji SA, Ji Q & Norell MA (1999) Anatomy and systematics of the Confuciusornithidae (Theropoda, Aves) from the late Mesozoic of northeastern China. Bull. Amer. Mus. Nat. Hist. 242, 1-89.
Falk AR, Kaye TG, Zhou Z & Burnham DA (2016) Laser fluorescence illuminates the soft tissue and life habits of the Early Cretaceous bird Confuciusornis. PLoS ONE 11(12): e0167284. doi:10.1371/journal.pone.0167284
Feduccia A (2001) The problem of bird origins and early avian evolution. J. Ornithol. 142 Sonderheft 1, 139-147.
Hou, L, Zhou Z, Gu Y & Zhang H (1995a) Confusiusornis sanctus, a new late Jurassic sauriurine bird from China. Chinese Sci. Bull. 40, 1545-1551.
Hou L, Zhou Z, Martin, LD & Feduccia A (1995b) A beaked bird from the Jurassic of China. Nature 377, 616-618.
Lingham-Soliar T (2014) Feather structure, biomechanics and biomimetics: the incredible lightness of being. J. Ornithol. 155, 323-336.
Longrich NR, Vinther J, Meng Q, Li Q & Russell AP (2012) Primitive wing feather arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Curr. Biol. 22, 2262-2267.
Navalón G, Marugán-Lobón J, Chiappe LM, Sanz JL & Buscalioni A (2015) Soft-tissue and dermal arrangement in the wing of an Early Cretaceous bird: Implications for the evolution of avian flight. Sci. Rep. 5:14864.
Paul GS (2010) Comment on „Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability“. Science 330, 320b.
Peters D & Ji Q (1998) The diapsid temporal construction of the Chinese fossil bird Confuciusornis. Senckenbergiana lethaea 78, 153-155.
Wang X, Nudds RL & Dyke GJ (2011) The primary feather lengths of early birds with respect to avian wing shape evolution. J . Evol. Biol. 24, 1226-1231
Originalzitate
1 „This new information indicates that Confuciusornis has a suite of relatively modern soft tissue structures that are more advanced than may be expected.“
2 „This suggests that the patagial system in birds developed early in their evolution and, when available for study, may be a uniting synapomorphy. … The extensive and robust propatagium in Confuciusornis would have generated a large amount of lift.“
3 The wider, broader wing in Confuciusornis confirms a flight style different than previously suggested (see [6]) (Fig 6). Wide, broad wings are found in 1) birds that live in a densely vegetated area requiring high maneuverability and 2) broad-winged soaring birds such as hawks and vultures. … implies Confuciusornis required more maneuverability and stability than speed in flight“ (FALK et al. 2016, 6). „The aspect ratio of Confuciusornis is between 6.4 (including tertial feathers) and 7.7 (excluding tertial feathers), indicates that it was not a dynamic soarer (e.g., seabirds), long-distance migrant (e.g., shorebirds), or a long-duration fast flyer (FALK et al. 2016, 6f.).
4 „If Confuciusornis, a primitive bird quite basal on the avian tree (e.g. see [39]), possessed a suite of characters so modern, it suggests that these features arose much earlier than perhaps previously implied. If that is so, then earlier rocks may contain the answers to many questions about the origin and early evolution of birds“ (FALK et al. 2016, 13).
Autor dieser News: Reinhard Junker
© 2017, http://www.genesisnet.info/schoepfung_evolution/n244.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
13.12.16 Dino-Federn in Bernstein?
Ein neuer Bernsteinfund sorgt für Furore: Erstmals wurden federähnliche Strukturen zusammen mit Knochenstrukturen in Bernstein entdeckt. Die Zuordnung des eingeschlossenen Schwanzwirbelsäulenfragments zu einem Dinosaurier aus der Gruppe der Coelurosaurier erscheint allerdings nicht gesichert. Der Federaufbau entspricht keiner bekannten Federstruktur und passt nicht gut zum derzeit favorisierten Federentstehungsmodell. Mit 99 Millionen radiometrischen Jahren (mittlere Kreide) ist der Fund viel zu jung, um Auskunft über eine hypothetische Evolution von Federn zu geben, da bereits der berühmte „Urvogel“ Archaeopteryx und andere Gattungen aus dem Oberjura und der Unterkreide moderne Federn besaßen.
Bernstein-Einschlüsse (Inklusen) bieten faszinierende Einblicke in die frühere Lebewelt. Ungezählte Insekten, Spinnentiere, Pflanzenteile, Mikroorganismen und in selteneren Fällen sogar Fröschchen, kleine Echsen oder Federn sind dadurch erhalten. Nun wurde erstmals ein kurzer Abschnitt (ca. 36 mm) eines (Dinosaurier-?)Schwanzes, an dem sich Wirbel abzeichnen, in einem Bernstein beschrieben (Xing et al. 2016a); das Stück wurde auf einem Bernsteinmarkt in Myanmar erworben und stammt aus einer der dortigen Bernsteinminen. Das Besondere an dieser Inkluse: Der Schwanz ist nicht nur von Wirbeln durchzogen; an seiner Oberfläche ist auch ein Besatz von federähnlichen Anhängen erhalten. Damit wurden erstmals federähnliche Strukturen zusammen mit Knochen in Bernstein entdeckt. Außerdem bietet der Fund Einblicke in die räumliche Anordnung der fiederigen Strukturen. Auch in dieser Hinsicht ist der detailreiche Fund aus Schichten der mittleren Kreide Myanmars, die radiometrisch auf 99 Millionen Jahre datiert werden, interessant.
Bei den zahlreichen bisher beschriebenen Dinosaurierfosssilien mit flaumartiger Körperbedeckung sind wegen schlechter Erhaltung die Details der Körperanhänge oft nicht genau rekonstruierbar, außerdem handelt es sich dabei um Kompressionsfossilien, d. h. deren Überreste sind zusammengedrückt und quasi nur 2-dimenional erhalten.
Aufgrund von wenigen und wenig spezifischen Merkmalen der erkennbaren Wirbel wagen die Autoren eine Zuordnung des Fossils zu den Coelurosauriern („Hohlschwanzechsen“); die abgeleitete geringe Größe interpretieren sie als Hinweis ein Jugendstadium. Xing et al. vermuten, dass das Tier spatzengroß war; wie groß es in ausgewachsenem Zustand war, ist schwer abzuschätzen. Coelurosaurier sind eine größere Gruppe unter den Dinosauriern; sie werden in zahlreiche Untergruppen unterteilt, zu denen auch die Deinonychosauriden gehören, die als nächstverwandt mit den Vögeln gelten (sie werden von den meisten Forschern als Schwestergruppe der Vögel angesehen). Auch die Tyrannosauriden gehören zu den Coelurosaurier; diese Gruppe ist also sehr vielseitig. Eine präzisere Zuordnung zu einer Familie innerhalb der Coelurosaurier ist nicht möglich.
Aus Größe und Form der Wirbel des erhaltenen Bruchstücks schließen die Autoren, dass das Tier mindestens 15, wahrscheinlich über 25 Wirbel besaß. Zum Vergleich: Der berühmte „Urvogel“ Archaeopteryx besaß 23 Schwanzwirbel, der Urvogel Jeholornis bis zu 27 (Zhou & Zhang 2003); deren Schwänze waren daher als Fiederschwänze ausgebildet. Die Autoren halten es aufgrund morphologischer Details der Wirbel allerdings für unwahrscheinlich, dass der Fund zu solchen Fiederschwanz-Vögeln gestellt werden könnte. (Heutige Vögel besitzen einen Fächerschwanz, der an einem Pygostyl – einige verwachsene Schwanzwirbel – ansetzt; das trifft auch auf die meisten fossilen Vögel aus der Kreide zu.)
Das Schwanzfragment ist an beiden Seiten dicht mit Reihen von wenige Millimeter langen Federn besetzt (http://www.cell.com/action/showImagesData?pii=S0960-9822%2816%2931193-9; Figure 1). Sporadisch sind Weichgewebe, vermutlich Muskeln, Bänder und Haut erkennbar.
Die Federn wurden mikroskopisch analysiert sowie einer computertomographischen Untersuchung unterzogen und weisen einen Aufbau auf, der in dieser Form bisher nicht bekannt war. Ein Schaft ist nur undeutlich ausgebildet, im Durchmesser kaum verschieden von den Federästen; die Federäste sind wechselständig (alternierend) und nur an der Basis fast gegenständig, zur Spitze hin ist die Verzweigung fast dichotom (gabelig) (http://www.cell.com/action/showImagesData?pii=S0960-9822%2816%2931193-9; Figure 3). Die ebenfalls gut erhaltenen, ca. 0,1 mm langen Federstrahlen sitzen nicht nur an den Ästen, sondern in gleicher Ausprägung auch an der Rachis; das kennt man bei heutigen Federn nur von der Basis des Schafts.
Die Autoren erwähnen mehrmals auch Follikel; das ist der in die Körperhülle eingesenkte Bereich der Federn. Eine nähere Beschreibung findet sich dazu nicht in ihrer Arbeit; auch aus den Abbildungen geht nicht hervor, woran deren Existenz erkennbar ist.
Ist die Bezeichnung „Feder“ für die fiederigen Strukturen gerechtfertigt? Es hat sich eingebürgert, jeglichen Körperbesatz von Dinosauriern als Federn oder Vorfedern zu bezeichnen. In vielen Fällen ist das nicht gerechtfertigt, sondern ausschließlich evolutionstheoretisch motiviert, wonach Vögel von Dinosauriern abstammen. Einfache Körperanhänge von Dinosauriern wiederum als Belege für diese populäre These zu verwenden, stellt einen Zirkelschluss dar.
Die Federn des von Xing et al. (2016a) beschriebenen Bernsteinfundes sind allerdings deutlich komplexer strukturiert als alle bekannten flaumartigen Strukturen bei Kompressionsfossilien. Denn immerhin gibt es zwei Ordnungen von Verzweigungen, wenn auch nicht in einer Ausgestaltung wie bei eindeutig flächigen Federn. Chuong et al. (2003) definieren Federn als komplexe Integumentanhänge, die hierarchisch verzweigt sind, aus Rachis, Federästen und Federstrahlen bestehen, aus einem Follikel heraus wachsen und besondere biochemische, morphologische und entwicklungsbiologische Eigenschaften besitzen. Die neu beschriebenen Bernsteinfedern erfüllen diese Kriterien nur zu einem kleinen Teil oder es ist unsicher, ob die Kriterien erfüllt sind. Klar ist, dass die Federn nicht flugtauglich waren. Über ihre Funktion kann nur spekuliert werden und die Funktion dürfte ohne Kenntnis des ganzen Tieres nicht bestimmbar sein.
Ist die Zuordnung des Fundes zu einem Dinosaurier gesichert? Xing et al. (2016, 3) stellen fest, dass nur zwei Wirbelkörper gut zu erkennen seien, da das erhaltene Weichgewebe so hoch verdichtet ist, dass es nur schwer von den weiteren Skelettelementen unterscheidbar ist. Daher blieben viele diagnostische Details der Knochen unklar. Der Schluss auf die Existenz von acht oder neun Wirbeln im erhaltenen Bernsteinstück erscheint daher unsicher und die Extrapolation auf die Existenz von wahrscheinlich mehr als 25 Wirbeln steht somit auf schwachem Fundament.
Dass die Form der Wirbel auf einen Theropoden hinweise, begründen die Autoren mit dem Nachweis einer bauchseitigen Kerbe auf dem Centrum (Wirbelzentrum) der Wirbelkörper, was typisch für Theropoden sei; im online-Zusatzmaterial merken die Autoren allerdings an, dass nicht ausgeschlossen werden könne, dass eine solche Kerbe auch bei Archaeopteryx und Jeholornis ausgebildet war. Auch die sehr geringe Größe des Tieres ist für einen Theropoden ungewöhnlich. Die Bestimmung des Fossils als Coelurosaurier-Theropode kann daher nicht als sicher gelten.
Was trägt der Fund zur Klärung der Entstehung von Vogelfedern bei? Xing et al. (2016a) ordnen die Federstruktur des neuen Fundes in das derzeit favorisierte Federentstehungsmodell von Prum (1999) ein. Die Struktur der beschriebenen „Schwanzfedern“ passen aber zunächst einmal zu keiner der dort hypothetisch angenommenen Formen; die Autoren ordnen sie in die Nähe von Stadium IIIB dieses Modells ein (vgl. http://www.cell.com/action/showImagesData?pii=S0960-9822%2816%2931193-9; Figure 4). Dieses Stadium ist durch Ausbildung von Federästen und Federstrahlen gekennzeichnet. Die Autoren sehen die Bernsteinfedern als Beleg dafür, dass zuerst Federstrahlen entstanden sind, bevor die Federn flächig wurden. Allerdings sind die Federn wie beschrieben einzigartig ausgebildet und passen nicht in jeder Hinsicht in Prums Modell, sie stehen eher im Zwischenbereich der als Alternativen beschriebenen hypothetischen Stadien IIIA und IIIB. Xing et al. (2016a, 7) halten es daher für möglich, dass es sich um eine spezialisierte Federform handelt, die außerhalb der Federevolutionslinie steht.
Die Bedeutung des neuen Fundes für Hypothesen zur Entstehung von Vogelfedern muss aber ohnehin als sehr gering eingestuft werden. Denn der Fund ist nach gängiger zeitlicher Einordnung ca. 60 Millionen Jahre (radiometrisch) jünger als die älteste Vogelgattung, die unzweifelhaft flächige Federn heutiger Bauart besaß (Anchiornis) und ca. 50 Millionen radiometrische Jahre (MrJ) jünger als Archaeopteryx, dessen Federn von Federn heutiger Vögel kaum zu unterscheiden sind. Aus der Unterkreide sind ab etwa 130 MrJ mit den sogenannten Gegenvögeln (Enantiornithes) und den Ornithurae („Vogelschwänze“) Dutzende von Vogelgattungen mit „fertigen“ Federn bekannt. Daher kann dem auf 99 MrJ datierten Bernsteinfund kaum eine Bedeutung für die Federevolution zugemessen werden. Darstellungen in der Wissenschaftspresse, wonach der Fund zeige, dass der Federschaft später entstand als die feinsten Federhärchen (http://www.scinexx.de/wissen-aktuell-20926-2016-12-09.html), sind daher fehl am Platz.
Problematisch ist auch, dass nur ein kleines Bruchstück erhalten ist, das zudem vermutlich von einem noch nicht ausgewachsenen Tier stammt. Rekonstruktionen eines ganzen Tieres, wie sie in Pressmeldungen verbreitet werden, sind nicht seriös. Hatte das Tier vielleicht an anderen Körperstellen „moderne“ Federn wie sie bei einer anderen Gattung aus denselben Schichten gefunden wurden (Xing et al. 2016b)? Wie waren die Federn beim ausgewachsenen Tier ausgebildet? Angesichts der Existenz verschiedenster Mosaikformen ist auch die Frage angebracht, ob es sich wirklich um einen Dinosaurier handelt. Acht oder neun Schwanzwirbel sind zu wenig, um auf den Körperbau und die Befiederung des ganzen Tieres schließen zu können.
Literatur
Chuong CM, Wu P, Zhang FC, Xu X, Yu M. Widelitz RB, Jiang TX & Hou L (2003) Adaptation to the sky: Defining the feather with integument fossils from Mesozoic China and experimental evidence from molecular laboratories. J. Exp. Zool. 298B, 42-56.
Prum RO (1999) Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291-306.
Xing L, McKellar RC et al. (2016a) A feathered dinosaur tail with primitive plumage trapped in mid-Cretaceous amber. Curr. Biol. 26, 1-9, http://dx.doi.org/10.1016/j.cub.2016.10.008
Xing L, McKellar RC, et al. (2016b) Mummified precocial bird wings in mid-Cretaceous Burmese amber. Nat. Comm. 7:12089, doi: 10.1038/ncomms12089.
Zhou Z & & Zhang F (2003) Jeholornis compared to Archaeopteryx, with a new understanding of the earliest avian evolution. Naturwissenschaften 90, 220-225.
Bilder:
http://www.cell.com/action/showImagesData?pii=S0960-9822%2816%2931193-9
http://www.zeit.de/wissen/2016-12/dinosaurier-bernstein-fund-myanmar
Autor dieser News: Reinhard Junker
© 2016, http://www.genesisnet.info/schoepfung_evolution/n242.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
08.12.15 Alte Vögel mit moderner Flugkunst
Eines der ältesten Fossilien aus der nur in der Kreide überlieferten Gruppe der „Gegenvögel“ (Enantiornithes) weist eine Reihe von Merkmalen auf, die für eine sehr gute Flugfähigkeit sprechen – trotz deutlich verschiedener Anatomie im Vergleich mit heutigen Vögeln. Gemessen am Fossilbericht sind die Gegenvögel somit gute Flieger von Anfang an.
Die fossile Überlieferung der Vögel zeigt zwei Phasen eines plötzlichen Erscheinens vielfältiger Formen. Die eine liegt am Beginn des Tertiärs (wo die auch heute vorkommenden Vogelordnungen explosiv im Fossilbericht auftauchen), die andere in der Unterkreide, in deren geologischen Schichten ebenfalls ziemlich abrupt eine große Vielfalt von Vögeln mit anderer Anatomie als den heutigen überliefert ist. Der berühmte „Urvogel“ Archaeopteryx aus dem Oberjura datiert noch ca. 20 Millionen radiometrische Jahre älter; es ist aber nach wie vor umstritten, ob er aktiv fliegen konnte oder eher ein Gleiter war (s. Archaeopteryx – Gleitflieger und Bindeglied?, https://www.genesisnet.info/index.php?News=193). Andere eindeutig befiederte Formen aus dem Oberjura (z. B. Anchiornis) gelten als flugunfähig (s. Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137).
Die Kreidevögel unterscheiden sich anatomisch mehr oder weniger deutlich von den tertiären Vögeln und zeigen manche Gemeinsamkeiten mit zeitgleich lebenden Theropoden-Dinosauriern (zweibeinige Raubdinosaurier), die als ihre stammesgeschichtlichen Vorfahren gelten. Sie werden in zwei Gruppen unterteilt, die Enantiornithes („Gegenvögel“1; sind am Ende der Kreide ausgestorben) und die Ornithurae („Vogel-Schwänze“2), zu denen u. a. auch die heutigen Vögel gehören.
Eine Forschergruppe hat nun herausgefunden, dass die Enantiornithes trotz deutlich verschiedener Anatomie im Vergleich zu heutigen Vögeln überraschenderweise vergleichbare Fähigkeiten zu einem versierten Flug besaßen. Nachgewiesen wurde dies anhand eines sehr gut erhaltenen vorderen Teils einer Vorderextremität aus einem Kalkstein von Las Hoyas, Spanien (datiert auf 125 Millionen radiometrische Jahre), die aufgrund ihres Baus von einem enantiornithinen Vogel stammt (Navalón et al. 2015). Erhalten sind auch Hand- und Armschwingen. Das Besondere an dem Fund: Es sind Details von Muskeln, Bändern und Follikeln (in der Haut befindliche Basis der Federn) aus den Bereichen zwischen den Flügelteilen und Fingern (Propatagium, Postpatagium und Patagium der Alula3) in Form von Abdrücken erhalten. Der Fund aus Las Hoyas lässt auch Verbindungen von Bindegewebsstrukturen mit den Ansätzen der Flugfedern erkennen; das gesamte komplizierte Netzwerk weist große Ähnlichkeiten mit den anatomischen Verhältnissen heutiger Vögel auf. Ein solch komplexes System aus Sehnen, Bändern, Bindegewebe und Muskeln ermöglicht eine ausgefeilte Manövrierbarkeit während des Fluges. Aus ihren Beobachtungen schließen die Autoren, dass die „Gegenvögel“ der Kreidezeit ebenso gut wie viele moderne Vögel „über den Köpfen der Dinosaurier“ fliegen konnten, wie der Leiter der Forschergruppe, L. M. Chiappe, es formuliert.4 „The anatomical match between the muscle network preserved in the fossil and those that characterize the wings of living birds strongly indicates that some of the earliest birds were capable of aerodynamic prowess like many present-day birds“ (Chiappe5). Bislang war man davon ausgegangen, dass die Gegenvögel der Unterkreide eher schlechte Flieger waren; das kann in dieser Pauschalität mit diesem Fund als widerlegt gelten. Die Gegenvögel starten gleichsam als gut ausgebildete Flieger; eine Evolution der Flugfähigkeit ist innerhalb dieser Gruppe somit nicht dokumentiert. Das Beispiel zeigt auch, dass es zu Fehlschlüssen kommen kann, wenn man die Funktionalität nur anhand von Knochenüberresten erschließen möchte (bzw. kann).
Navalón et al. (2015) stellen allgemein über die Gegenvögel am Schluss ihres Artikels fest, dass obwohl die Enantiornithinen eine Anzahl von primitiven Skelettelementen aufwiesen, bereits die frühesten unter ihnen (Protopteryx fengningensis) Vorderextremitäten mit modernen Proportionen besaßen, außerdem ein gekieltes Brustbein, einen „fortschrittlichen“ Schultergürtel mit einem Kanal für die Passage der Flugmuskeln (triosseal canal), was alles auf die Fähigkeit zu aktivem Flug und Flügelschlag ähnlich dem heutiger Vögel hinweise. Auch Ähnlichkeiten der Körperbedeckung mit heutigen Vögeln mit identischer Federanordnung würden in diese Richtung weisen.
Es sei noch angemerkt, dass auch unter den Ornithurae eine der ältesten Formen als guter Flieger angesehen wird (vgl. Der älteste „echte“ Vogel: überraschend modern, https://www.genesisnet.info/index.php?News=226).
Literatur
Navalón G, Marugán-Lobón J, Chiappe LM, Sanz JL & Buscalioni A (2015) Soft-tissue and dermal arrangement in the wing of an Early Cretaceous bird: Implications for the evolution of avian flight. Sci. Rep. 5:14864.
Anmerkungen
1 Es gibt in der Fachliteratur keine einheitliche Begründung für die Bezeichnung „Gegenvögel“. Es wird auf zwei anatomische Besonderheiten hingewiesen, die die Enantiornithes von anderen Vögeln unterscheiden: Zum einen die Art der Verwachsung einiger Fußknochen, zum anderen der Bau des Gelenks zwischen Schulterblatt und Rabenbein. Bei den Gegenvögeln sind Gelenkkopf und Gelenkpfanne im Vergleich zu allen anderen Vögeln sozusagen vertauscht. Es ist schwer denkbar, wie die eine anatomische Ausprägung in die andere evolutiv überführt werden könnte.
2 Es gibt keine eingebürgerte deutsche Bezeichnung, der Name bezieht sich auf den kurzen knöchernen Schwanz, wie er auch für die heutigen Vögel typisch ist.
3 Daumenfittich, wichtig z. B. für Manöver beim Landen.
4 http://www.sciencenutshell.com/the-bird-who-flew-above-dinosaurs-heads/
5 http://www.eurekalert.org/pub_releases/2015-10/nhmo-taf100615.php
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n233.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
09.11.15 Dino-Doppelgänger des Vogel Strauß
Ein jüngst veröffentlichter Dinosaurier-Fund bestätigt, dass die Ornithomimiden („Vogel-Nachahmer“) befiedert waren, und dokumentiert darüber hinaus eine sehr ähnliche Befiederung wie bei heutigen großen Laufvögeln – eine bemerkenswerte Parallele ohne nähere Abstammungsverwandtschaft. In die bisher favorisierten evolutionären Abstammungsfolgen passt der Fund nicht gut bzw. nur mit Zusatzannahmen, insbesondere der Annahme einer unabhängigen Entstehung typischer Elemente des Laufvogel-Bauplans und eventuell der Federn.
Die „Vogel-Nachahmer“ unter den Dinosauriern, die Ornithomimidae, sind schon seit langem bekannt, vor allem aus den geologischen Schichten der Oberkreide Ostasiens und des westlichen Nordamerikas. Sie gehörten zu den Theropoden (zweibeinig laufende Raubdinosaurier), besaßen einen schlanken Körperbau, wurden meist 3-6 Meter lang, und man vermutet, dass sie schnelle Läufer waren, vergleichbar den heutigen Laufvögeln (dafür spricht z. B., dass die Unterschenkel länger waren als die Oberschenkel, was ein Indiz für schnelles Laufen ist). Vogeltypisch war auch der zahnlose Schnabel. Der Schädel war langgestreckt und relativ klein, mit großen seitlich platzierten Augen und vermutlich gut entwickeltem Gehirn; Hirnschädel und Schnauzenregion besaßen luftgefüllte Hohlräume.

Abb. 1: Skelett von Ornithomimus. (Wikimedia: Sebastian Bergmann, https://www.flickr.com/photos/90237600@N00/64228455/, CC BY-SA 2.0)
Erst vor kurzem sind bei einigen Exemplaren auch Reste federartiger Anhänge und eine flügelartige Struktur mit verlängerten Federn (ein sogenanntes „Pennibrachium“) nachgewiesen worden (Zelenitsky et al. 2012). Die schlechte Erhaltung erlaubt keine sichere Klärung darüber, ob es sich bei den Anhängen um flächige Federn handelt. Die neuen Befunde waren insofern überraschend, als die Ornithomimiden zwar aufgrund ähnlicher Merkmale als „Vogelnachahmer“ bezeichnet (s. o.), aber ansonsten mit den Vögeln nicht als näher verwandt betrachtet werden. (Der evolutive Weg zu den Vögeln soll über andere Gruppen gelaufen sein.) Daher wurde die evolutionstheoretische Hypothese formuliert, dass schon bei den gemeinsamen Vorfahren der Ornithomimiden und der Vögel Federn ausgebildet waren.1 Daraus würde allerdings folgen, dass viele Formen danach die Federn wieder zurückgebildet oder verloren hätten. Außerdem müssten diese gemeinsamen Vorfahren im evolutionären Modell mehr als doppelt so alt wie die befiederten Formen der Ornithomimiden gewesen sein – alles Befunde, die evolutionstheoretisch nicht gut passen. Heute ist aber auch die Annahme nicht mehr tabu, dass Federn in verschiedenen evolutionären Linien mehrfach unabhängig entstanden sein könnten (Brusatte et al. 2015, R892), was nicht-zielgerichtete evolutionäre Mechanismen vor noch größere Herausforderungen stellt als die Erklärung einer einmaligen Entstehung.
Nun berichten van der Reest et al. (2016) von einem neuem Ornithomimus-Fund. Er stammt wie die anderen Ornithomimus-Funde mit Federresten aus der Oberkreide Nordamerikas (Dinosaur Park Formation of Alberta, Kanada). Das Tier war bis zu vier Meter lang; Kopf und Vorderextremitäten sind allerdings nicht erhalten. Das Besondere an diesem bereits 2009 gemachten Fund: Nicht nur Federn sind in sehr guter Erhaltung überliefert, sondern es ist auch klar erkennbar, welche Körperteile nicht befiedert waren, da im Bereich des Fußes auch fossile Spuren von Hautresten sehr gut sichtbar sind. Die Anordnung der Befiederung an Schwanz und an den Beinen und die nackten Bereiche ähneln stark den Verhältnissen heutiger großer Laufvögel wie Strauß oder Emu (Bild eines rekonstruierten Tieres z. B. hier: http://cdn.phys.org/newman/gfx/news/hires/2015/preshistoric.jpg). So sind die Unterseite des Schwanzes und die Beine ab etwa der Mitte des Oberschenkels ungefiedert. Die Forscher schließen aus der gesamten Befundsituation auf eine ähnliche Thermoregulation bei Ornithomimus wie bei den heutigen Straußen. „Die federlose Haut unter dem Körper deutet darauf hin, dass die Tiere sogar den gleichen Mechanismus nutzen, um überschüssige Wärme loszuwerden. Auch die Chemie der Federn sei von modernen Vögeln praktisch nicht zu unterscheiden, betont van der Reest.2
Da die Ornithomimiden und die heutigen großen Laufvögel im aktuell favorisierten Evolutionsmodell auf relativ weit entfernten Ästen angeordnet sind, werden die genannten weitreichenden Ähnlichkeiten als Konvergenzen* interpretiert; sie werden also nicht als Indizien für gemeinsame Abstammung gewertet. Erstaunlich ist vor diesem Hintergrund die Auffassung eines der Autoren, der Fund erhärte die klassische These, nach der Vögel eine Gruppe innerhalb der Dinosaurier darstellen.3 Das ist aus zwei Gründen schwer nachvollziehbar:
* Konvergenzen (= unabhängig entstandene Ähnlichkeiten)
- Die Schichten, in denen Ornithomimus gefunden wurde, werden auf etwa 72 Millionen radiometrische Jahre datiert (Oberkreide, Campanium). Der berühmte „Urvogel“ Archaeopteryx mit unbestritten „moderner“ Befiederung hat das doppelte radiometrische Alter, und zahlreiche eindeutig flugfähige Vogelgattungen der Unterkreide sind ebenfalls deutlich älter datiert als Ornithomimus.
- Da die Ornithomimiden auf einen systematischen Ast gestellt werden, der nicht zu den Vögeln überleitet, können sie auch nicht in eine enge phylogenetische Beziehung zu den Vögeln gebracht werden. Eine evolutive Verbindung Vögel-Dinosaurier wird durch die Ornithomimiden allenfalls sehr indirekt unterstützt (z. B. durch die Annahme mutmaßlicher entfernterer gemeinsamer Vorfahren; s. o.), und zweifellos gibt es für eine solche Verbindung bessere Kandidaten unter den Theropoden. Die Ornithomimiden sind für den Übergang von den Dinosauriern zu den Vögeln jedenfalls keine geeigneten Kronzeugen – im Gegenteil: Sie zeigen, dass auch die genannten weitreichenden Ähnlichkeiten nicht ohne weiteres als Hinweise auf eine gemeinsame Abstammung gedeutet werden können, sondern unter Umständen (wie hier) als Konvergenzen anzusehen sind. An die steigende Zahl von Konvergenzen auch von Schlüsselmerkmalen scheint man sich im Rahmen des Evolutionsmodells zwar zu gewöhnen; die damit verbundenen evolutionstheoretischen Fragen nach ihrer unabhängigen Entstehung bleiben dennoch ungelöst und fordern komplett neue Evolutionsmechanismen (Diskussion dazu bei Braun 2012).
Keine Auskunft kann der neue Fund geben, wie im Verlauf der Evolution Federn entstanden sind. Das gilt insbesondere deshalb, weil das Federkleid von Ornithomimus als zurückgebildet interpretiert wird. Eine evolutionäre Neubildung von Federn bei Ornithomimus wäre noch unwahrscheinlicher, da dann noch weiter reichende Konvergenzen angenommen werden müssten.
Literatur
Braun HB (2012) Warten auf einen neuen Einstein. Stud. Integr. J. 19, 12-19.
Brusatte SL, O’Connor JK & Jarvis ED (2015) The origin and diversification of birds. Curr. Biol. 25, R888-R898.
Van der Reest AJ, Wolfe AP & Currie PJ (2016) A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada. Cretaceous Res. 58, 108-117.
Zelenitsky DK, Therrien F, Erickson GM, DeBuhr CL, Kobayashi N, Eberth DA & Hadfield F (2012) Feathered non-avian dinosaurs from North America provide insight into wing origins. Science 338, 510-514.
Anmerkungen
1 vgl.: „Alberta researcher’s discovery offers insight into feathered dinosaurs“, http://www.theglobeandmail.com/news/national/alberta-researchers-discovery-offers-insight-into-feathered-dinosaurs/article27024287/
2 „Dino hatte Schwanzfedern und nackte Beine“, http://www.spektrum.de/news/dino-hatte-schwanzfedern-und-nackte-beine/1373283
3 „Ornithomimus dinosaur with preserved tail feathers and skin tightens linkages between dinosaurs and birds“, http://phys.org/news/2015-10-ornithomimus-dinosaur-tail-feathers-skin.html
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n232.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
22.07.15 Zhenyuanlong – befiederter Dinosaurier, flugunfähiger Vogel oder …?
Mit über 1,50 m Größe ist Zhenyuanlong die größte Dinosaurier-Gattung mit Federn, die der Fiederung heutiger Vögel gleicht – falls diese Gattung wirklich ein Dinosaurier war. Einige Merkmale sprechen eher dafür, dass es sich um einen sekundär flugunfähigen Vogel handelt. Wahrscheinlich sind bislang zu wenig Details bekannt, um die seltsame und unerwartete Merkmalskombination dieses Fossilfundes erklären zu können. Wie auch in anderen Fällen passt das neu entdeckte Fossil nicht in bisherige evolutionstheoretische Vorstellungen.
Erneut wurde in Unterkreide-Schichten in der Liaoning-Provinz Chinas ein ungewöhnliches Fossil entdeckt, das als befiederter Dinosaurier interpretiert wird (Lü & Brusatte 2015). Das Fossil besitzt eine ungewöhnliche Merkmalskombination, die keineswegs evolutionstheoretischen Erwartungen an Übergangsformen zwischen Dinosauriern und Vögeln entspricht. Es geht um Zhenyuanlong suni; diese neue Art wird zu den Dromaeosauriden gestellt, eine Dinosaurier-Familie, die von den meisten Paläontologen in nächste Nähe zu den Vögeln gebracht wird. Diese Stellung ist aber aus guten Gründen umstritten und Zhenyuanlong dürfte Kritiker der Dino-Vogel-Theorie bestärken. Denn Zhenyuanlong besitzt zwar einerseits Federn wie heutige Vögel, mehrlagig angeordnet, mit Schaft, Feinstrukturen und z. T. mit asymmetrischer Form, was als Indiz für die Flugtauglichkeit zu werten ist (die Asymmetrie ist eine entscheidende Voraussetzung, dass Auftrieb erzeigt werden kann).1 Andererseits aber sind seine Vorderextremitäten viel zu kurz, um einen Flug zu ermöglichen. Wofür dienten dann die gut ausgebildeten Federn?
Dazu kommt, dass Zhenyuanlong mit einer Größe von über 1,5 m für einen gefiederten Dinosaurier ungewöhnlich groß war – der größte mit unzweifelhaften Federn, aber auch deutlich zu groß, um in eine engere Vorfahrenschaft zu den Vögeln gestellt werden zu können, da ein evolutiver Übergang zu Vögeln nur bei kleinen Formen flugtechnisch denkbar erscheint (worüber unter den Fachleuten Konsens besteht). Erst vor etwa einem Jahr erschien eine Studie, wonach die Dinosaurier-Gattungen, die evolutionär in eine Reihe zu den Vögeln gestellt werden, zunehmend kleiner geworden seien – ein Trend der als wesentliche Voraussetzung für die Entstehung von Vögeln gewertet wird (Lee et al. 2014). Zhenyuanlong passt deshalb (neben anderen Formen) gar nicht ins Bild.
Wenn die neue Gattung trotz Besitz „moderner“ Federn nicht in eine Vorfahrenstellung zu den Vögeln einzugliedern ist und flugunfähig war, was bleibt als Erklärung für den Besitz der Federn? Die Forscher neigen zur Auffassung, die Federn hätten als Schauorgane eine wichtige Rolle bei der Balz gespielt. Aber weshalb sollten sich zu diesem Zweck asymmetrische, eigentlich flugtaugliche Federn evolutiv herausbilden? Die Schaufunktion könnte auch durch ein gefärbtes, gemustertes Schuppenkleid oder durch einfache Haare erfüllt werden, wie sie bei vielen Dinosaurier-Gattungen bekannt sind. Ausgefeilte Federstrukturen tragen dazu nichts bei. Die Idee, dass die Flügel zum Schutz des Geleges vor ungünstiger Witterung2 oder vor Feinden3 evolviert seien, ist aus demselben Grund unglaubhaft. Auch dafür bräuchte es keine komplexen, flugtauglichen Federn.
Die Deutung, dass Zhenyuanlong sich auf einem evolutiven Weg zur Flugfähigkeit befand, kann ebenfalls ausgeschlossen werden. Denn der Erwerb ausgefeilter flugtauglicher Federn erfordert unter anderem entsprechende Selektionsdrücke. Diese sind aber nicht gegeben, solange es nicht wenigstens eine rudimentäre Flugfähigkeit gibt. Diese Deutung wird allgemein und auch von Lü & Brusatte (2015) als sehr unwahrscheinlich angesehen.
Es bleibt als weitere Deutungsmöglichkeit, dass es sich um eine sekundär flugunfähige Form handelt. Diese Möglichkeit wird von einigen Forschern auch bei kleineren, gefiederten Dinosauriern wie Caudipteryx und Protarchaeopteryx diskutiert (z. B. Ruben & Jones 2000, Olson 2002, Feduccia & Czerkas 2015). Das würde heißen, dass Zhenyuanlong kein Dromaeosaurier und folglich kein befiederter Dinosaurier ist, sondern Abkömmling eines Vogels. Flugunfähige Formen, die von flugfähigen abstammen, sind aus der heutigen Vogelwelt zahlreich bekannt. Mit diesem Status würde Zhenyuanlong genauso wie manche andere „gefiederte Dinosaurier“ nichts zum Verständnis der Evolution der Vögel beitragen.
Angesichts der ungewöhnlichen und unerwarteten Merkmalskombination von Zhenyuanlong erscheint es angebracht, mit Deutungen vorsichtig zu sein. Möglicherweise wissen wir immer noch einfach zu wenig über die Struktur- und Funktionsbeziehungen und über die Ökologie bei dieser und anderen rätselhaften Formen. So ist auch der Streit über die Flugfähigkeit des „Urvogels“ Archaeopteryx aus dem älteren Oberjura nicht beigelegt (vgl. Longrich et al. 2012; Carney et al. 2012, Foth et al. 2014; vgl. Archaeopteryx – Gleitflieger und Bindeglied?, https://www.genesisnet.info/index.php?News=193; Immer rätselhafter: Der „Urvogel“ Archaeopteryx, https://www.genesisnet.info/index.php?News=216). Auch die Gefiederausstattung des ebenfalls im Oberjura entdeckten Anchiornis ist ziemlich rätselhaft (Hu et al. 2009; vgl. Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137).
Für Zhenyuanlong gilt schließlich wie für die meisten befiederten Dinosaurier (sofern diese überhaupt tatsächlich Dinos waren), dass sie stratigraphisch (in der Schichtenfolge) deutlich zu spät überliefert sind, um als Vorfahren der Vögel gelten zu können. Für eine Vorfahrenform könnten die Unterkreide-Gattungen allenfalls modellhaften Charakter haben, aber mit den Fragezeichen, die besonders (aber nicht nur) bei Zhenyuanlong anzubringen sind (s. o.).
Lu & Brusatte (2015) weisen noch darauf hin, dass die Merkmalskombination von Zhenyuanlong die Abstammungsverhältnisse unter den Dromaeosauriden verkompliziert, weniger stimmig als zuvor mache und die Annahme einer großen Zahl von Homoplasien (Konvergenzen* oder Rückentwicklungen) erfordere; so müsse nun z. B. angenommen werden, dass kurze Arme mehrfach unabhängig entstanden sind (Lü & Brusatte 2015, 9). Das heißt, Zhenyuanlong passt nicht in bisherige Stammbaumrekonstruktionen. Auch in dieser Hinsicht ist der Fund evolutionstheoretisch eine Überraschung.
*Konvergenzen (= ähnliche, aber unabhängig entstandene Merkmale)
Literatur
Carney RM, Vinther J, Shawkey MD, D’Alba L & Ackermann J (2012) New evidence on the colour and nature of the isolated Archaeopteryx feather. Nature Comm. 3, Art.-No. 637, doi:10.1038/ncomms1642.
Feduccia A & Czerkas SA (2015) Testing the neoflightless hypothesis: Propatagium reveals flying ancestry of oviraptorosaurs. J. Orn., doi:10.1007/s10336-015-1190-9.
Foth C, Tischlinger H & Rauhut OWM (2014) New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature 511, 79-82.
Hu D, Hou L, Zhang L & Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 460-463.
Lee MSY, Cau A, Naish D & Dyke GJ (2014) Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds. Science 345, 562-566.
Lü J & Brusatte SL (2015) A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution. Sci. Rep. 5:11775, doi: 10.1038/srep11775.
Longrich NR, Vinther J, Meng Q, Li Q & Russell AP (2012) Primitive wing feather arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Curr. Biol. 22, 2262-2267.
Olson SL (2002) Review: New perspectives on the origin and early evolution of birds. Proceedings of the International Symposium in Honor of J.H. Ostrom. The Auk 119, 1202-1205.
Ruben JA & Jones TD (2000) Selective factors associated with the origin of fur and feathers. Amer. Zool. 40, 585-596.
Genesisnet-Newsbeiträge zu Urvögeln:
- Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137
- Fällt eine Ikone vom Sockel?, https://www.genesisnet.info/index.php?News=174
- Archaeopteryx – Gleitflieger und Bindeglied?, https://www.genesisnet.info/index.php?News=193
- Immer rätselhafter: Der „Urvogel“ Archaeopteryx, https://www.genesisnet.info/index.php?News=216
- Der älteste „echte“ Vogel: überraschend modern, https://www.genesisnet.info/index.php?News=226
Anmerkungen
1 „‘Sie waren wie bei einem Adler oder Geier‘, sagt Co-Autor Steve Brusatte von der University of Edinburgh.“ http://www.wissenschaft.de/erde-weltall/palaeontologie/-/journal_content/56/12054/7294559
2 http://www.spektrum.de/news/bislang-groesster-dino-mit-fluegeln-gefunden/1356293
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n230.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
10.06.15 Hühnchen mit Dinosaurier-Schnauze?
Durch eine Manipulation von Hühnerembryonen ist es Wissenschaftlern gelungen, die Schnäbel in ihrer Entwicklung im Hühnerei so zu verändern, dass sie in mancher Hinsicht Schnauzen von manchen Dinosauriern ähneln. Für eine mutmaßliche evolutive Abstammung der Vögel von Dinosauriern gibt dieses Experiment aber keine nennenswerten Anhaltspunkte.
„Kiefer statt Schnäbel: Forscher erschaffen ‚Dino-Hühner‘ im Labor“; „Rückbau der Vögel zu Dinos“; „Forscher züchten Hühner-Embryos mit Dino-Schnauzen“ – das eine kleine Auswahl von Überschriften der Tages- und Wissenschaftspresse1, mit denen kürzlich auf ein interessantes Experiment aufmerksam gemacht wurde. Was haben die Wissenschaftler herausgefunden?
Mit den enormen Fortschritten in der Erforschung des Erbguts verschiedenster Arten von Lebewesen sind Wissenschaftler heute in der Lage, zunehmend die genetischen Grundlagen von Organen und ihrer korrekten Ausformung herauszufinden. Es muss aber beachtet werden, dass es bei den dabei genutzten Genen nur um einen notwendigen Teilaspekt, aber nicht um hinreichende Erklärung der Formbildung in der Embryonalentwicklung handelt: Wann werden welche Gene aktiviert? Das Genom* ist nicht der Bauplan, wie man heute weiß. Dennoch verhilft die Erforschung genetischer Zusammenhänge zunehmend zu einem besseren Verständnis der Entwicklungszusammenhänge während der Ontogenese**. Das gilt auch für die Erforschung der bei der Schnabelbildung bei Vögeln beteiligten Gene. Hier konnten Wissenschaftler um Bhart-Anjan Bhullar von der Yale University zeigen, dass in der zukünftigen Gesichtsregion von Vögeln zwei Proteine, Fgf8 (Fibroblast growth factor 8) und Lef1 (das zum Wnt-Signalweg gehört) sowie die nachgeschalteten Kaskaden eine wichtige Rolle bei der Ausbildung des Schnabels spielen. Diese beiden Proteine werden im Vergleich zu Reptilien in einer sehr viel größeren Region im Kopfbereich produziert. Das hat zur Folge, dass zwei Knochen, die bei anderen Wirbeltieren paarig ausgebildet sind, nämlich die Praemaxillarknochen, miteinander zu einem einzigen Knochen verwachsen, die den Schnabel bilden. Dieser Befund motivierte die Idee, dass eine Änderung in der Produktion dieser Proteine zur Bildung des Vogelschnabels beigetragen haben könnte.
*Genom (= das gesamte Erbgut einer Art)
**Ontogenese (= Entwicklung bis zum ausgewachsenen Organismus)
Bhart-Anjan Bhullar und Mitarbeiter führten nun folgendes Experiment an Hühnerembryonen durch (Bhullar et al. 2015): Sie unterdrückten die Expression* der die beiden Proteine codierenden Gene in der Mitte des sich entwickelnden Gesichts mithilfe von Chemikalien, womit sie gleichsam einen mutmaßlichen Evolutionsschritt zurückgingen. Diese Prozedur führte tatsächlich dazu, dass der Bereich des Gesichtes seine Form änderte, so dass anstelle des typischen Vogelschnabels ein Gebilde entstand, das teilweise eher der Schnauze eines Alligators oder eines Dinosauriers mit langer Schnauze ähnelte. Die Praemaxillarknochen wuchsen nicht zusammen und auch die Form des Gaumens änderte sich in einer Weise, die den Gaumenknochen von Dinosauriern ähnelte (vgl. Pennisi 20152). Dazu verglichen die Forscher die Ausformung der betreffenden Knochen der manipulierten Hühnerembryonen mit den entsprechenden Knochen verschiedener Reptilienarten.
*Expression (= Ablesung eines Gens, „Übersetzung“ in mRNA bzw. Proteine)
Ist damit ein Schritt zurück in der Evolution gelungen? Die Details des Experiments sprechen dagegen. So weist der Entwicklungsbiologe Ralph S. Marcucio darauf hin, dass die eingesetzte Gen-unterdrückende Substanz schädliche Nebenwirkungen hat und meint, sie führe lediglich zum Absterben von Gewebe.3 Außerdem gehe die Produktion von Fgf8 im Bereich des Gesichts schon zurück, bevor überhaupt die Praemaxillarknochen gebildet werden, womit die Bedeutung von Fgf8 für die Schnabelbildung in Frage gestellt sei. Die genetischen Grundlagen der Schnabelbildung seien viel komplizierter.
Ein Vergleich des manipulierten Hühnchenschädels mit dem Schädel eines Alligators (siehe Abbildung bei Pennisi 2015, online z. B. hier: www.nature.com/news/dino-chickens-reveal-how-the-beak-was-born-1.17507) unterstreicht den Eindruck, dass durch die Manipulation der Forscher dem Vogelschädel notwendiges Material vorenthalten wird: Insgesamt sieht er nach wie vor deutlich vogelähnlicher als reptilähnlich aus und ist viel komplexer als der Reptilschädel. Der Vergleich des normalen Vogelschnabels mit dem manipulierten erweckt gleichsam den Eindruck, als versuche der Organismus, mit dem unzureichenden Material zum bestmöglichen Ergebnis zu gelangen. Im Bild gesprochen: Die Grundstruktur der „Baustelle“ Vogelschnabel ist dieselbe und deutlich verschieden von der Baustelle „Reptilschnauze“, doch es fehlt teilweise am nötigen Baumaterial. In diesem Sinne äußert sich auch Bhullar: „The altered chicks still had a flap of skin over their would-be beaks, so the difference is not obvious, … Looking at these animals externally, you would still think it’s a beak. But if you saw the skeleton, you’d just be very confused, … I would not say we gave birds snouts“ (Callaway 2015; Hervorhebung nicht im Original). Außerdem ist das Versuchsergebnis nicht einheitlich: manche manipulierten Embryonen haben deutlich geteilte und verkürzte Praemaxillarknochen, bei anderen sind sie teilweise verwachsen, wieder andere zeigen gar keine besonderen Unterschiede zur normalen Situation (Callaway 2015).
Anders ist die Situation beim veränderten Gaumenknochen, der in den manipulierten Embryonen stärker mit Nachbarknochen verwachsen ist als in den normalen Individuen. In diesem Fall könnte eher von einem Rückschlag in die hypothetische evolutionäre Vergangenheit gesprochen werden. Hier ist eine vogeltypische Differenzierung gestört, die sonst einen Beitrag zur Beweglichkeit des Schnabels leisten würde (vgl. Pennisi 2015).
Die Situation ist ähnlich wie bei Atavismen (Als Atavismen bezeichnet man Rückschläge in mutmaßliche frühere stammesgeschichtliche Stadien, s. https://www.genesisnet.info/index.php?Artikel=41304&Sprache=de&l=1): Es gibt eine enorme Anzahl verschiedener Missbildungen, doch nur ganz wenigen davon könnte evolutionsbiologische Aussagekraft zugestanden werden, nämlich denjenigen, die mehr oder weniger zufällig auch einmal an Strukturen der angenommenen Vorfahren erinnern; in diesem Beispiel ausgelöst durch eine experimentelle Manipulation. Pro Evolution kann hier somit nur sehr selektiv argumentiert werden.
Spektrum der Wissenschaft bemerkt, dass „noch offen sei“, „welche genetischen Mechanismen einstmals die veränderte Genaktivität bewirkten“.4 Aus evolutionsbiologischer Sicht ist das die entscheidende Frage. Aber nicht nur das ist kritisch anzumerken. Es sei an die von Marcucio geäußerten Einwänden erinnert (s. o.). Es ist klar, dass mit den beiden Genen fgf8 und lef1 nur einige Elemente der gesamten genetischen Grundlagen erfasst sein können, die zur korrekten Ausformung des Vogelschnabels notwendig sind. Das Ausschalten oder die Manipulationen einzelner Gene bringen Erkenntnisse über deren Rolle im komplexen Ganzen. Aber es wäre angesichts der Komplexität der Formbildungsprozesse unsachgemäß, allein daran wesentliche evolutionäre Schritte zur Umbildung von Reptilienschnauen zu Vögeln festmachen zu wollen.
Angesichts der Details der Forschungsergebnisse und der genannten kritischen Anmerkungen, die die Bearbeiter zum Teil selber äußern, ist auch klar: Die eingangs zitierten reißerischen Überschriften sind nicht seriös und geben auch nicht wieder, was die Wissenschaftler selber behaupten. Sie sind eher geeignet, an Forschung interessierte Zeitgenossen irrezuführen, womit der Wissenschaft letztlich ein Bärendienst erwiesen wird.
Literatur
Bhullar BA, Morris ZS et al. (2015) A molecular mechanism for the origin of a key evolutionary innovation, the bird beak and palate, revealed by an integrative approach to major transitions in vertebrate history. Evolution, doi: 10.1111/evo.12684
Callaway E (2015) ‚Dino-chickens‘ reveal how the beak was born Chicken embryos have been altered so that the birds grow dinosaur-like snouts. Nature News, doi:10.1038/nature.2015.17507
Pennisi E (2015) How birds got their beaks. Science 348, 744.
Anmerkungen
1 http://derstandard.at/2000015769961/Kiefer-statt-Schnaebel-Forscher-erschaffen-Dino-Huehner-im-Labor; diepresse.com/home/science/4730284/Ruckbau-der-Vogel-zu-Dinos; http://www.nachrichten.de/panorama/Zurueckdrehen-der-Evolution-Forscher-zuechten-Huehner-Embryos-mit-Dino-Schnauzen-aid_5821237407263981857.html (Zugriff jeweils 5. 6. 15)
2 „In many vertebrates, this bone is flat and fused to surrounding bones. But in birds, it’s reduced and disconnected, which frees the top part of the bill to move upward, expanding birds’ gape. In the treated chick embryos, the palate looked more like it does in other vertebrates: flat and seemingly reconnected to the jaw bones.“
3 http://www.nytimes.com/2015/05/12/science/reverse-engineering-birds-beaks-into-dinosaur-bones.html (Zugriff 5. 6. 15)
4 http://www.spektrum.de/alias/bilder-der-woche/ein-huehnchen-mit-dino-schnabel/1346081
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n228.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
19.05.15 Buntes Merkmalsmosaik: Ein „Schnabeltier“ unter den Raubdinosauriern
Als „Schnabeltier der Dinosaurier“ wird eine neu beschriebene Theropoden-Gattung aus Chile beschrieben. Denn wie das Schnabeltier mit Haaren, Milchdrüsen, Eierlegen und einem Hornschnabel eine ungewöhnliche Merkmalskombination aufweist, so vereinigt auch Chilesaurus eine „bizarre“ Mischung von Merkmalen. Einmal mehr entspricht dies nicht evolutionstheoretischen Erwartungen und wirft die Frage auf, inwiefern Merkmale des Körperbaus verlässliche Indizien für Verwandtschaftsverhältnisse sein können, aber auch, wie ein solcher Merkmalsmix auf evolutionärem Wege entstehen kann.
Die „eierlegende Wollmilchsau“ gibt es zwar nur sprichwörtlich, aber manche Geschöpfe verbinden durchaus recht unterschiedliche Merkmale zu einem kuriosen Mix. Relativ bekannt ist das heute lebende Schnabeltier, ein Säugetier mit Haarkleid und Milchdrüsen, aber auch mit einer eierlegenden Fortpflanzungsweise, einem eigenartigen Hornschnabel, einem Ruderschwanz und mit Giftspornen an den Hinterbeinen; Merkmale, die sonst verschiedenen Tiergruppen zugeordnet werden.
Kürzlich ist nun bekannt geworden, dass auch die Dinosaurier eine Art „Schnabeltier“ in ihren Reihen haben: ein pflanzenfressender Theropode – eigentlich ein Widerspruch in sich, da es bislang zum Kennzeichen dieser zweibeinigen Dinosaurier gehört, räuberisch zu leben. Populär wurden die Theropoden nicht zuletzt durch den Science-Fiction-Film Jurassic Park, vor allem aber auch, weil aus ihren Reihen eine evolutionäre Linie zu den Vögeln führen soll.
Die Art, um die es geht, wurde schon vor einigen Jahren von einem Jungen namens Diego Suarez in Chile in Schichten des oberen Jura entdeckt. Neuerdings wurde sie anhand weiterer, gut erhaltener Funde genauer untersucht und zu Ehren des Finders Chilesaurus diegosuarezi genannt (Novak et al. 2015). Die Fundschicht entspricht altersmäßig etwa den Solnhofener Plattenkalken, in denen der „Urvogel“ Archaeopteryx gefunden wurde.

Abb. 1: Skelett von Chilesaurus. (Wikimedia: Evelyn D’Esposito – https://www.flickr.com/photos/zigzagfrenetico/25980265823/, CC BY-SA 2.0)
Wie bei allen Theropoden handelt es sich bei Chilesaurus um einen Zweibeiner; er lief aufrecht auf kräftigen Hinterbeinen. Ungewöhnlich ist zunächst sein Pflanzenfressergebiss; die Zähne waren klein und spatelförmig; aber Chilesaurus hatte noch weitere Überraschungen parat. Eine phylogenetische Analyse (basierend auf Merkmalsvergleichen) ergab einerseits, dass Chilesaurus an die Basis der Gruppe der Tetanurae1 zu stellen ist, andererseits erscheinen die einzelnen Merkmale gleichsam aus ganz verschiedenen Gruppen zusammengesetzt. Novas et al. (2015) verglichen einzelne Körperregionen von Chilesaurus gesondert mit den entsprechenden Ausprägungen verschiedener Dinosauriergruppen. Dabei stellte sich heraus: Die kräftigen Beine samt Fuß und Fußgelenk waren wie bei Sauropoden ausgebildet (Sauropoden sind große vierbeinige Dinosaurier mit langem Hals und langem Schwanz, z. B. Brontosaurus), dazu passt auch der relativ kleine Kopf. Das Achsenskelett gleicht dem von Ceratosauriern (zweibeinig laufende Fleischfresser, die häufig Kopfornamente besaßen); die Vorderbeine, der Schultergürtel und Hinterbeine passen zu den Tetanuren; das Becken wiederum ist typisch für Coelurosaurier2; der Bau der Wirbel ist theropodenartig. Die Vorderextremitäten wurden als Arme benutzt; allerdings hatte Chilesaurus statt der für Theropoden üblichen Klauen nur zwei stumpfe Finger.
Angesichts dieser völlig unerwarteten Merkmalskombination wundert es nicht, dass die Bearbeiter (Novas et al. 2015) Chilesaurus als „bizarr“ bezeichnen; er repräsentiere einen extremen Fall einer „Mosaik-Evolution“ unter den Dinosauriern. Koautor Martin Ezcurra von der Universität Birmingham hält ihn für einen „der interessantesten Fälle von konvergenter Evolution in der gesamten Geschichte des Lebens“.3 Und ein weiterer Mitautor, Alexander Vargas, lässt verlauten: „Ich denke, er hat es verdient, als das Schnabeltier der Dinosaurier bezeichnet zu werden.4
Kommentar
Der bunte Merkmalsmix von Chilesaurus erfordert im Rahmen des Evolutionsmodells die Annahme mehrerer Konvergenzen oder Rückentwicklungen (Letzteres im Bereich des Fußes [Novas et al. 2015, 4]). Das heißt: Eine Reihe von Merkmalen müsste sich unabhängig in verschiedenen Linien evolutionär herausgebildet haben, z. B. die pflanzenfressende Ernährungsweise. Auf wissenschaft.de3 wird Martin Ezcurra mit dem Satz zitiert, Chilesaurus liefere ein gutes Beispiel dafür, wie Evolution funktioniert. Das ist eigentlich ein überraschendes Statement. Denn ein solcher Merkmalsmix war nicht erwartet worden, und zwar gerade deshalb, weil man von einer Evolution der Lebewesen ausgeht. Die Tatsache, dass man Arten und größere Gruppen problemlos in ein hierarchisches, eingeschachteltes System einordnen kann, ist (oder war nur noch?) eines der klassischen Argumente für Evolution. Das scheint, je mehr man über die Lebewesen weiß, desto weniger möglich zu sein, was folglich das Argument für Evolution schwächt. Chilesaurus ist diesbezüglich ein besonders eindrucksvolles Beispiel. Dass Evolution so funktionieren soll, dass es viele Konvergenzen gibt, stellt lang vertretene Überzeugungen auf den Kopf. Und wenn Evolution wirklich so funktionieren würde, wie könnte man dann überhaupt noch auf der Basis von Merkmalsvergleichen Abstammungsverhältnisse ermitteln? Wenn ähnliche Merkmale gleichermaßen durch Abstammung von einem gemeinsamen Vorfahren wie auf unabhängigen Wegen entstehen können (ohne Abstammungsbeziehung), wie könnten sie dann noch Marker für Abstammungsverwandtschaft sein?
Das Statement von Ezcurra ist auch aus einem zweiten Grund fragwürdig: Der unerwartete Merkmalsmix mit den anzunehmenden Konvergenzen sagt an sich gar nichts darüber aus, wie Evolution funktioniert. Auf galileo.tv heißt es dazu: „Grund für die Ausprägung so verschiedener Merkmale ist wohl konvergente Evolution.5 Das ist natürlich keine Erklärung – kein Grund –, sondern nur eine Beschreibung bzw. Benennung bzw. Zuschreibung. Wie es dazu kommen kann, dass vielfach in einer Art konvergente Merkmalsausprägungen auftreten, ist nicht Gegenstand der Arbeit von Novak et al., aber für evolutionäre Hypothesen ist das eine Herausforderung.
Das Beispiel Chilesaurus mahnt abgesehen von diesen grundsätzlichen Fragen auch zur Vorsicht bei der Deutung unvollständig erhaltener Fossilien: Die Ähnlichkeit verschiedener Arten in einem Teil des Bauplans lässt nicht sicher schließen, dass auch die restlichen Teile bauplanähnlich sind. Der gut erhaltene Chilesaurus zeigt, dass das nicht so sein muss.
Literatur
Novas FE, Salgado L, Suárez M, Agnolín FL, Ezcurra MND, Chimento NSR, de la Cruz R, Isasi MP, Vargas AO & Rubilar-Rogers D (2015) An enigmatic plant-eating theropod from the Late Jurassic period of Chile. Nature, doi: 10.1038/nature14307.
Video
Anmerkungen
1 Ein Taxon, das eine Reihe von Untergruppen der Theropoden umfasst; der Name bedeutet „starre Schwänze“.
2 Novas wird bei nationalgeographic.com jedoch so zitiert: „Pelvic bones from the fossils, which dated to about 150 million years ago, at first appeared to be from the ornithischian group, which includes Stegosaurus and Triceratops“ (http://news.nationalgeographic.com/2015/04/150427-theropod-dinosaur-vegetarian-rex-science/).
4 https://www.wired.de/collection/latest/das-schnabeltier-unter-den-dinosauriern
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n227.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
15.05.15 Der älteste „echte“ Vogel: überraschend modern
Ein sehr gut erhaltener Fossilfund eines amselgroßen Vogels aus der Jehol-Gruppe Nordchinas (Unterkreide) erweist sich als überraschend „modern“. Die neue Gattung Archaeornithura wird zur Gruppe der Ornithurae gestellt und ist deren ältester fossiler Nachweis. Da der „Urvogel“ Archaeopteryx auf einen evolutionären Seitenast gestellt wird, gilt nun Archaeornithura als ältester „echter“ Vogel – der aber alles andere als primitiv ist.
Von einem bemerkenswerten Fund eines Vogelfossils aus der fossilreichen Jehol-Gruppe im Nordosten Chinas berichten Min Wang und Kollegen von der Chinese Academy of Science, Beijing. Die etwa amselgroße Gattung Archaeornithura meemannae wird als ältester echter Vogel aus der Gruppe der Ornithurae (s. u.) eingestuft. Zu den Ornithurae werden auch die heute lebenden Vögel gerechnet. Bemerkenswert ist Archaeornithura wegen seines „modernen“ Aussehens – der älteste bekannte Vertreter der Ornithurae ist weit davon entfernt, zu den primitivsten seiner Gruppe zu gehören (Rekonstruktion unter https://i.dailymail.co.uk/i/pix/2015/05/06/11/285E7F9800000578-3069971-Dubbed_Archaeornithura_meemannae_the_species_illustrated_had_fan-a-8_1430906577914.jpg).
{kind=link}

Abb. 1: Fossil von Cruralispennia multidonta. Beschriftung in Übersetzung: „Cruralispennia multidonta Holotypus (IVPP V21711). (a) Photographie; (b) Strichzeichnung. ca, Schwanzwirbel; cv, Halswirbel; il, Darmbein; is, Sitzbein; lad, linkes Alularglied; lco, linkes Korakoid; lde, linkes Dentarium; lfe, linker Oberschenkelknochen; lhu, linker Oberarmknochen; lmd, linkes Hauptglied; lpd, linkes Tretglied; lra, linker Radius; lta, linker Tarsometatarsus; lti, linker Tibiotarsus; lul, linke Ulna; pu, Schambein; py, Pygostyle; qu, Quadrate; rco, rechtes Coracoid; rfe, rechtes Femur; rhu, rechter Humerus; rmd, rechter Hauptfinger; rpd, rechter Fußfinger; rra, rechter Radius; rsc, rechtes Schulterblatt; rta, rechter Tarsometatarsus; rti, rechter Tibiotarsus; rul, rechte Ulna; sk, Schädel; st, Sternum; sy, Synsacrum; tv, Brustwirbel. Die weißen Kreise (nummeriert 1-5) und der Kasten zeigen die Lage der Federn bzw. der histologischen Proben an. Maßstabsbalken, 10 mm.“ (Wikimedia: Wang, Minet al. (2017) A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nature Communications. 8: 14141. doi:10.1038/ncomms14141., CC BY 4.0)
Zur besseren Einordnung dieser neuen Art, von der zwei sehr gut erhaltene Exemplare fossil überliefert sind, muss ein wenig ausgeholt werden. Die Vögel werden in zwei Gruppen unterteilt, die Enantiornithes und die Ornithurae. Die Enantiornithes (deutsch: „Gegenvögel“) werden den anderen Vögeln (den Ornithurae) „gegenübergestellt“, weil bei ihnen die Gelenke zwischen dem Schulterblatt und dem Rabenbein im Vergleich mit den heute lebenden Vögeln (und den anderen Ornithurae) genau anders herum angeordnet sind: Bei den Enantiornithinen liegt am Schulterblatt eine Gelenkpfanne und am Rabenbein ein Gelenkzapfen vor; bei den Ornithuren ist es genau umgekehrt. Diese beiden gegensätzlichen Konstellationen können kaum auseinander hervorgegangen sein; deshalb werden die Enantiornithes und die Ornithurae in evolutionär zwei getrennte Linien gestellt.1
Der berühmte „Urvogel“ Archaeopteryx wird keiner der beiden Gruppen zugeordnet, sondern als phylogenetisch älter eingestuft als der mutmaßliche gemeinsame Vorfahr der Enantiornithinen und Ornithurinen und wie einige andere Gattungen aus dem Jura und der Unterkreide auf ein evolutionäres „Nebengleis“ gestellt. (Die Diskussion über seine stammesgeschichtliche Position und der überaus umfangreiche und kontroverse Gelehrtenstreit über die Art des Fluges bei Archaeopteryx sind bis heute nicht beigelegt; vgl. auch Fällt eine Ikone vom Sockel?, https://www.genesisnet.info/index.php?News=174 und Immer rätselhafter: Der „Urvogel“ Archaeopteryx, https://www.genesisnet.info/index.php?News=216.)
Die Enantiornithes bilden eine formenreiche Gruppe (Chiappe 1995, 352) meist baumlebender Formen, die relativ plötzlich in großer Vielfalt in Schichten der Unterkreide auftauchen, aber am Ende der Kreide vollständig aus dem Fossilbericht verschwinden und vermutlich ausstarben. Dagegen lebten die gleichzeitig existenten Ornithurae in Wassernähe; diese Gruppe überlebte die Kreide-Tertiär-Grenze.
Archaeornithura besaß ein ziemlich modernes Federkleid mit asymmetrischen Schwungfedern und gut entwickelter Alula (Daumenfittich) mit mindestens drei Federn und fächerförmigen Schwanzfedern, die heutigen Vögeln erstaunlich ähnlich sind (Wang et al. 2015, 7). Beide Merkmale sind bei heutigen Vögeln für langsamen Flug und Manövrierbarkeit aerodynamisch wichtig. Archaeornithura dürfte daher ein guter Flieger gewesen sein. Die Knochen am Ende der Flügel sind stark verschmolzen und es ist eine U-förmige Furkula (verwachsenes Schlüsselbein) ausgebildet (Balter 2015) – beides Kennzeichen auch heutiger Vögel. Seine langen Beine waren nicht befiedert, was darauf hinweist, dass es sich um einen Watvogel gehandelt haben dürfte, der im flachen Wasser herum stakste und dort nach Nahrung suchte.
Archaeornithura gehört zu den Ornithurae und innerhalb dieser Gruppe zur Familie der Hongshanornithidae. Sein Alter wurde auf 130,7 Millionen (radiometrische) Jahre bestimmt, womit diese Gattung die älteste bisher bekannte ornithurine Form ist und das bisherige Höchstalter dieser Gruppe um ca. 5 Millionen (radiometrische) Jahre heraufsetzt. Man sollte erwarten, dass die älteste bekannte Form zu den eher primitiven Gattungen gehört, doch genau das Gegenteil ist der Fall. Archaeornithura ist im Cladogramm (Ähnlichkeitsbaum) der Gruppe tief eingeschachtelt (Wang et al. 2015, 6); die neue Gattung ist innerhalb der Hongshanornithidae eine spezialisierte Form, wobei diese Familie ihrerseits innerhalb der Ornithuromorpha ebenfalls tief eingeschachtelt ist, anders als andere Taxa aus jüngeren Schichten (entgegen der zu erwartenden Reihenfolge). Wang et al. (2015, 6) sprechen von Inkonsistenzen zwischen Stratigraphie und Phylogenie, was die Annahme sogenannter Geisterlinien erforderlich mache. Außerdem müsse angenommen werden, dass der Ursprung der Ornithurae in die Zeit zurück verlegt werden muss. „Geisterlinien“ bedeutet, dass aufgrund der Stammbaumrekonstruktion (phylogenetische Analyse, die auf Merkmalsvergleichen beruht) die Existenz evolutionärer Linien zwar angenommen werden muss, dass dafür aber in größerem Umfang fossile Nachweise fehlen. (Das gilt natürlich nur, falls Evolution abgelaufen ist.)
Kommentar
Sowohl die Ornithurae als auch die Enantiornithes erscheinen in der Fossilabfolge nicht in Form einer wachsenden Verschiedenartigkeit, sondern relativ ausdifferenziert. Man sagt in solchen Fällen, die evolutionäre Aufspaltung sei rasch erfolgt (Balter 2015). Dass es – unter der Annahme von Makroevolution – Lücken in der Fossilüberlieferung gibt, ist nachvollziehbar. Wenn solche Lücken aber systematischer Natur zu sein scheinen und in größerem Umfang Geisterlinien angenommen werden müssen, ist aufgrund der fossilen Daten ein kritische Rückfrage an das zugrundeliegende Evolutionsmodell angebracht. Das gilt erst recht, wenn Geisterlinien auch bei anderen Tier- und Pflanzengruppen angenommen werden müssen (vgl. dazu Frühe fossile Fährten rangieren Übergangsformen aus, https://www.genesisnet.info/index.php?News=141 zur Entstehung der Vierbeiner, Neuer Text über die kambrische Explosion, https://www.genesisnet.info/index.php?News=215 über die kambrische Explosion oder Ullrich (2015) zur Entstehung der Säugetiere).
Literatur
Balter M (2015) When modern birds took flight. Science 348, 617.
Chiappe LM (1995) The first 85 million years of avian evolution. Nature 379, 349-355.
Feduccia A (2012) Riddle of the feathered dragons. Hidden birds of China. Yale University Press. New Haven and London.
Padian K & Chiappe LM (1998) The origin an early evolution of birds. Biol. Rev. 73, 1-42.
Ullrich H (2015) Wann entstanden die modernen Säugetiere? Einsichten aus Fossilien, Molekülen und Datierungen. Stud. Integr. J. 22, 23-29.
Wang M (2015) et al. The oldest record of ornithuromorpha from the early cretaceous of China. Nat. Comm. 6:6987, doi: 10.1038/ncomms7987
Anmerkung
1 Weitere markante Unterschiede sind unter anderem: Die Knochen haben im Vergleich mit anderen Vögeln ein einzigartiges Muster in der histologischen Struktur; es gab zyklische Pausen beim Knochenwachstum (Chiappe 1995, 352f.); die Zähne sind nicht gezackt und habe eine eingeschnürte Basis (Padian & Chiappe 1998, 27). Außerdem unterschied sich die Flugarchitektur (Feduccia 2012, 111).
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n226.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
06.05.15 Yi qi – „merkwürdiger Flügel“ eines (Dino-?)sauriers
Ein seltsam gebauter mutmaßlicher Dinosaurier verwundert die Fachwelt. Die neu entdeckte Art wurde Yi qi genannt, das heißt „merkwürdiger Flügel“ in der Mandarin-Sprache. Merkwürdig ist vor allem ein langer stabförmiger Knochen, der vom Handgelenk ausgeht und an dem vermutlich eine Flughaut aufgespannt war (was aber nicht ganz gesichert ist). Der auch mit büscheligen haarartigen Anhängen („Federn“?) ausgestattete Yi qi vergrößert die Vielfalt der Formen im Verwandtschaftsumfeld von Dinosauriern und Vögeln, passt aber nicht gut in evolutionäre Szenarien.
Als „obskur und merkwürdig“ bezeichnet Kevin Padian (2015) ein neu beschriebenes Dinosaurier-Fossil, das in den weiteren Verwandtschaftskreis von Urvögeln gestellt wird. Es stammt aus Schichten aus dem Grenzbereich Mittel-/Oberjura der Tiaojishan-Formation in der Hebei-Provinz in China (Xu et al. 2015). Verwunderung löst vor allem eine lange, knöcherne Struktur aus, die vom Handgelenk ausgeht, und fossilisierte Reste membranartiger (häutiger) Strukturen im Bereich der Hand und der Vorderextremitäten. Für das lange, leicht gebogene und sich etwas zuspitzende Skelettelement ist keine Entsprechung bei irgendwelchen anderen Organismen bekannt. Es hat keine Gelenke und kann von seiner Lage her wahrscheinlich nicht als regulärer vierter Finger der Hand interpretiert werden.1 Wie die Verbindung mit dem Handgelenk erfolgte, ist unklar, daher kann auch nicht ermittelt werden, wie das stabförmige Element bewegt wurde und welche Lage es genau hatte. Die Forscher sind sich auch nicht sicher, ob es sich um einen Knochen oder einen kalzifizierten (verkalkten) Knorpel handelt. Ziemlich klar scheint aber zu sein, dass es sich um ein Stützelement für eine Flughaut gehandelt hat; dafür spricht auch die Kombination mit den erhaltenen Hautresten. Der eigenartige Knochenfortsatz dürfte als Aufspannsporn für eine Gleitflügelmembran fungiert haben; aufgrund der schlechten Erhaltung der mutmaßlichen Flughaut bleibt diese Deutung aber unsicher.
Darüber hinaus wurde an vielen Körperteilen eine haarartige Körperbedeckung nachgewiesen. Diese Körperanhänge werden von den Forschern zwar als „Federn“ bezeichnet, aber es handelt sich nur um extrem dünne, büschelige, pinselartige Fasern, die kaum eine aerodynamische Bedeutung haben konnten. Die Federnatur der Körperanhänge wird auch durch den mutmaßlichen Nachweis von Melanosomen sehr unterschiedlicher Größe begründet (Xu et al. 2015). Melanosomen sind Organelle, die Farbstoffe enthalten, für die Gefiederfärbung sorgen und als typisch für Federn gelten. Der Nachweis, dass es sich bei solchen rundlichen bis länglichen Gebilden bei Fossilien tatsächlich um Melanosomen handelt, ist allerdings schwierig und umstritten (Moyer et al. 2014, worauf Xu et al. selber hinweisen).
Die mutmaßliche Flughaut mit dem „Zusatzfinger“ spricht dafür, dass sich das Tier gleitend fortbewegen konnte; seltsamerweise passt aber der sonst bekannte Körperbau nicht so recht zu dieser Fortbewegungsweise, wie die Beschreiber Xu et al. (2015) bemerken. So sei der Körperschwerpunkt zu weit hinten, was zu einem Abkippen führe. Allerdings fehlen Überreste des Beckens und der meisten Teile der unteren Körperhälfte, deren Bau durch Vergleich mit ähnlichen Formen erschlossen wird (s. u.). Eine aktive Flugfähigkeit erscheint dagegen ausgeschlossen (Padian 2015). Der Dinosaurier mit der mutmaßlichen Flughaut ist also ein wirklich seltsames und rätselhaftes Tier, was sich auch in seiner Benennung niedergeschlagen hat: Yi qi – wohl der kürzeste Artname überhaupt – bedeutet in der Mandarin-Sprache „merkwürdiger Flügel“. Das Tier war ziemlich klein, nicht einmal taubengroß und es hatte kleinere und weniger Zähne als mit ihm verwandte2 Dinosaurier und als der bezahnte „Urvogel“ Archaeopteryx, der aus jüngeren Schichten des Oberjura stammt.

Abb. 1: Rekonstruktion von Yi qi mit seinen Membran-Flügeln. (Wikimedia: Emily Willoughby, (e.deinonychus@gmail.com, emilywilloughby.com), CC BY-SA 4.0)
Yi qi wird zur Gruppe des Scansoriopterygiden („kletternde Flügel“) gestellt, von der bisher drei Arten bekannt waren. Diese Gruppe ist auch ohne Yi qi schon ungewöhnlich, weil der dritte Finger der zu ihr gehörenden Arten deutlich der längste ist, während bei verwandten Formen (Dinosauriern und Vögeln) sonst immer der zweite Finger der längste ist. Außerdem sind die Arme ungewöhnlich lang und der Kopf ziemlich klein. Dies alles verbindet Yi qi mit dieser Gruppe, weshalb die Zuordnung gesichert scheint – der dritte Finger ist bei Yi qi besonders lang, womit er als zusätzliche Stütze für die mutmaßliche Flughaut gedient haben könnte. Aufgrund der Zuordnung zu den Scansoriopterygiden kann man vermuten, dass die bei Yi qi fossil nicht erhaltenen Körperteile ähnlich waren wie bei den anderen Mitgliedern dieser Gruppe (daraus wird auch der oben erwähnte Körperschwerpunkt ermittelt).
Die Scansoriopterygiden waren sehr wahrscheinlich kletternde Formen (Zhang et al. 2002) und befiedert; allerdings sind die Federn zu schlecht erhalten, um ihre aerodynamische Bedeutung sicher einschätzen zu können. Ihre taxonomische Stellung wird unterschiedlich bestimmt, mehrheitlich zu Theropoden (zweibeinige Raubdinosaurier) an der Basis der Avialae (Vögel und unmittelbar verwandte Formen), alternativ von einer Minderheit aber auch zu den Archosauriern3 ohne phylogenetischen Zusammenhang mit theropoden Dinosauriern (Zhang et al. 2002; Czerkas & Juan 2002; Feduccia 2012, 149ff.; Czerkas & Feduccia 2014; Xu et al. 2015).
Kommentar
Padian (2015) bezeichnet Yi qi als einen gefiederten Dinosaurier von einem völlig unerwarteten Zweig des Dinosaurier-Stammbaums. Da der berühmte „Urvogel“ Archaeopteryx aus dem Oberjura Federn eines modernen Typs besaß, werden Vorläuferformen in älteren Schichten gesucht. Gefunden wurde eine Reihe gefiederter Dinosaurier jedoch in jüngeren Schichten der Kreide-Formation, was stratigraphisch folglich nicht passt. Yi qi wäre von der Position in der stratigraphischen Tabelle her zwar als Vorläufer geeignet, aber diese eigenartige Form passt vom Körperbau überhaupt nicht und man hätte eine solche Form evolutionstheoretisch an der mutmaßlichen Basis der Vögel keinesfalls erwartet (Xu et al. 2015, 4).4 Einmal mehr taucht eine eher spezialisierte Form einer Gruppe relativ früh in der Schichtenabfolge auf. Und mutmaßliche Vorläuferformen müssen daher einmal mehr zurückverlegt werden.
Yi qi fällt unter den Dinosauriern und Vögeln der Kreidezeit völlig aus dem Rahmen.5 Statt Hinweise auf Vogelvorfahren zu geben erweitert Yi qi das Spektrum verschiedenartiger Formen aus Mittel-/Oberjura und Unterkreide, die kaum in evolutionäre Abfolgen zu bringen sind. Cladogramme (Ähnlichkeitsbäume) beinhalten entsprechend in großem Umfang Konvergenzen* und Reversionen**; so müsste bei Yi qi aufgrund seiner phylogenetischen Position angenommen werden, dass es sich bei seinen einfachen federartigen Körperanhängen um rückgebildete Strukturen handelt. Czerkas & Feduccia (2014) halten die Scansoriopterygiden gar nicht für Dinosaurier und sind – als allerdings viel kritisierte Minderheit – der Auffassung, dass die Evolution der Vögel nicht über Dinosaurier, sondern über phylogenetisch ältere Archosaurier verlief. Sie stellen die kletternden Scansoriopterygiden an die Basis der Vögel und sehen sie als Beleg dafür, dass der Vogelflug nicht von laufenden Theropoden-Dinosauriern, sondern von kletternden Formen ausging. Doch der neue Fund Yi qi mit seinem eigentümlichen Körperbau steht auch quer zu diesem Szenario.
*Konvergenzen (= unabhängige Entstehung baugleicher Merkmale)
** Reversionen (= Rückentwicklungen)
Die Bearbeiter Xu et al. (2015, 1, 4) interpretieren Yi qi als Beispiel eines evolutionären Experiments – ein Deutungsversuch, der in der Wissenschaftspresse aufgegriffen wurde und auch in vielen vergleichbaren Situationen bei anderen Tier- und Pflanzengruppen zur Anwendung kommt. „Experiment“ ist aber eigentlich ein evolutionstheoretisches Unwort, da dieser Begriff eine Zielsetzung impliziert, ein Ausprobieren, um gute Lösungen zu finden. Doch nach dem evolutionären Ansatz gibt es weder einen Experimentator noch irgendeine Zielorientierung. Daher verschleiert der im Originalartikel verwendete Begriff „experimentation“ eher ein evolutionstheoretisches Problem, als dass er irgendeine Erklärung beinhaltet.
Xu et al. (2015, 4) merken an, dass es sich bei der mutmaßlichen Flughaut, die an dem langen „Zusatzfinger“ aufgespannt ist, um ein eindrucksvolles Beispiel einer Konvergenz handelt, der Bau sei mit den Flugsauriern vergleichbar, die einen einzigen stark verlängerten Finger zum Aufspannen der Flughaut besaßen, aber die Ähnlichkeit sei nur oberflächlich. Eine genauere Entsprechung liege auch zu anderen gleitenden Tieren nicht vor. Ohnehin muss angenommen werden, dass die Fähigkeit zum Gleitflug in verschiedenen Wirbeltiergruppen mehrfach unabhängig entstanden ist.
Insgesamt zeigt sich einmal mehr, dass die vielzitierte Auffassung des Evolutionsbiologen Theodosius Dobzhansky, nichts ergebe Sinn außer im Licht der Evolution, fragwürdig ist. Evolutionäre Zusammenhänge liegen im Falle von Yi qi nicht ohne weiteres auf der Hand; das neue Fossil wirft für evolutionäre Rekonstruktionen mehr neue Fragen auf, als dass es alte beantworten würde.
Anzumerken ist noch, dass der genaue Fundort nicht bekannt ist; das Fossil wurde nicht durch die Paläontologen in situ* geborgen, sondern von einem Bauern erworben. Die Autoren haben sich aber große Mühe gegeben, den Fundort zu rekonstruieren und sicherzustellen, dass es sich nicht um eine Fälschung handeln kann.
*in situ (= an Ort und Stelle, am ursprünglichen Fundort)
Literatur
Czerkas SA & Feduccia A (2014) Jurassic archosaur is a non-dinosaurian bird. J. Orn. 155, 841-851.
Czerkas SA & Yuan C (2002) An arboreal maniraptoran from Northeast China. The Dinosaur Museum, Blanding, Utah. http://www.dinosaur-museum.org/featheredinosaurs/arboreal_maniraptoran.pdf.
Feduccia A (2012) The riddle of the feathered dragons. New Haven & London: Yale Univ. Press.
Moyer AE et al. (2014) Melanosomes or microbes: testing an alternative hypothesis for the origin of microbodies in fossil feathers. Sci. Rep. 4, 4233.
Padian K (2015) Dinosaur up in the air. Nature 521, 40-41, doi:10.1038/nature14392.
Xu X, Zheng X, Sullivan C, Wang X, Xing L, Wang Y, Zhang X, O’Connor JK, Zhang F & Pan Y (2015) A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings. Nature 521, 70-73, doi: 10.1038/nature14423.
Zhang F, Zhou Z, Xu X & Wang X (2002) A juvenile coelurosaurian tetrapod from China indicates arboreal habits. Naturwissenschaften 89, 394-398.
Anmerkungen
1 „… the styliform element of Yi qi has no joints and comes directly off the carpal bones without the intermediary of a metacarpal (palm bone), so it is probably not a finger“ (Padian 2015).
2 mit „verwandt“ ist hier nicht im deutenden Sinne „abstammungsverwandt” gemeint, sondern im beschreibenden Sinne „im Körperbau insgesamt ähnlich“.
3 ab dem Oberperm fossil überlieferte Gruppe, die u.a. Krokodile, Dinosaurier, Flugsaurier und Vögel umfasst.
4 „… the evident occurrence in this taxon of a membranous wing supported by a styliform element represents an unexpected aerodynamic innovation close to the origin of birds …“ (Xu et al. 2015, 4).
5 „… that Yi had membranous aerodynamic surfaces totally different from the archetypal feathered wings of birds and their closest relatives. … Indeed, no equivalent of the rod-like bone is known in any other dinosaur even outside Theropoda, but similar structures are present in a diverse array of extant and extinct flying or gliding” (Xu et al.)
Autor dieser News: Reinhard Junker
© 2015, http://www.genesisnet.info/schoepfung_evolution/n225.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
24.07.14 Immer rätselhafter: Der „Urvogel“ Archaeopteryx
Im Jahr 2011 wurde das elfte fossile Exemplar des „Urvogels“ Archaeopteryx aus einer Privatsammlung bekannt und seither wissenschaftlich untersucht. Kürzlich wurden Details in einem Nature-Artikel genauer beschrieben (Foth et al. 2014). Erstmals ist das komplette Gefieder erhalten, auch die Befiederung von Rumpf, Beinen und Schwanz. Ein Vergleich mit anderen gefiederten Fossilformen aus Oberjura und Unterkreide lässt die Wissenschaftler schließen, dass Federn ursprünglich nicht zum Fliegen entstanden sind, sondern andere Aufgaben hatten. Erst später seien sie zusätzlich zum Fliegen verwendet worden und das wahrscheinlich mehrmals unabhängig. Die frühen Vögel und gefiederten Theropoden-Dinosaurier zeigen eine komplexe und verworrene Vielfalt unterschiedlicher Mosaike, deren Anordnung in einem Cladogramm (= Ähnlichkeitsbaum) nicht mit den zeitlichen (stratigraphischen) Positionen der betreffenden Gattungen zusammenpasst.
Vor etwa 20 Jahren war der paläontologische* (Befund im hypothetischen evolutionären Übergangsbereich zwischen Reptilien und Vögeln noch relativ überschaubar. Der berühmte „Urvogel“ Archaeopteryx aus dem Oberjura, auf etwa 150 Millionen radiometrische Jahre datiert, stand dort als Vogel recht verloren da. Doch er galt schon immer als guter Beleg für einen evolutionären Übergang von zweibeinigen Raubdinosauriern zu Vögeln. Mit ersteren verbindet ihn eine Reihe morphologischer Merkmale wie z. B. ein bezahnter Kiefer oder ein langer Wirbelschwanz. Die Verbindung mit Vögeln war vor allem durch den Besitz von Federn begründet, die lange Zeit als typisch für flugtaugliche Federn heutiger Vögel galten.
*paläontologisch (= Fossilien betreffend)
Diese Situation hat sich in den letzten beiden Jahrzehnten grundlegend gewandelt. Vor allem in der Unterkreide Chinas wurden zahlreiche Fossilien befiederter Dinosaurier und eindeutiger Vögel wie z. B. Confuciusornis gefunden. Die Federnatur der Körperbedeckungen von Dinosauriern wurde allerdings teilweise in Frage gestellt (z. B. Feduccia et al. 2005), und bei einigen dieser Formen dürfte es sich um sekundär flugunfähige Arten handeln. Alle diese Formen hatten allerdings – in evolutionstheoretischer Perspektive – den „Makel“, dass sie deutlich jünger waren als Archaeopteryx, der vorerst das älteste Federtier blieb.
In den letzten Jahren hat sich auch das geändert, denn es wurden z. B. mit Haplocheirus (Choiniere et al. 2010, 571) eine möglicherweise und mit Anchiornis (Hu et al. 2009) eine eindeutig befiederte Gattung entdeckt, die älter als Archaeopteryx datiert wurden (vgl. Löst Haplocheirus das Zeit-Paradox der Vogelevolution?, https://www.genesisnet.info/index.php?News=143 und Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137). Im Jahr 2011 machte dann die neu entdeckte Gattung Xiaotingia in Bezug auf Archaeopteryx von sich reden, denn sie schien den „Urvogel“ aus der Linie, die zu den Vögeln führt, abzudrängen (Xu et al. 2011), was später revidiert wurde bzw. sich evolutionstheoretisch vorerst als nicht klar entscheidbar herausstellte (Lee & Worthy 2011; vgl. Fällt eine Ikone vom Sockel?, https://www.genesisnet.info/index.php?News=174). Es wurde also richtig gesellig um Archaeopteryx; der „Urvogel“ ist heute sowohl in morphologischer Hinsicht als auch hinsichtlich seiner zeitlichen Stellung nur noch einer unter vielen und seine Position als erster Vogel ist dadurch fraglich geworden.
Darüber hinaus kamen Zweifel an der Flugfähigkeit von Archaeopteryx auf. Die Federschäfte seien dafür zu schwach gewesen (Nudds & Dyke 2010). Eine Analyse von Longrich et al. (2012) zeigte, dass bei Archaeopteryx (wie auch bei der befiederten Gattung Anchiornis) eine bisher unerkannte und überraschende Anordnung von mehreren stark überlappenden Lagen dünner Federn vorlag. Moderne und fast alle fossil bekannten Vögel besitzen dagegen nur zwei Federlagen, die sich nur teilweise überlappen. Die Forscher schlossen aus den Gefiedermerkmalen auf eine eingeschränkte Flugfähigkeit bei Archaeopteryx (während Anchiornis ganz flugunfähig gewesen sein soll). (vgl. Archaeopteryx – Gleitflieger und Bindeglied?, https://www.genesisnet.info/index.php?News=193)
Das elfte Archaeopteryx-Exemplar. Nun wurden jüngst Untersuchungen an einem erst im Jahr 2011 bekannt gewordenen 11. Exemplar von Archaeopteryx veröffentlicht (Foth et al. 2014), das im Besitz eines privaten Sammlers ist. Dieses Exemplar weist einige Merkmale auf, die bisher nicht bekannt waren und das Bild von Archaeopteryx erneut verändern. Es ist das erste Exemplar, bei dem das komplette Gefieder fossile Spuren hinterlassen hat, so dass nun mehr als bisher oder überhaupt etwas über das Federkleid des Rumpfes, der Beine und des Schwanzes bekannt ist. Zudem sind die Details der Federstrukturen gut erhalten. Der Fund belegt, dass der ganze Körper mit Konturfedern bedeckt war, wie sie bei heutigen Vögeln bekannt sind. Der Federschaft war leicht gebogen. Die Form der Flügel entspricht der heutiger Vögel. Die bis zu 4,5 Zentimeter langen Deckfedern an den Beinen sind als so genannte Federhose ausgebildet und erinnern damit an die Ausprägung bei rezenten Raubvögeln. Sie ziehen in parallelen Reihen von den Oberschenkeln bis zum Knöchel. Die hinteren Schwanzfedern sind mit über 10 cm Länge extrem lang, ebenso die Federn an den Seiten des leicht gegabelten Schwanzes und zeigen eine aerodynamische Form, was neben den anderen genannten Gefiedermerkmalen für die Flugfähigkeit spricht. Auch die Federhose könnte den Nebeneffekt einer vertikalen Gleithilfe beim Landen gehabt haben, ähnlich heutigen Greifvögeln (http://www.scinexx.de/wissen-aktuell-17738-2014-07-03.html), könnte aber auch einen Schutz vor Bissen gewährt haben.
Die Schwungfedern weisen kräftige Schäfte auf. Foth et al. (2014, 80) halten daher die oben erwähnte Deutung von Nudds & Dyke (2010) und Longrich et al. (2012), die Federn seien angesichts eines schwach entwickelten Federschaftes kaum flugtauglich gewesen, für falsch und möglicherweise auf schlechte Erhaltung der von ihnen untersuchten fossilen Federabdrücke zurückzuführen.
Ursprüngliche Funktion von Federn? Die Forscher verglichen die Befiederung von Archaeopteryx mit dem Federkleid anderer früher fossiler Formen. Welche Federtypen kommen bei den einzelnen Gattungen in welchen Körperregionen vor? Es zeigte sich, dass auch bei flugunfähigen Formen eine große Variation von Federtypen und Befiederung vorliegt. Daraus schließen die Wissenschaftler, dass die Federn ursprünglich nicht zum Fliegen, sondern in anderen funktionellen Zusammenhängen entstanden sind. Die Vielfalt von Federn und der Befiederung spreche gegen eine anfängliche Selektion auf Flugfähigkeit, da diese Variationen sonst vermutlich eingeschränkt worden wären.
Die Frage nach der anfänglichen Funktion in der hypothetischen Federevolution wird schon lange diskutiert. Eine Auslese direkt auf Flugtauglichkeit wird nämlich als schwierig angesehen, weil dabei – salopp gesagt – zu viel auf einmal gegenüber Vorläuferstrukturen geändert werden müsste. Zwischenschritte auf dem Weg zu flugtauglichen Federn und einem flugtauglichen Federkleid sind also gefragt. Foth et al. (2014, 81) vermuten, dass das Federkleid anfangs im Zusammenhang mit der Balz evolvierte. Federn können aber auch zur Wärmeisolation genutzt werden, sie helfen außerdem beim Brüten und können zur Tarnung eingesetzt werden. Archaeopteryx und andere Arten nutzten die Armschwingen vielleicht, so die Autoren weiter, als Balancehilfe beim schnellen Laufen (wie das bei heute sekundär flugunfähigen Vögeln vorkommt). Der Umweg dieser Federn oder federartigen Strukturen, die primär die o.g. Aufgaben erfüllen, erscheint den Autoren eher gangbar als der direkte Weg zur Flugtauglichkeit.
Funktionswechsel hin zur Flugtauglichkeit? Die Vorstellung, dass Federn in der Evolution zunächst gar nichts mit dem Flug zu tun hatten und erst im Laufe der Zeit gleichsam zweckentfremdet wurden (Foth et al. sprechen von Exaptation), wirft jedoch andere Fragen auf. Zum einen stellt sich die Frage nach den Mechanismen der Federentstehung auch dann, wenn die Federn anfangs nichts zur Flugfunktion beigetragen haben sollten. Für die Funktionen Wärmeisolation, Balz oder Schutz werden des Weiteren keine Federn benötigt, dafür genügen einfachere Strukturen, und es ist nicht klar, weshalb bei Selektion auf diese Funktionen überhaupt Federn in einer Ausprägung entstehen sollten, die später nebenbei auch Flugtauglichkeit garantieren. Außerdem würde Selektion auf Wärmeisolation oder auf andere Funktionen hin dazu führen, dass diese Funktionen verbessert werden, nicht aber die andere Funktion der Flugtauglichkeit. Diese wird bei Selektion z. B. auf Wärmeisolation eher verschlechtert, weil sich die jeweiligen Erfordernisse an die Federstruktur widersprechen. Dass unter diesen Umständen plötzlich durch einen glücklichen Umstand die Federn auch zum Fliegen genutzt werden konnten, ist sehr unwahrscheinlich. Dafür brauchen die Federn besondere Eigenschaften wie z. B. Steifheit und aerodynamische Form. Es kommt noch hinzu, dass fürs Fliegen oder Gleiten nicht nur passend strukturierte Federn benötigt werden, sondern auch weitere konstruktive Neuheiten und physiologische Anpassungen wie eine geeignete Muskulatur und Körperform, ein leistungsfähiges Atmungs- und Kreislaufsystem, spezialisierte Sinnesorgane, Leichtbau der Knochen, entsprechende Verhaltensweisen u. v. m. Diese Aspekte und die Problematik gegenläufiger Selektionsdrücke je nachdem, welche Funktion der Federn optimiert werden soll, müssen bedacht werden. Vergleichende Studien fossiler und rezenter Vögel tragen bei diesen Fragestellungen nach den Mechanismen der Entstehung des Vogelfluges nichts zum Verständnis bei.
Mehrfache Entstehung des Vogelflugs? Der Vergleich des Vorkommens verschiedener Federtypen und der Befiederung bei verschiedenen Gattungen bringt unter evolutionstheoretischen Voraussetzungen ein problematisches Ergebnis: Werden die verglichenen Gattungen in ein Cladogramm (= Ähnlichkeitsbaum) gebracht und wird dieses stammesgeschichtlich interpretiert, muss angenommen werden, dass der Übergang von Federtypen, die z. B. zur Wärmeisolation oder als Schmuck verwendet wurden, zu flugtauglichen Federn mehrfach unabhängig erfolgt sein müsste (Foth et al. 2014; Details dort auf S. 80f.). Ein solches Szenario hätte man früher ausgeschlossen, denn die oben angedeuteten Probleme für einen solchen Übergang vermehren sich nicht nur, sondern vergrößern sich: Denn man muss sich immer wieder vor Augen halten, dass Evolution, im bisher verstandenen Sinne, völlig richtungslos verläuft und keine wie auch immer geartete Zielorientierung beinhaltet. Beispielsweise ist zu bedenken, dass ein flugunfähiges Tier weder die fürs Fliegen erforderlichen morphologischen und physiologischen Begleitmerkmale besitzt noch die geringste „Ahnung“ vom Flugverhalten hat (die fürs Fliegen erforderlichen Verhaltensweisen). Wie kann mehrfach auf der Basis richtungsloser Änderungen (Mutationen) und Auslese auf Verbesserung aktueller (nicht zukünftiger) Funktionen (z. B. Wärmeisolation oder Balz) dasselbe, primär nicht durch eine Selektion bewertete „Ergebnis“ in Form der Flugfedern erreicht werden, deren Herausbildung offenkundig viele tiefgreifende Änderungsschritte erfordern?
Die Forscher kommentieren diese Situation damit, dass der Ursprung und die Evolution des Vogelflugs komplexer ablief als zuvor gedacht und nicht auf linearen Wegen. Die damit verknüpfte Frage nach den Selektionsdrücken wird im Artikel von Foth et al. (2014) nicht aufgeworfen.
Rätselhaftes Merkmalsmosaik. Insgesamt erscheint das Merkmalsmosaik Archaeopteryx immer rätselhafter und widersprüchlicher: Einige Merkmale weisen auf Flugtauglichkeit hin, andere passen nicht gut dazu. Man kann daraus wohl schließen, dass die Biologie des „Urvogels“ zu wenig verstanden ist. Das ist Grund genug, Vorsicht bezüglich evolutionstheoretischer Schlussfolgerungen walten zu lassen. Hat Archaeopteryx seine Flugfähigkeit vielleicht teilweise eingebüßt? Diese Hypothese brachte Michael Habib (2013) von der University of Southern California beim Jahrestreffen der Society of Vertebrate Paleontology in Los Angeles ins Spiel (Kaplan 2013). Er schließt dies aus dem Vergleich von Beinlängen und Federsymmetrien bei Archaeopteryx und heutigen Vögeln; diese seien bei Archaeopteryx ähnlich wie bei heutigen sekundär flugunfähigen Vögeln. Natürlich ist diese Hypothese umstritten, und der Biologe Ahsley Heers stellt fest, dass ein besseres Verständnis des Zusammenhangs von Anatomie und Lebensweise erreicht werden müsse, um den Fossilbericht besser interpretieren zu können; außerdem müsse berücksichtigt werden, dass viele heutige Vögel im Jugendstadium besser fliegen könnten als im erwachsenen Zustand (Kaplan 2013).
Schlussfolgerungen bezüglich Evolution und Schöpfung. Die Fossilüberlieferung früher Vögel hat sich in den letzten 20 Jahren grundlegend verbessert. Die zuvor noch relativ „einsame“ Ikone Archaeopteryx findet sich heute in einem bunten Mosaik von Formen mit verschiedensten Federkleidern, seien es Vögel, die zum Teil sekundär flugunfähig sind, oder befiederte Dinosaurier. Kandidaten für evolutionäre Übergangsformen in Bezug auf Einzelmerkmale gibt es durchaus. Paradoxerweise erlaubt aber gerade die mosaikartige Merkmalsvielfalt der neu entdeckten Tiere keine Benennung von Übergangsformen auf der Ebene der Arten oder Gattungen. Die Formenvielfalt führt bei evolutionstheoretischer Deutung zu einer widersprüchlichen Verteilung von Einzelmerkmalen im Stammbaum, was die Annahme zahlreicher Konvergenzen* erfordert. Auch der Übergang zum Flug wird mittlerweile nicht mehr unbedingt als ein einmaliges evolutionäres Ereignis betrachtet. Warum dies problematisch ist, wurde oben erläutert.
*Konvergenzen (= unabhängig entstandene ähnliche Merkmale)
Foth et al. (2014) veröffentlichen ein Cladogramm der Maniraptoren-Theropoden (Fig. 3 ihres Artikels), in dem die verschiedenen Federmerkmale der einzelnen Gattungen bzw. höherer Taxa eingetragen sind. (Dabei zeigen sich die oben erwähnten Konvergenzen.) Trägt man in dieses Cladogramm auch die Zeitpunkte des fossilen Auftretens der betreffenden Formen ein, ergibt sich, dass sehr viele Vertreter von vermeintlichen Vorläuferlinien paradoxerweise deutlich später als die Vertreter der vermeintlichen Nachfahrenlinien erscheinen. Es gibt in diesem Bereich keinerlei Tendenz einer Parallelität zwischen dem zeitlichem Auftreten eines Fossils und seiner morphologischen Komplexität. Das heißt: Bereits im Oberjura und in der Unterkreide erscheint insgesamt eine sehr bunte Vielfalt unterschiedlichster Mosaikformen von flugfähigen und (evtl. sekundär) flugunfähigen Formen. Das zeigt beispielhaft, dass ein Cladogramm nicht ohne weiteres stammesgeschichtlich gedeutet werden kann bzw. dass dies nur möglich ist, wenn in großem Umfang sogenannte Geisterlinien angenommen werden. (Von Geisterlinien spricht man, wenn die Merkmalsverteilung in evolutionstheoretischer Deutung die Annahme längerer Zeiträume erfordert, in denen die betreffenden Formen existiert haben müssten, diese aber fossil nicht belegt sind.)
Wie im Artikel Fällt eine Ikone vom Sockel?, https://www.genesisnet.info/index.php?News=174 angemerkt, kann man in einer schöpfungsorientierten Perspektive den Befund häufiger Mosaikformen, die zudem bezüglich ihres zeitlichen Nachweises nicht in Abstammungsfolgen hineinpassen, als Ausdruck eines designerspezifischen Baukastensystems werten. Merkmale sind dann beliebig kombinierbar, sofern der Gesamtkomplex eine lebensfähige Art ergibt. Das Nebeneinander verschiedener Mosaikformen und das verbreitete Fehlen einer zeitlichen Passung ist aus dieser Perspektive kein grundsätzliches Problem. Für die biblische Schöpfungslehre, sofern sie von einer kurzen Erdgeschichte ausgeht, bleibt aber die Frage nach den Ursachen der stratigraphisch geordneten Abfolgen größerer Gruppen: Warum ist die Vogelwelt in den Ablagerungen des Jura und in der Kreide eine ganz andere als die des Tertiärs?
Literatur
Choiniere JN, Xu X, Clark JM, Forster CA, Guo Y & Han F (2010) A Basal Alvarezsauroid Theropod from the Early Late Jurassic of Xinjiang, China. Science 327, 571-574.
Feduccia A, Lingham-Soliar T & Hinchliffe JR (2005) Do feathered dinosaurs sxist? Testing the hypothesis on neontological and paleontological evidence. J. Morphol. 266, 125-166.
Foth C, Tischlinger H & Rauhut OWM (2014) New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature 511, 79-82.
Hu D, Hou L, Zhang L & Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 460-463.
Kaplan M (2013) Theory suggests iconic early bird lost its flight. Nature, doi:10.1038/nature.2013.14142
Lee MSY & Worthy TH (2011) Likelihood reinstates Archaeopteryx as a primitive bird. Biol. Lett. doi: 10.1098/rsbl.2011.0884
Longrich NR, Vinther J, Meng Q, Li Q & Russell AP (2012) Primitive Wing Feather Arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Curr. Biol. 22, 2262-2267.
Nudds RL & Dyke GJ (2010a) Narrow primary feather rachises in Confuciusornis and Archaeopteryx suggest poor flight ability. Science 328, 887-889.
Xu X, You H, Du K & Han F (2011) An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 475, 465-470.
Überblick über weitere Genesisnet-Newsbeiträge zu Urvögeln:
- Löst Haplocheirus das Zeit-Paradox der Vogelevolution?, https://www.genesisnet.info/index.php?News=143
- Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137
- Fällt eine Ikone vom Sockel?, https://www.genesisnet.info/index.php?News=174
- Archaeopteryx – Gleitflieger und Bindeglied?, https://www.genesisnet.info/index.php?News=193
Anmerkungen
1 Bei Archaeopteryx und Anchiornis seien wegen der schlanken Federschäfte die einzelnen Federn zu schwach für eine Flugtauglichkeit gewesen, nur ihre ungewöhnliche Zusammenlagerung zu mehreren Lagen habe eine dicke, robuste Tragfläche ermöglicht. Allerdings dürfte weder das Spreizen noch das Schrägstellen der Schwungfedern bei den Flügeln möglich gewesen sein, so dass Archaeopteryx sich möglicherweise nur gleitend durch die Lüfte bewegen konnte, während Anchiornis flugunfähig war.
2 „‘Wären Federn primär für das Flugvermögen entstanden, dann hätte das die Variation aus funktionalen Gründen vermutlich eingeschränkt. So sehen wir in den Flügeln früherer Vögel weniger Variation als in den Hinterbeinen oder am Schwanz‘, erklärt Foth“ (http://www.uni-muenchen.de/informationen_fuer/presse/presseinformationen/2014/rauhut_archaeopteryx.html).
Autor dieser News: Reinhard Junker
© 2014, http://www.genesisnet.info/schoepfung_evolution/n216.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
29.01.13 Archaeopteryx – Gleitflieger und Bindeglied?
Eine neue Analyse des Federkleids des berühmten „Urvogels“ Archaeopteryx und des befiederten Dinosauriers Anchiornis weist bei diesen beiden Gattungen eine bisher unerkannte und überraschende Anordnung von mehreren Lagen dünner Federn auf. Die Forscher schließen daraus auf eine eingeschränkte Flugfähigkeit bei Archaeopteryx, während Anchiornis flugunfähig gewesen sein dürfte. Diese Befunde eigenen sich jedoch aus verschiedenen Gründen nicht als Indizien dafür, dass es sich um Übergangs-Merkmalsausprägungen handelt. Das gesamte Merkmalsspektrum lässt sich besser verstehen, wenn man die beiden Gattungen als spezialisierte Formen betrachtet. Aufgrund der nicht sicher bestimmbaren Struktur-Funktions-Beziehungen müssen weitergehende Schlussfolgerungen unsicher bleiben.
Man könnte meinen, dass über Archaeopteryx, den berühmten Urvogel aus dem Oberjura bereits alles gesagt ist. Schließlich konnte er 150 Jahre lang untersucht werden und es erschienen zahllose Artikel über diesen „komischen Vogel“. Dennoch konnte ein Forscherteam um Nicholas R. Longrich von der Yale University New Haven (USA) kürzlich neue überraschende Befunden der Öffentlichkeit vorstellen (Longrich et al. 2012). Bisher war angenommen worden, dass das Federkleid von Archaeopteryx ähnlich angeordnet war wie das heutiger Vögel. Eine erneute genaue Analyse der Lage der Federn zeigte jedoch, dass die bisherige Rekonstruktion des Federkleids teilweise auf einem Deutungsfehler der fossilen Federabdrücke beruhte. Es zeigte sich, dass Archaeopteryx und der ebenfalls untersuchte befiederte Dinosaurier Anchiornis huxleyi mehrere Lagen von dünnen, stark überlappenden Federn besitzen und nicht wie moderne und fast alle fossil bekannten Vögel nur zwei Lagen, die sich nur teilweise überlappen. Heutige Vögel (Neornithes) besitzen über den langen, asymmetrischen Flugfedern kurze Deckfedern. Diese gestaffelte Struktur macht es den Vögeln möglich, „die Schwungfedern gezielt auseinander zu spreizen, um bei langsamem Flug mehr Auftrieb zu erzeugen. Zudem können die heutigen Vögel ihre Schwungfedern schräg stellen, sodass beim Aufwärtsschwingen der Flügel Lücken entstehen. Dadurch kann Luft hindurchfließen, und der Kraftaufwand vor allem beim flatternden Langsamflug reduziert sich“ (http://www.zeit.de/wissen/2012-11/archaeopteryx-fliegen-federn-gleiten).
Aus ihren Befunden schließen Longrich und Mitarbeiter, dass aufgrund der schlanken Federschäfte von Archaeopteryx und Anchiornis zwar die einzelnen Federn schwach waren; ihre Zusammenlagerung zu mehreren Lagen aber eine dicke, robuste Tragfläche ermöglicht habe. Allerdings dürfte weder das Spreizen noch das Schrägstellen der Schwungfedern bei den Flügeln der beiden Gattungen möglich gewesen sein. Die Folgen waren sehr wahrscheinlich ein erschwerter Start vom Boden aus und Einschränkungen beim langsamen Flug. Die Vielfalt der Funktionen heutiger Flügel dürfte also gefehlt haben, so dass die Flugfähigkeit insgesamt eingeschränkt gewesen sein muss. Möglicherweise konnte Archaeopteryx sich nur gleitend durch die Lüfte bewegen. Aus evolutionstheoretischer Sicht würde das dafür sprechen, dass die ersten gefiederten Dinosaurier und Vögel von Bäumen aus durch die Luft glitten (Arborealtheorie), während der Flugerwerb durch schnelles Laufen vom Boden aus nicht in Frage käme (Cursorialtheorie).
Bei Anchiornis ist die Situation ähnlich wie bei Archaeopteryx, kann aber insofern als „primitiver“ gewertet werden, als diese Gattung einfache, bandartige Federn besaß, und zwar in großer Zahl und dichter Überlappung. Eine ähnliche Situation sei unter den heutigen Vögeln nur bei Pinguinen bekannt, wird Longrich in Pressemeldung zitiert. Bei Anchiornis war schon früher angenommen worden, dass er flugunfähig war, was allerdings einige Fragen aufwirft (s. u.).
Weitere Untersuchungen. Schon vor einigen Jahren war eine Analyse der Federschäfte der Handschwingen von Archaeopteryx veröffentlicht worden, wonach – verglichen mit heute lebenden Vogelarten – die Schäfte deutlich zu schwach für einen kontinuierlichen Ruderflug waren (Nudds & Dyke 2010a), obwohl die Länge der Handfedern der Größe bei vergleichbar großen heute lebenden Vögeln entspricht. Die Forscher schließen daraus, dass die untersuchten fossilen Vögel nur kurze Strecken gleiten, aber nicht aktiv fliegen konnten. Sonderbar ist aber, dass der Bau der Flügel im Ganzen nicht zum längeren Gleiten passt (Nudds & Dyke 2010a, 889), obwohl weitere anatomische Befunde (z. B. Anatomie des Schultergürtels, Federinnervation) bei Archaeopteryx dafür sprechen. An dieser Schlussfolgerung wurde kritisiert, dass Nudds & Dyke (2010a) falsche Körpermassen zugrundegelegt hätten, und dass nicht berücksichtigt worden sei, dass viele Merkmale von Archaeopteryx für die Fähigkeit zum aktiven Flug sprechen (Paul 2010; vgl. auch die Kritik von Longrich et al. 2012, 2264, aber auch die Antwort von Nudds & Dyke [2012b]). Anfang 2012 wurde dann eine Untersuchung veröffentlicht, wonach die Federn von Archaeopteryx wie die Federn moderner Vögel Melanosomen (Strukturen in farbstoffproduzierenden Zellen) besaßen, die zur Festigkeit und Haltbarkeit der Federn beitragen (Carney et al. 2004), was von den Forschern als weiteres Indiz dafür gewertet wurde, dass der Urvogel aktiv fliegen konnte. Carney et al. (2012) zeigten außerdem, dass die Struktur der Federn mit der von heutigen Vögeln identisch war.
Das Problem der fehlenden Analogie. Offenbar ergeben die Merkmale von Archaeopteryx in Bezug auf seine Flugfähigkeit also kein einheitliches Bild. Woran liegt das? Es ist gut möglich, dass die Struktur-Funktions-Beziehungen nicht genügend aufgeklärt sind und vielleicht sogar im Dunklen blieben müssen, weil wir unter den heutigen Arten keine Analogie für das gesamte Merkmalsspektrum haben. Dieser Umstand sollte vor vorschnellen evolutionären Interpretationen warnen, bei denen mit „Primitivität“ argumentiert wird. So könnte man zwar auf den ersten Blick Anchiornis und Archaeopteryx als Übergangsformen interpretieren, insbesondere wenn man einzelne Merkmale wie den Aufbau des Federkleids zugrundelegt. Dem stehen aber zwei Probleme entgegen.
Die erste Schwierigkeit ist das bereits erwähnte Fehlen eines heute lebenden Vorbilds für die Merkmalskombination von Archaeopteryx und die scheinbare (?) Widersprüchlichkeit der Daten, insbesondere bei Anchiornis. Die Schädelanatomie und die davon abgeleiteten neurologischen Fähigkeiten und andere oben bereits erwähnte Merkmale sprechen für die Fähigkeit zum aktiven Flug (Alonso et al. 2004), während Federstruktur und Art des Federkleids dieser Deutung entgegenstehen (s. o.). Eine Einschätzung der Lebensweise und Ökologie aufgrund des gesamten Merkmalsmosaiks ist daher unsicher. Von dieser Einschätzung hängt aber auch eine evolutionstheoretische Interpretation ab, die daher ebenfalls unsicher bleiben muss. Die ungewöhnliche Merkmalskombination deutet darauf hin, dass die beiden Gattungen spezialisierte Formen mit im Einzelnen nicht sicher rekonstruierbarer Lebensweise sind.
Offen bleiben auch die Fragen nach Selektionsdrücken und Umbaumechanismen. Wenn dünne Federn nur im Verband eine gewisse Flugfähigkeit (inklusive Gleitflug) ermöglichen, müssen sie auch im Verband vorliegen, damit sie einen Selektionswert im Hinblick darauf haben. Zwar wird angenommen, dass Federn zunächst andere Funktionen wie Wärmeregulation oder als Signalgeber hatten, aber eine selektionsgesteuerte Veränderung auf diese Funktionen hin steht teilweise im Widerspruch zur Selektion auf Flugfähigkeit und würde eine Veränderung in Richtung auf Flugtauglichkeit eher verhindern.
Erst recht problematisch bezüglich des Verständnisses der Lebensweise ist Anchiornis. (Vergleiche dazu Vierflügelige Vögel am Anfang?, https://www.genesisnet.info/index.php?News=137) Diese Gattung, von der nahezu vollständig erhaltene Fossilfunde bekannt sind, hatte sehr lange Unterschenkel, was auf eine laufende Lebensweise hinweist; dazu passen aber das lange und umfassende Federkleid und die Befiederung der Beine nicht, da diese beim Laufen eher hinderlich wären. Federn können eine Vielfalt verschiedener Funktionen ausüben (Stettenheim 2000). Wozu also hatte diese Gattung ein ausgebildetes Federkleid mit Konturfedern an Arm- und Handschwingen wie bei modernen Vögeln und war dennoch möglicherweise flugunfähig? (Hu et al. 2009) Der Bau der Federn und des Federkleides als Ganzes sowie der lange Steuerschwanz sprechen am ehesten für eine gemischte Boden-Baum-Lebensweise als Gleitflieger. Weitere Federmerkmale könnten Balz- und Territorialfunktion gehabt haben. Ein ausdauernder Läufer kann Anchiornis nicht gewesen sein. Es ist fraglich, ob diese ungewöhnliche Merkmalskombination eine weitere Evolution in Richtung Vögel überhaupt gefördert haben kann.
Handelt es sich vielleicht um eine Rückbildung, wie das auch bei einigen kreidezeitlichen Formen vermutet wird? Das aber wäre insofern evolutionstheoretisch problematisch, als Anchiornis auf ca. 160 Millionen Jahre und damit ca. 10 Millionen Jahre älter als Archaeopteryx datiert wird. Denn dann kämen ausgerechnet Formen mit rückgebildeten Federn in der stratigraphischen Abfolge unter den eindeutig befiederten Formen als erste, was eine evolutionäre Abfolge auf den Kopf stellen würde. Darauf deuten möglicherweise auch phylogenetische Analysen hin, wie im Folgenden dargestellt wird.
Vorläufer oder Rückbildung? Longrich et al. (2012) diskutieren eine zweite Schwierigkeit: Die phylogenetischen Positionen von Archaeopteryx und Anchiornis sind unsicher. Nach gegenwärtig gut begründeter, wenn auch unsicherer Deutung sind beide Gattungen am nächsten mit den Deinonychosauriden verwandt und befinden sich daher nicht in der Linie, die zu den Vögeln führt (vgl. Xu et al. 2011 und Fällt eine Ikone vom Sockel?, https://www.genesisnet.info/index.php?News=174). Sollte diese Position zutreffen, wären die langen, asymmetrischen Schwungfedern bei Archaeopteryx einerseits und bei modernen Vögeln andererseits konvergent entstanden (also zweimal unabhängig). Oder die kurzen symmetrischen Schwungfedern von Anchiornis sind abgeleitet (d. h. somit rückgebildet), mithin keine Vorläufer-Ausprägung. Es könnte aber auch das mehrlagige Feder-Arrangement für den Archaeopteryx-Deinonychosaurier-Zweig abgeleitet und nicht primitiv sein. Zudem hat der Dromaeosauride Microraptor gui lange, asymmetrische Handschwingen wie Archaeopteryx und wie die heute lebenden Neornithes (Deckfedern sind bei Microraptor gui unbekannt). Microraptor könnte die fortschrittliche Federmorphologie unabhängig erworben haben oder die Situation bei Anchiornis sich erneut als Rückbildung herausstellen (Longrich et al. 2012, 2264). Auf der Basis des gegenwärtigen Kenntnisstandes kann unter diesen Alternativen nicht die zutreffende bestimmt werden. Sie sind aber allesamt evolutionstheoretisch nicht unproblematisch.
Alternative? Aus den genannten Gründen bietet sich ein alternativer Ansatz an: Die Vielfalt von Formen hängt mit einer hohen Diversität von Lebensräumen zusammen, ohne eine phylogenetische Bedeutung zu haben. Die Lebensräume sind uns teilweise unbekannt, was das Verständnis der teilweise widersprüchlich erscheinenden Merkmalskonstellationen (s. o.) der jurassischen und unterkretazischen Formen aus dem mutmaßlichen „Dinosaurier-Vogel-Übergangsfeld“ erschwert. Die Formenvielfalt erfordert jetzt bereits ein erhebliches Ausmaß an Konvergenzen und/oder Reversionen (mit Parallelentwicklungen als „Homoplasien“ begrifflich zusammengefasst). So müssen die Gattungen mit befiederten Vorder- und Hinterbeinen (Anchiornis, der Dromaeosauride Microraptor und die zu den Avialae gerechnete Gattung Pedopenna) verschiedenen Linien zugeordnet werden. Evolutionäre Mechanismen, die dieses gehäufte Auftreten von Homoplasien durch konkrete Szenarien plausibel machen könnten, sind bislang nicht beschrieben (vgl. Braun 2012; Junker 2003).
Es gibt Prüfmöglichkeiten für evolutionäre und ökologische Deutungen: Führen weitere Funde zu noch komplexeren Verwandtschaftsbeziehungen und einer weiteren Vernetzung von Merkmalsübereinstimmungen oder kristallisieren sich zunehmend stammesgeschichtliche Linien heraus? Und, sollte ersteres der Fall sein: Können evolutionäre Mechanismen nachgewiesen werden, die zu widersprüchlichen Merkmalsverteilungen führen?
Literatur
Alonso DP, Milner AC, Ketcham RA, Cookson JM & Rowe TM (2004) The avian nature of the brain and inner ear of Archaeopteryx. Nature 430, 666-669.
Braun HB (2012) Warten auf einen neuen Einstein. Stud. Int. J. 19, 12-19.
Carney RM, Vinther J, Shawkey MD, D’Alba L & Ackermann J (2012) New evidence on the colour and nature of the isolated Archaeopteryx feather. Nature Comm. 3, Art.-No. 637, doi:10.1038/ncomms1642
Hu D, Hou L, Zhang L & Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 460-463.
Junker R (2003) Baum, Baukasten, Netzwerk. Ist die evolutionäre Systematik zirkelschlüssig? Stud. Int. J. 10, 3-11.
Longrich NR, Vinther J, Meng Q, Li Q & Russell AP (2012) Primitive Wing Feather Arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Curr. Biol. 22, 2262-2267.
Nudds RL & Dyke GJ (2010a) Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability. Science 328, 887-889.
Nudds RL & Dyke GJ (2010b) Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability”. Science 330, 320d.
Paul GS (2010) Comment on “Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability”. Science 330, 320b.
Stettenheim PR (2000) The integumentary morphology of modern birds; Am. Zool. 40, 461-477.
Xu X, You H, Du K & Han F (2011) An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 475, 465-470.
Autor dieser News: Reinhard Junker
© 2013, http://www.genesisnet.info/schoepfung_evolution/n193.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
12.12.11 Verzwickte Biogeographie des Hoatzins
Neue Fossilfunde der ungewöhnlichen Hoatzins (Schopfhühner) werfen Fragen nach ihrer Entstehung auf. Bisher hatte man angenommen, dass sie erst nach der Trennung von Afrika und Südamerika entstanden sind, doch Funde aus Namibia lassen nur zwei Schlussfolgerungen zu: Entweder die Hoatzins entstanden schon viel früher, sind dann aber über den größten Teil ihrer Geschichte fossil nicht belegt, oder sie haben den Atlantik auf Treibholz überquert, was aber fast schon einem Wunder gleich käme. Insgesamt passt die (paläo)geographische Verteilung der Hoatzins kaum zu Evolutionsszenarien.
Eine ungewöhnliche Vogelart in den tropischen Regenwäldern des nördlichen Südamerika ist der Hoatzin, auch Schopfhuhn genannt. Auffällig sind der kleine Kopf und eine mehrere Zentimeter lange Haube, dazu sind Hals und Schwanz sehr lang. Hoatzins erreichen eine Länge von bis zu 70 cm und werden bis zu knapp 1 kg schwer. Bei der Gefiederfärbung dominieren beige und braune Töne. Wegen ihrer schwachen Flugmuskulatur sind Hoatzins schlechte Flieger, aber auch die Beine werden kaum zur Fortbewegung genutzt, obwohl sie kräftig sind. Stattdessen kriechen sie und schieben sich durch das Geäst, eine für Vögel sehr eigenartige Fortbewegungsweise. Als weitere ungewöhnliche Merkmale besitzen Jungvögel Krallen an den Flügeln, die ihnen erlauben, auf allen Vieren im Geäst herumzuklettern, außerdem ein unter Vögeln einmaliges Verdauungssystem, das an Wiederkäuer erinnert. Die Verdauung der pflanzlichen Nahrung (hauptsächlich Blätter) findet im muskulösen Kropf und in der unteren Speiseröhre statt, nicht im Magen. Entsprechend ist der Kropf fünfzigmal so groß wie der Magen und macht 13 % des Gesamtgewichts des Vogels aus (Niethammer 1993). In diesen Merkmalen unterscheidet sich der Hoatzin von allen anderen Vögeln. Die schwach ausgeprägte Flugfähigkeit ist kein Ausdruck von Primitivität, sondern hängt mit dieser Ernährungsweise zusammen, da sie eine deutliche Verkleinerung des Brustbeins und damit der Flugmuskulatur erfordert. Auch die Krallen sind eher sekundäre Spezialisierungen als evolutionäre Überbleibsel (vgl. Niethammer 1993, 77).
Die Verwandtschaft des Hoatzins ist ungeklärt, auch unter Einbeziehung molekularer Merkmale (Hackett et al. 2008, Mayr et al. 2011). Daher wird er meist in eine eigene Ordnung (Opisthocomiformes) gestellt. Diverse vermutete Zugehörigkeiten (z. B. zu den Turakos, den Kuckucksvögeln, den Tauben oder den Hühnervögeln) wurden durch genetische Untersuchungen widerlegt (Hackett et al. 2008). Die evolutionäre Geschichte der Hoatzins ist unbekannt (Mayr et al. 2011). Bis vor kurzen war nicht nur das Verbreitungsgebiet der heute lebenden Opisthocomiformes auf Südamerika beschränkt, sondern auch das der fossilen Formen. Doch nun beschreiben Mayr et al. (2011) zwei neue Funde fossiler Opisthocomiformes aus Brasilien und – überraschenderweise – aus Namibia. Jeweils ein Oberarmknochen, Schulterblatt und Rabenschnabelbein aus dem Oligo-Miozän (22-24 Millionen Isotopenjahre) Brasiliens sind dem Schultergürtel des heutigen Hoatzin sehr ähnlich. Die Forscher schließen daraus, dass die spezialisierte Ernährung der Hoatzins bei dieser fossilen Form bereits verwirklicht war. Besonders bemerkenswert ist aber der Fund der Gattung Namibiavis aus dem Miozän Namibias (auf etwa 17 Millionen Isotopenjahre datiert). Fossile Knochen (drei Rabenschnabelbeine, sechs Oberarmknochen und ein Unterschenkelknochen) dieser Gattung waren schon seit einigen Jahren bekannt und bislang zu einer ausgestorbenen Familie der Kranichvögel gestellt worden. Eine genauere Untersuchung zeigte, dass diese Klassifikation fehlerhaft war, denn die Fossilien weisen charakteristische Knochenmerkmale von Hoatzins auf. Nun vermuten die Forscher, dass die Hoatzins nicht in Südamerika, sondern in Afrika entstanden sind.
Die geographische Verteilung wirft allerdings Fragen auf: Wie gelangten die Hoatzins bzw. nah verwandte Vögel über den Atlantik von einem Kontinent zum anderen? Afrika und Südamerika waren nach geologischen Befunden bereits vor 100 Millionen Isotopenjahren zur Kreidezeit weit voneinander getrennt. Dass diese Vogelgruppe damals schon existierte, wird im evolutionstheoretischen Rahmen nicht angenommen. Die Bewältigung einer so großen Flugdistanz ist für die kaum flugfähigen Vögel nicht möglich, auch wenn man annehmen würde, dass die miozänen Formen etwas besser fliegen konnten. Aufgrund des fossilen Materials schließen Mayr et al. (2011) jedoch aus, dass sie gute Flieger waren. Die Forscher nehmen daher an, dass sie den weiten Weg auf Treibholz-Inseln schafften; es handle sich um das erste Beispiel eines transatlantischen Raftings bei Vögeln. Doch diese Lösung des biogeographischen Problems ist nur aufgrund evolutionstheoretischer Voraussetzungen gefordert und kaum glaubwürdig. Die sich von Blättern ernährenden Vögel müssten einen ordentlichen Nahrungsvorrat auf ihrer Reise gehabt haben. Der Hinweis von Mayr et al., dass Rafting über eine so große Strecke schon länger auch bei Säugetieren und Reptilien angenommen werde, macht diese Annahme nicht glaubwürdiger, da die Begründung dieselbe ist: durch Ozeane getrennte geographische Verbreitung, nicht aber Kenntnisse darüber, dass und wie dies tatsächlich möglich ist. Heads (2009, 108) hält eine solche Überquerung für Primaten, bei denen ozeanweite Ausbreitung ebenfalls diskutiert wird, für ausgeschlossen.
Alternativ könnte man der Spur nachgehen, dass nicht fossil belegte Hoatzins schon viel früher als fossil belegt in geologisch nicht überlieferten Lebensräumen existierten, und zwar bereits vor der Kreidezeit, als die Kontinente noch nicht getrennt waren. Heads (2009) hält aufgrund der biogeographischen Verteilung die Annahme für geboten, dass ein Vorläufer der Primaten bereits im Jura vor 185 Millionen Isotopenjahren gelebt haben könnte. Das würde allerdings bedeuten, dass bei mehreren Formengruppen während der meisten Zeit ihrer Existenz fossile Belege fehlen. Ob solche Szenarien im Rahmen großer Zeiträume glaubhaft sind, kann man bezweifeln.
Literatur
Hackett SJ, Kimball RT, Reddy S, Bowie RCK, Braun EL, Braun MJ, Chojnowski JL, Cox WA, Han K-L, Harshman J, Huddleston CJ, Marks BD, Miglia KJ, Moore WS, Sheldon FH, Steadman DW, Witt CC & Yuri T (2008) A phylogenomic study of birds reveals their evolutionary history. Science 320, 1763-1767.
Heads M (2009) Evolution and biogeography of primates: a new model based on molecular phylogenetics, vicariance and plate tectonics. Zoologica Scripta 39, 107-127; doi: 10.1111/j.1463-6409.2009.00411.x
Mayr G, Alvarenga H & Mourer-Chauviré C (2011) Out of Africa: Fossils shed light on the origin of the hoatzin, an iconic Neotropic bird. Naturwissenschaften 98, 961-966.
Niethammer G (1993) Unterordnung Hoatzins. In: Grzimek B u. a. (Hg) Grzimeks Tierleben. Vögel 2. München, S. 76-78.
Autor dieser News: Reinhard Junker
© 2011, http://www.genesisnet.info/schoepfung_evolution/n179.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
04.08.11 Fällt eine Ikone vom Sockel?
Ein neuer Fund könnte den berühmten Urvogel Archaeopteryx degradieren
Ein neuer Fossilfund eines befiederten, etwa huhngroßen Dinosauriers aus der Gruppe der Theropoden zeigt deutliche Ähnlichkeiten mit dem berühmten „Urvogel“ Archaeopteryx. Eine Merkmalsanalyse unter Einbeziehung dieser neuen Gattung führt zum Ergebnis, dass Archaeopteryx in die Gruppe der Deinonychosauria gestellt werden muss, die evolutionstheoretisch nicht als Vorläufergruppe der Vögel betrachtet werden kann. Damit würde Archaeopteryx seinen Status als „Urvogel“ verlieren. Dennoch sind die Konsequenzen für evolutionstheoretische Interpretationen eher gering. Die neue Gattung Xiaotingia vergrößert die Vielfalt früher befiederter Dinosaurier. Aufgrund des häufigen Auftretens von Mosaikformen mit Merkmalswidersprüchen und damit einhergehenden verbreiteten Konvergenzen* ergeben sich Probleme für das Verständnis evolutionärer Abläufe, andererseits ist diese Situation aus der Sicht des Schöpfungsparadigmas leichter verstehbar.
*Konvergenzen (= Entstehung ähnlicher Strukturen von unähnlichen Vorstadien)
Ausgerechnet zum 150. Geburtstag seiner Erstbeschreibung gibt es schlechte Nachrichten für den berühmten Urvogel Archaeopteryx: Er soll gar kein Vogel gewesen sein. Das gilt jedenfalls, wenn man den Untersuchungen an einem neuen Fund eines hühnergroßen, befiederten Dinosauriers – Xiaotingia zenghi – und den daran anknüpfenden Deutungen folgt, die eine chinesische Forschergruppe veröffentlicht hat (Xu et al. 2011; Bild unter http://scienceblogs.com/pharyngula/upload/2011/07/xiaotingia.php). Xiaotingia gehört zu den Theropoden, das sind zweibeinige, meist relativ kleine Raubdinosaurier, die evolutionstheoretisch in die nähere Verwandtschaft zu den Vögeln gestellt werden. Die stratigraphische und damit zeitliche Einordnung ist unsicher, da das Fossil von einem Fossilienhändler erworben wurde und daher der genaue Fundort unbekannt ist. Nach Auffassung von Xu et al. (2011) stammt der Fund aus der Tiaojishan-Formation des Oberjuras in der chinesischen Provinz Liaoning, diese Formation wird auf etwa 155 Millionen Jahre datiert, womit Xiaotingia geringfügig älter ist als Archaeopteryx.
„An icon knocked from ist perch“ – Eine Ikone wurde von ihrem Ast gestoßen – mit dieser reißerischen Zeile überschreibt denn auch der Paläontologie Lawrence M. Witmer seinen Kommentar zur Originalarbeit von Xu et al. in Nature. Mit der „Ikone“ ist Archaeopteryx gemeint, von dem seit 1861 mindestens zehn Exemplare in den Plattenkalken des fränkischen Altmühltals gefunden wurden. In der Tat ist Archaeopteryx das Sinnbild für eine evolutionäre Übergangsform. Seinen Merkmalsmix aus reptiltypischen und vogeltypischen Merkmalen muss jeder Gymnasiast lernen, der das Fach Biologie bis zum Abitur belegt.
Warum soll Archaeopteryx „kein Vogel mehr“ sein?1 Wie aber kommt es, dass Archaeopteryx seinen prominenten Platz als Urvogel verliert, ohne dass es eine einzige neue Erkenntnis über ihn gibt? Die Antwort liegt im Procedere der Cladistik, einem systematischen Verfahren, das auf der Basis von Merkmalsvergleichen die Organismen in ein Verzweigungsschema (ein sog. Cladogramm) bringt. Dieses stellt in evolutionstheoretischer Interpretation ein hypothetisches Abstammungsschema (ein Phylogramm) dar. (Für die nachfolgenden Ausführungen sei ein Blick auf das Cladogramm unter http://scienceblogs.com/pharyngula/upload/2011/07/bird_phylo.php sehr empfohlen!)
Bislang war Archaeopteryx an der Basis der Avialae (Vögel und verwandte Formen) platziert und damit evolutionstheoretisch als Urvogel interpretierbar. Durch den neuen Fund Xiaotingia verschieben sich jedoch die cladistischen Verwandtschaftsbeziehungen. Xiaotingia weist deutliche Ähnlichkeiten mit Anchiornis auf, der zu den Deinonychosauria gestellt wird, einer Gruppe der theropoden Dinosaurier, die die nicht zu den Vögeln gehört und auch nicht zu den Vogelvorläufern gestellt wird. Aber auch mit Archaeopteryx teilt Xiaotingia so viele gemeinsame Merkmale, dass nun nach den Regeln der Cladistik auch der Urvogel – anders als bisher – ebenfalls zu den Deinonychosauria gestellt werden muss. Die drei Gattungen Anchiornis, Xiaotingia und Archaeopteryx werden innerhalb der Gruppe der Deinonychosauria als Archaeopterygidae zusammengefasst. Die Archaeopterygidae und damit auch Archaeopteryx repräsentieren somit einen Zweig des Stammbaums, der nicht zu den Vögeln führt (vgl. Abb. http://scienceblogs.com/pharyngula/upload/2011/07/bird_phylo.php).
Für die nahe Verwandtschaft von Xiaotingia mit Archaeopteryx werden u. a. die flache Schnauze, eine ausgedehnte Region hinter der Augenhöhle, die Form des Schlüsselbeins, extrem lange mittlere und letzte Fingerknochen, die spezialisierte zweite Zehe und Merkmale des Beckens genannt. (Details bei Xu et al. 2011, 467ff.) Frühe Vögel der Avialae haben deutlich andere Schädel.2 Außerdem zeigt sich, dass viele Merkmale, die bislang als vogeltypisch galten, darunter die langen und robusten Vordergliedmaßen und der Besitz von Federn, weiter verbreitet waren und für die größere Gruppe der Paraves charakteristisch sind (Xu et al. 2011, 465). (Die Paraves umfassen die Avialae und die Deinonychosauria.)
Von Xiaotingia ist nur ein Exemplar bekannt, das insgesamt relativ gut erhalten ist. Xiaotingia hatte scharfe Zähne und Krallen am Ende der Vordergliedmaßen. Um das ganze Skelett herum sind schwache Federabdrücke erhalten, auch an den hinteren Extremitäten und in der Nähe von dessen Zehen, was auch von Anchiornis bekannt ist. Xiaotingia war also anders als Archaeopteryx vierflügelig. Vierflügelige Formen sind schon seit einigen Jahren auch aus der Gattung Microraptor bekannt (Xu et al. 2003). Leider sind die Federn zu schlecht erhalten, um irgendwelche Details an den Abdrücken erkennen zu können. Die Federn in der Nähe des Femur (Schienbein) sind mit über 5 cm Länge sehr lang.
Konsequenzen. Aus wissenschaftlicher Sicht sind die Konsequenzen aus dem neuen Fund eher gering. Verschiebungen in den Dendrogrammen aufgrund neuer Funde oder nochmaligen genaueren Untersuchungen gab es immer wieder und schon manche Arten wurden hin und her geschoben (Witmer 2011). Wenn Archaeopteryx nicht mehr an der Basis der Avialae steht, vergrößert sich die Lücke im Bereich der frühen Vögel, bemerkt P. Z. Myers auf seinem Blog (http://scienceblogs.com/pharyngula/2011/07/xiaotingia_zhengi.php). Andererseits löst sich ein Merkmalswiderspruch auf. Archaeopteryx hatte das Gebiss eines Räubers, während andere frühe Vogelfossilien Pflanzenfresser waren und in dieser Hinsicht besser mit mutmaßlichen Theropoden-Vorfahren verknüpft werden können (Witmer 2011, 459).
Die Bedeutung der Neupositionierung liegt jedoch eher darin, dass mit Archaeopteryx eine der bekanntesten fossilen Gattungen von einer solchen Umgruppierung betroffen ist und dies den Status als „Urvogel“ tangiert. Bei oberflächlicher Betrachtung könnte man meinen, dass ein evolutionäres Bindeglied gefallen ist, doch die Situation ist komplizierter. Denn Archaeopteryx stand schon lange nicht mehr alleine im Übergangsbereich von Reptilien und Vögeln. In den letzten etwa 30 Jahren wurden vor allem in China zahlreiche neue Fossilien von Vögeln und vogelähnlichen Dinosauriern entdeckt, so dass die Vielfalt der fossilen Formen an der Basis des evolutionären Stammbaums sich mittlerweile als schwer zu entwirrendes Gestrüpp darstellt. Modelle für evolutionäre Übergangsformen sind also vorhanden. Die Indizien- und Deutungslage bezüglich der Frage nach Übergangsformen hat sich seit einiger Zeit bereits deutlich verschoben. Wo die evolutionstheoretischen Probleme heute liegen, sei nachfolgend erörtert.
Witmer (2011) schreibt in seinem Kommentar, die Grenzen zwischen den verschiedenen Gruppen, die an die Basis der Vogelevolution gestellt werden, seien zunehmend verwischt worden. Auf seinem Blog (http://witmerlab.wordpress.com) behauptet er darüber hinaus: „Eine Vorhersage von Evolution ist, dass die Arten sich zunehmend ähnlich und die Unterschiede unbedeutender werden, wenn wir in der Zeit zurück gehen und uns einem gemeinsamen Vorfahren annähern.“ Doch das stimmt so nicht mit der fossilen Befundsituation überein. Witmer selber setzt fort: „Es gab in einem massiven Umfang unabhängige Evolution (Homoplasie) verschiedener ausgetüftelter Eigenschaften, was es schwierig macht, den Knoten dieses evolutionären Beginns aufzutrennen …“ Unter Homoplasien werden Konvergenzen*, Parallelentwicklungen und Rückentwicklungen zusammengefasst, also solche Ähnlichkeiten, die nicht auf gemeinsame Vorfahren zurückgeführt werden.
*Konvergenzen (= Entstehung ähnlicher Strukturen von unähnlichen Vorstadien)
Der fossile Befund zeigt also nicht zunehmende Ähnlichkeit an der Basis des mutmaßlichen evolutionären Stammbaums, es gibt vielmehr verschiedene Mosaikformen und das erzwingt die Annahme zahlreicher Konvergenzen – diese machen ihrerseits die Taxonomie und die Zuordnung zu einzelnen Gruppen schwierig. Xu et al. (2011, 465) schreiben, dass viele basale Avialae (Gruppe, die zu den häutigen Vögeln führt) sich „beträchtlich“ von Archaeopteryx unterscheiden. Die Forscher diskutieren eine Reihe von Merkmalen, die die Archaeopterygidae (Archaeopteryx, Anchiornis und Xiaotingia) einerseits mit anderen Deinonychosauria teilen, die sie andererseits von anderen basalen Avialae-Gruppen unterscheiden (Xu et al. 2011, 468f.). Xu et al. stellen aber auch wie Witmer fest, dass es viele funktional signifikante Homoplasien gebe und dass Homoplasien weit verbreitet seien. Viele auffällige anatomische Merkmale von Xiaotingia gebe es in verschiedenen Gruppen der Paraves. Und dieses Phänomen sei auch bei anderen größeren Übergängen bekannt.3 Vor einem Jahr berichteten Choiniere et al. (2010) über einen befiederten Alvarezsauriden (Haplocheirus sollers), der trotz der Befiederung weit von den mutmaßlichen Vogelvorläufern entfernt ist; auch diese Forscher konstatieren „extreme morphologische Konvergenzen“ (vgl. Junker 2010). Es muss immer wieder darauf hingewiesen werden, dass diese Situation evolutionstheoretisch unerwartet ist und Fragen nach den Mechanismen aufwirft, die häufig zu gleichen Merkmalen hätten führen können. Xu et al. (2011, 469) weisen darauf hin, dass wegen der zahlreichen Homoplasien der neue Stammbaum nur schwach gestützt ist. Witmer (2011, 459) bemerkt, dass der nächste Fund das Bild wieder umdrehen könnte.
All dies zeigt, dass Merkmale an sich keine verlässlichen Verwandtschaftsanzeiger sein können. Je nach Gesamtbefund werden bestimmte Ähnlichkeiten als Homologien (d. h. als Belege für gemeinsame Abstammung) oder als Homoplasien gewertet; das Pendel kann hin und her schlagen. Witmer schließt seinen Kommentar damit, dass evolutionäre Ursprünge nun mal chaotische Angelegenheiten („messy affairs“) seien.
Ob nun Archaeopteryx ein Vogel war oder nicht, ist Definitionssache und auch eine Frage der Methode, wie die Merkmalsspektren der Arten dargestellt werden. Nach der cladistischen Analyse von Xu et al. befindet sich Archaeopteryx auf einem evolutionären Seitenast, der zu einer Gruppe (Deinonychosauria) gehört, die nicht zu den heutigen Vögeln leitet. Wie erwähnt, kann der nächste Fund dies schon wieder ändern. Das Merkmalsspektrum von Archaeopteryx ändert sich freilich nicht, solange keine neuen Funde von ihm gemacht werden. Und daher bleibt es dabei, dass Archaeopteryx flugtaugliche Federn, den Federn heutiger Vögel vergleichbar, und manche anderen vogeltypischen Merkmale hatte. Bevor weitreichende Schlüsse gezogen werden, sollte die monografische Bearbeitung abgewartet werden. Unklar ist auch, wie die paläobiogeographische Verteilung der Archaeopterygidae erklärt werden kann. Archaeopteryx sollte also nicht vorschnell als die „Urfeder“ abgeschrieben werden.
Und aus der Sicht der Schöpfungslehre? Zunächst muss auf eine eigentlich banale Selbstverständlichkeit hingewiesen werden: Ein Cladogramm ist nicht notwendigerweise ein Phylogramm. Man benötigt weder die Voraussetzung von Evolution, um ein Cladogramm aufzustellen, noch besteht die Notwendigkeit, ein Cladogramm als Phylogramm zu interpretieren. Cladogramme sind Ordnungsschemata, die evolutionstheoretisch interpretiert werden können. Und weiter ist das cladistische Verfahren zwar Standard, aber nicht konkurrenzlos. Wenn Homoplasien überhand nehmen, werden cladistische Ergebnisse unsicher (vgl. Anm. 3 und die von Xu et al. konstatierte schwache Stützung des neuen Cladogramms, s. o.). Cladogramme werden so konstruiert, dass möglichst wenige Homoplasien auftreten, da Homoplasien als relativ unwahrscheinlich gelten. Dies hängt mit den zugrundegelegten Vorstellungen über Evolutionsmechanismen zusammen. Könnte man Evolutionsmechanismen plausibel benennen, die Homoplasien wahrscheinlich machen, wäre dagegen dem Cladismus der Boden entzogen. In einer schöpfungsorientierten Perspektive kann man den Befund der verbreiteten Mosaikformen mit anderen Augen sehen und als Ausdruck eines Baukastensystems werten. Merkmale sind dann beliebig kombinierbar, sofern der Gesamtkomplex eine lebensfähige Art ergibt. Das Nebeneinander verschiedener Mosaikformen ist hier kein grundsätzliches Problem.
Es bleibt für die Schöpfungslehre aber – wie schon zuvor – die Frage nach den stratigraphischen Abfolgen: Warum ist die Vogelwelt im Jura und in der Kreide eine ganz andere als die im Tertiär?
Literatur
Choiniere JN, Xu X, Clark JM, Forster CA, Guo Y & Han F (2010) A Basal Alvarezsauroid Theropod from the Early Late Jurassic of Xinjiang, China. Science 327, 571-574.
Junker R (2010) Löst Haplocheirus das Zeit-Paradox der Vogelevolution? Stud. Int. J. 17, 36-38.
Witmer LM (2011) An icon knocked from its perch. Nature 475, 458-459.
Xu ZZ, Wang X, Kuang X, Zhang F & Du X (2003) Fourwinged dinosaurs from China. Nature 421, 335-340.
Xu X, You H, Du K & Han F (2011) An Archaeopteryx--like theropod from China and the origin of Avialae. Nature 475, 465-470.
Anmerkungen
1 „In other words, Archaeopteryx was no longer a bird“ (Witmer 2011, 458).
2 Vgl. auch http://www.nature.com/news/2011/110727/full/news.2011.443.html.
3 „This low support is partly caused by various homoplasies, many of which are functionally significant, that are widely distributed across coelurosaurian phylogeny. Xiaotingia possesses salient anatomical features also seen in different paravian taxa, further highlighting the phenomenon of widespread homoplasy. This phenomenon is also seen in some other major transitions, including the origins of major mammalian groups, and creates difficulties in recovering robust phylogenies“ (Xu et al. 2011, 467).
Autor dieser News: Reinhard Junker
© 2011, http://www.genesisnet.info/schoepfung_evolution/n174.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
09.02.10 Löst Haplocheirus das Zeit-Paradox der Vogelevolution?
Der Fund eines Alvarezsauriden der Gattung Haplocheirus scheint ein Paradox der Vogelevolution zu lösen, das darin besteht, dass die am ehesten als Vogelvorfahren geeigneten Formen später in der Fossilabfolge auftauchen als die ältesten Vögel. Haplocheirus könnte einfache Federn gehabt haben und wird etwa 15 Millionen Jahre älter datiert als der „Urvogel“ Archaeopteryx. Haplocheirus kann jedoch aufgrund spezialisierter Merkmale nicht zu den Vogelvorfahren gestellt werden. Für die Frage nach der evolutiven Entstehung der Vögel ist mit dem neuen Fund außer einer gewissen Entschärfung des Zeit-Paradoxons nichts gewonnen. Vielmehr trägt Haplocheirus weiter dazu bei, dass „extreme morphologische Konvergenzen“ (Choiniere et al. 2010, 571), also die unabhängige Entstehung gemeinsamer Merkmale, angenommen werden müssen.
Kürzlich berichteten Wissenschaftler um Xing Xu vom Institute of Vertebrate Paleontology Beijing von einem in mehrfacher Hinsicht bemerkenswerten Fund eines Alvarezsauriers (Choiniere et al. 2010). Diese Gruppe wird zur Theropoden-Dinosaurier-Gruppe der Maniraptora gestellt, aus denen sich nach verbreiteter Auffassung die Vögel entwickelt haben sollen. Bislang wurden die schlanken und langbeinigen, wegen ihrer kurzen, spezialisierten Vorderextremitäten rätselhaften Alvarezsauriden allerdings als flugunfähig gewordene Nachkommen von Vögeln angesehen, denn sie weisen im Skelettbau eine Reihe von Gemeinsamkeiten mit den Vögeln auf: ein kleines Brustbein mit einer Brustbeinleiste, die Verschmelzung von Handwurzel- und Mittelhandknochen und federartige Strukturen bei der Gattung Shuvuuia. Die Vorderextremitäten sind aber für einen Flug viel zu kurz. Die Alvarezsauriden kannte man bislang erst aus Schichtfolgen, die sehr viel jünger sind als die ältesten Vögel. Die Vorderbeine waren, unterstützt von starken Brust- und Armmuskeln, sehr wahrscheinlich zum Graben nach staatenbildenden Insekten geeignet. Der stark vergrößerte Daumen besaß eine Klaue, während die beiden anderen Finger stark reduziert waren.
Eine zum Graben spezialisierte Hand besaß auch der neue Fund aus dieser Gruppe, Haplocheirus sollers; der Name bedeutet etwa „einfache geschickte Hand“. Dieser Fund ist deshalb besonders bemerkenswert, weil er in Schichtfolgen des unteren Oberjura der Wüste Gobi (China) gefunden wurde, die auf etwa 160 Millionen Jahre datiert werden (Choiniere et al. 2010). Diese Schichten sind damit ca. 15 Millionen Jahre älter als der berühmte Urvogel Archaeopteryx aus dem obersten Oberjura (Tithonium-Stufe) Frankens. Bislang waren gut erhaltene Fossilien, die am ehesten als Vorfahrenkandidaten der Vögel interpretiert werden konnten, in Schichtfolgen gefunden worden, die jünger (oft deutlich jünger) als der Oberjura sind. Das betraf auch Fossilien, die Federn oder federartige Strukturen besaßen (deren Interpretation teilweise umstritten ist). Dieses „Zeit-Paradox“ – vereinfacht: Die Vorfahren der Vögel erscheinen fossil nach den Vögeln – ist viel diskutiert worden. Solche unpassenden Abfolgen können natürlich immer auf Lücken in der Fossilüberlieferung zurückgeführt werden. Für manche Kritiker ist dieses Zeit-Paradox aber auch einer der Gründe, die evolutive Abfolge Dinosaurier -> Vögel anzuzweifeln; die Gemeinsamkeiten vieler Theropoden mit den Vögeln werden von ihnen als Konvergenzen angesehen (also als unabhängig entstandene Gemeinsamkeiten, die keine Hinweise auf gemeinsame Vorfahren sind). (Dass diese Gemeinsamkeiten auch abgesehen von den stratigraphischen Fundhorizonten genausogut Konvergenzen sein könnten, wird auch durch eine neue cladistische Studie gezeigt [James & Pourtless 2009].)
Nun scheint dieses Zeit-Paradox mit dem Fund von Haplocheirus aufgelöst zu sein, und zwar „ein für alle Mal“, wird der Paläontologe Hans-Dieter Suess von Stone (2010) zitiert. Auch die Deutung, es handle sich um flugunfähig gewordene Vögel sei damit definitiv erledigt, zumal Haplocheirus einige vogelähnliche Merkmale fehlen, die die viel jüngeren Funde zeigen (Choiniere 2010; Stone 2010). Haplocheirus besitzt möglicherweise federartige Strukturen an den Armen (so in einer Rekonstruktion bei Stone 2010). Die zahlreichen Gemeinsamkeiten zwischen den jüngeren Alvarezsauriden und Vögeln müssen daher konvergent entstanden sein. Einmal mehr wird deutlich, wie unsicher die Schlüsse sind, die aus Merkmalsüberseinstimmungen gezogen werden.
Doch ist das Zeit-Paradox damit wirklich gelöst? Ja – insofern, als ein Vertreter aus der Gruppe der Maniraptora in älteren Schichtfolgen gefunden wurde. In dieser Gruppe werden am ehesten die Vogelvorfahren gesucht. Die Beschreiber des Fundes, Choiniere et al. (2010), stellen Haplocheirus jedoch nicht in die verwandtschaftliche Nähe von Archaeopteryx und der Linie, die zu den Vögeln führen soll. Die zum Graben spezialisierten Vorderbeine, die auch Haplocheirus besaß, sind zudem nicht gerade eine passende Vorläuferstruktur für Flügel. Als Form, die zu den Vögeln überleitet, kann Haplocheirus also nicht interpretiert werden. Ungewöhnlich ist auch die Größe. Der Schwanz ist zwar nicht erhalten, aber aus dem Kopf und Rumpf kann eine Körpergröße von 190-230 cm geschlossen werden, womit diese Gattung die größte unter den Alvarezsauriden ist. Daraus ergibt sich in dieser Gruppe ein phylogenetischer Trend hin zu kleineren Formen, was unter Dinosauriern ungewöhnlich ist.
Bemerkenswert bei diesem Fund ist auch die große zeitliche Lücke zu den kreidezeitlichen Formen dieser Familie; sie beträgt 63 Millionen Jahre nach radiometrischen Datierungen, womit eine lange sogenannte „Geisterlinie“ („ghost range“) angenommen werden muss.
Für die Frage nach der evolutiven Entstehung der Vögel ist mit dem neuen Fund außer einer gewissen Entschärfung des Zeit-Paradoxons nichts gewonnen. Es ist nun klar, dass die Alvarezsauriden als eine zur Vogellinie parallele Linie angesehen werden müssen, deren Merkmalsübereinstimmungen nicht auf Abstammung von den Vögeln zurückgeführt werden können. Vielmehr müssen sie als „extreme morphologische Konvergenzen“ (Choiniere et al. 2010, 571) interpretiert werden – ein Umstand, der evolutionäre Deutungen nicht gerade erleichtert. Der Fund hilft aber zum Verständnis der (Mikro-?)Evolution der Alvarezsauriden, da die Vorderbeine (mit längerem Arm und etwas kürzerer Klaue) und andere Merkmale als Vorfahrenstadium der oberkreidezeitlichen Vertreter dieser Gruppe angesehen werden können.
Literatur
Stone R (2010) Bird-Dinosaurier Link Formed Up, And in Brilliant Technicolor. Science 327, 508.
Choiniere JN, Xu X, Clark JM, Forster CA, Guo Y & Han F (2010) A Basal Alvarezsauroid Theropod from the Early Late Jurassic of Xinjiang, China. Science 327, 571-574.
James FC & Pourtless IV JA (2009) Cladistics and the Origin of Birds: A Review and Two New Analyses. Ornithological Monographs 66, 1-78.
Autor dieser News: Reinhard Junker
© 2010, http://www.genesisnet.info/schoepfung_evolution/n143.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
27.10.09 Vierflügelige Vögel am Anfang?
Der Fund eines vierflügeligen Vogels scheint eine evolutionstheoretische Vorhersage zur Entstehung der Vögel zu bestätigen, aber er stellt auch bisherige Vorstellungen in Frage. Das neu entdeckte Exemplar aus der Gattung Anchiornis besaß gut ausgebildete Federn und ist älter als der bisher älteste unumstrittene Vogel, der „Urvogel“ Archaeopteryx. Ein solcher Fund war erwartet worden. Das „zeitliche Paradox“, dass Formen mit gut ausgebildeten Federn vor solchen mit sogenannten Protofedern fossil auftauchen, löst dieser Fund jedoch nicht. Anchiornis stützt die Hypothese, dass am Anfang der Vogelevolution vierflügelige Formen standen. Die zuletzt favorisierte Hypothese, dass der Vogelflug ausgehend von schnell laufenden zweibeinigen Dinosauriern erworben wurde, wird damit in Frage gestellt.
In den letzten Monaten wurde eine Reihe von Entdeckungen gemacht, die neue Bausteine zur Kenntnis über die ältesten Vögel beisteuerten. Manche entsprechen teilweise evolutionstheoretischen Erwartungen, andere sind diesbezüglich eher überraschend (vgl. „Vögel doch nicht Nachfahren der Dinos?“ http://www.scinexx.de/wissen-aktuell-10026-2009-06-10.html und Wellnhofer 2009).
Als lang gesuchter Mosaikstein in der Evolution der Vögel wird die Gattung Anchiornis gewertet, von der kürzlich ein sehr gut erhaltenes Exemplar beschrieben wurde (Hu et al. 2009). Bisher hatte die unter Evolutionsforschern fast einhellig vertretene Theorie, dass Vögel von zweibeinig sich fortbewegenden Raubdinosauriern (Theropoden) abstammen, mit einem „zeitlichen Paradox“ zu kämpfen. Denn es sind zwar zahlreiche Theropoden bekannt, die Federn oder (z. T. umstrittene) federähnliche Strukturen besaßen, aber sie wurden alle in meist deutlich jüngeren Schichten als der sogenannte „Urvogel“ Archaeopteryx entdeckt. Dessen Federn aber gleichen im Wesentlichen den Federn heutiger Vögel. Man konnte daher erwarten, dass Formen mit Federvorstufen in Schichten zu finden sein sollten, die älter sind als die Oberjuraschichten des Altmühltals, in denen die Archaeopteryx-Fossilien entdeckt wurden.
Der als vogelähnlicher Dinosaurier bezeichnete Anchiornis huxleyi erfüllt nun einerseits diese Erwartung, denn der neue Fund dieser Gattung stammt aus Schichten, die etwas älter als Archaeopteryx sind (Beginn des Oberjura, datiert auf 151-161 Millionen Jahre). Andererseits ist diese Gattung vierflügelig, was nicht ohne Weiteres den Erwartungen für Vogelvorfahren entspricht (s. u.). Das zur Familie der Troodontidae gerechnete Tier hatte gut ausgebildete Federn an Armen und Beinen und damit vier Flügel, ähnlich wie der Dromaeosaurier Microraptor. und die Gattung Pedopenna, die zu den Avialae gerechnet wird. Vorderarm, Hand, Unterschenkel und Fuß hatten jeweils 10-13 lange Schwungfedern. Anders als bei Archaeopteryx und bei Microraptor lag der breitere Teil des Flügels zum Körperzentrum hin. Dennoch halten die Beschreiber Hu et al. (2009) Anchiornis nicht für flugfähig, da die sehr langen Unterschenkel eher auf eine laufende Lebensweise hinweisen würden. Das lange und umfassende Federkleid passe allerdings wiederum dazu nicht.
Federn an den Beinen kommen bei vielen heutigen Vögeln wie auch sehr wahrscheinlich bei Archaeopteryx vor und haben Schutzfunktion und dienen dem Wärmehaushalt. Sie sind dort aber nicht wie bei Anchiornis, Microraptor und Pedopenna in einer zusammenhängenden ebenen Oberfläche angeordnet. Daher ist zu vermuten, dass die Beinfedern dieser fossilen Formen eine andere Funktion hatten, vermuten Hu et al. (2009).
Da die Federn gut ausgebildet sind, bleibt es aber auch mit diesem neuen Fund beim oben erwähnten zeitlichen Paradox, dass Fossilien mit als Protofedern interpretierbaren Strukturen erst deutlich später bekannt sind als solche mit „fertigen“ Federn. (Die Interpretation als „Protofedern“ ist zudem nicht gesichert; vgl. https://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-5.html.)
Die bisher bekannten vierflügeligen Formen unter den vogelähnlichen Dinosauriern sind nicht näher miteinander verwandt (Hu et al. 2009, Witmer 2009). Ihre weite taxonomische Verbreitung spricht evolutionstheoretisch betrachtet dafür, dass die Vorfahren der Vögel vierflügelig waren. Diese Hypothese war mit der 2003 entdeckten Gattung Microraptor diskutabel geworden (damals übrigens eine völlig unerwartete Entdeckung, wie Witmer vermerkt). Solange mit Microraptor nur eine einzige vierflügelige Gattung bekannt war, neigte man dazu, vierflügelige Formen als gescheitertes frühes „Experiment“ der Vogelevolution anzusehen. Mit den weiteren Funden hat sich die Situation nun geändert. Allerdings ist damit die Vorstellung fragwürdig geworden, dass die Vögel von zweibeinig laufenden Theropoden abzuleiten sind. Denn gut ausgebildete Federn an den Beinen passen kaum zu schnellen Läufern. Schneller Lauf gilt aber eine Voraussetzung für die Entstehung des Flugs, ausgehend vom Boden. Damit steigt die Baumtheorie der Flugentstehung wieder im Kurs. Witmer hält die Frage nach der Entstehungsweise des Vogelflugs wieder für offen. Wie Anchiornis geschickt und schnell mit langen Federn laufen konnte (wofür eigentlich die langen Beine sprechen), erfordere „ernsthafte Überlegungen“.
Bemerkenswert ist auch, dass es offenbar mosaikartig verteilte Merkmalskonstellationen bei den frühen vierflügeligen Formen gibt; Witmer spricht von einem „mix-and-match“ und Hu et al. (2009, 462) konstatieren ein komplexes Muster der morphologischen (= die Gestalt betreffend) Evolution und eine schnelle Diversifizierung (= Verschiedenwerden) im Mittel- und frühen Oberjura, die im Zusammenhang mit paläogeographischen (= Geographie in geologischer Vergangenheit) Veränderungen stehen könnte. Die drei vierflügeligen Gattungen sind nach cladistischen Analysen auf verschiedenen evolutiven Ästen angesiedelt; der neue Fund kommt als Vorfahre von Archaeopteryx nicht in Frage (Witmer 2009).
Die rasch auftretende Formenvielfalt früher Vögel und die mosaikartige Merkmalsverteilung könnten als Indizien für ein nicht-evolutionäres, ökologisches Szenario gewertet werden: Das Heraustreten bereits existenter Formen aus geologisch nicht überlieferten Lebensräumen (Stephan 2002) und eine anfangs größere Vielfalt von Formen. Gerade die in den letzten Jahrzehnten explosiv zugenommene Vielfalt fossiler Vögel und vogelähnlicher Dinosaurier zeigt bunte Merkmalsmosaike, die evolutionstheoretisch zahlreiche Konvergenzen und Rückentwicklungen (z. B. auch den mehrfachen Verlust von Flugfähigkeit) erfordern. Außerdem muss eine schnelle Differenzierung angenommen werden (s. o.). Kürzlich wurde auch eine Studie über vogelartige Spuren veröffentlicht, die am ehesten und zwanglos als Spuren von Strandvögeln interpretiert werden können (Genise et al. 2009). Sie werden aber ca. 50 Millionen Jahre älter datiert als Archaeopteryx und der hier vorgestellte Anchiornis – ein Hinweis auf geologisch nicht überlieferte Lebensräume? Ein solches Szenario würde unterstützt, wenn gezeigt werden könnte, dass die betreffenden Fundschichten schnell entstanden sind und das Auftreten der Formenvielfalt in einen kurzen Zeitrahmen gestellt werden kann. Dies kann derzeit aber nur als Projekt für die Schöpfungsforschung formuliert werden.
Literatur
Genise JF, Melchor RN, Archangelsky M, Bala LO, Straneck R & de Valais S (2009) Application of neoichnological studies to behavioural and taphonomic interpretation of fossil bird-like tracks from lacustrine settings: The Late Triassic–Early Jurassic? Santo Domingo Formation, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 272, 143-161.
Hu D, Hou L, Zhang L & Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 460-463.
Stephan M (2002) Der Mensch und die geologische Zeittafel. Holzgerlingen.
Wellnhofer P (2009) Protofedern bei Vogelbeckensauriern? Nat. Rdsch. 62, 311-312.
Witmer LM (2009) Feathered dinosaurs in a tangle. Nature 461, 601-602.
Autor dieser News: Reinhard Junker
© 2009, http://www.genesisnet.info/schoepfung_evolution/n137.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
07.07.09 Proteinfragmente aus Dino-Fossilien klar bestätigt
Ein Team um die Paläontologin Mary Schweitzer hatte 2007 mit einer Veröffentlichung Aufsehen erregt, in der sie Analysen eines fossilen Oberschenkelknochens von Tyrannosaurus rex aus der Kreide so interpretieren, dass in den ca. 68 Millionen Jahre alten Fossilien noch Fragmente (= Bruchstücke) von Proteinen erhalten seien (vgl. Proteine aus einem fossilen Oberschenkelknochen von Tyrannosaurus rex, https://www.genesisnet.info/index.php?News=82). Nachdem bereits zuvor flexibles Gewebe aus dem fossilen Knochen isoliert und beschrieben worden war, hatten die Autoren mit massenspektrometrischen Methoden Peptide (= kurze Kette aus Aminosäuren) von Kollagen (= Strukturprotein des Bindegewebes) identifiziert. Die Arbeit löste intensive und kontroverse Diskussionen aus, da nach bisherigen Erkenntnissen und Erfahrungen die chemisch wenig stabilen Proteine die Prozesse der Fossilisierung und lange Lagerzeit in den Sedimentschichten nicht überdauern.
Nun legen Schweitzer et al. (2009) eine weitere Arbeit nach, in der sie den fossilen Hadrosaurier Brachylophosaurus canadensis (Entenschnabelsaurier) aus der Oberkreide untersuchen. Das Alter geben die Autoren mit ca. 80 Millionen radiometrischen Jahren an. Die Fossilien wurden unter größtmöglichen Vorsichtsmaßnahmen geborgen, im Labor präpariert und untersucht. Auch aus den B. canadensis-Fossilien konnte flexibles Gewebe isoliert werden, das aufgrund seiner mikroskopischen Charakterisierung nicht auf Mikroorganismen (Biofilm) zurückzuführen ist, die das Fossil besiedelt haben.
Immunologische Studien an Extrakten oder an demineralisierten Proben des Fossils ergaben, dass verschiedene Antikörper gegen Kollagen sowie auch solche gegen Proteine, die in Blutgefäßen vorkommen (Elastin, Laminin und Hämoglobin) positive Reaktionen zeigten. Zur Absicherung der Ergebnisse wurden die Untersuchungen unabhängig in verschiedenen Laboratorien vorgenommen; sie erbrachten weitgehend übereinstimmende Resultate.
Schweitzer et al. versuchten dann massenspektrometrisch Hinweise auf Proteinfragmente in Extrakten aus den Fossilien zu finden. Sie konnten acht Peptide mit insgesamt 149 Aminosäuren finden, die sie aufgrund umfangreicher Analysen als Kollagenfragmente identifizieren konnten. In einem zweiten Labor konnten drei der acht Peptide ebenfalls nachgewiesen und damit das Resultat bestätigt werden. Alle Peptide enthalten Hydroxyprolin, das aus Proteinen von Mikroorganismen nicht bekannt ist. Dies werten die Autoren ebenfalls als Hinweis darauf, dass die Peptide tatsächlich aus Dinosauriergewebe stammen.
In vergleichenden Sequenzanalysen wurden die Kollagenfragmente der beiden Dinosaurier tatsächlich den Archosauria zugeordnet, dazu gehören auch Vögel und Alligatoren, dies stützt ebenfalls die behauptete Authentizität der Peptide.
Mit dieser eindrucksvollen Arbeit, die in Kommentaren auch von Kritikern der ersten Veröffentlichung ausdrücklich geschätzt wird, haben Schweitzer et al. die Anwendbarkeit molekularbiologischer Methoden in der paläontologischen Forschung bestätigt. Die Autoren stellen jedoch auch in dieser Publikation abschließend fest, dass die Chemie dieser außerordentlichen Erhaltung von Peptiden noch nicht verstanden ist und weiterer Forschung bedarf. Denn bisherige Erkenntnisse lassen nicht erwarten, dass Proteinverbindungen über einen Zeitraum von Jahrmillionen erhalten bleiben können.
Quelle:
Schweiter MH, Zheng W, Organ CL, Avci R, Suo Z, Freimark LM, Lebleu VS, Duncan MB, Vander Heiden MG, Neveu JM, Lane WS, Cottrell JS, Horner JR, Cantley LC, Kalluri R & Asara JM (2009) Biomolecular characterization and protein sequence of the Campanian hadrosaur B. canadensis. Science 324, 626-630.
Hinweis: Eine ausführliche Beschreibung und Wertung der Befunde wird in der Herbstausgabe von Studium Integrale Journal veröffentlicht.
Autor dieser News: Harald Binder
© 2009, http://www.genesisnet.info/schoepfung_evolution/n134.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
02.08.08 Gewebereste in Dinosaurier-Fossilien – kontroverse Diskussion der Befunde
Die Aufsehen erregende Publikation von Schweitzer et al. (2005; s. auch Binder 2005, 2007) über flexible Gewebereste aus dem Oberschenkelknochen eines T. rex wurde 2006 auf einer Tagung der Society of Vertebrate Paleontology in Ottawa kontrovers diskutiert (Stokstad 2006). Wir berichteten auch an dieser Stelle („Proteine aus einem fossilen Oberschenkelknochen von Tyrannosaurus rex“, https://www.genesisnet.info/index.php?News=82). Nun haben Kaye und Mitarbeiter (2008) eigene Studien vorgestellt, die alternative Interpretationen nahezulegen scheinen. Die Autoren untersuchten Fossilien aus verschiedenen Lagerstätten mit elektronenmikroskopischen Techniken. Damit konnten sie zunächst die Befunde von Schweitzer et al (2005) bestätigen: auch sie fanden zellähnliche Strukturen innerhalb kleiner Kanälchen. Die zellähnlichen Kugeln fanden sie jedoch ebenfalls in wesentlich schlechter erhaltenen Fossilien und auch in Ammoniten, was eine Identifikation mit Blutzellen nicht unterstützt bzw. sogar ausschließt. Kaye et al. (2008) sprechen sie als Framboide an, das sind himbeerförmige Pyrit (FeS2)-Strukturen, die mit einem Durchmesser von ca. 20 µm im Größenbereich von Zellen liegen. In den mit Fossilien assoziierten Framboiden kann allerdings kein Schwefel nachgewiesen werden. Aus geologischen Studien sind Framboide aus der Umgebung von „Black Smokers“ (mineralreiche heiße Tiefseequellen), Algenmatten und Sedimenten beschrieben, wobei auch Hinweise vorliegen, dass der Schwefelanteil in Folge von Verwitterungsprozesse komplett durch Eisenoxid (FexOy) ersetzt sein kann.
Für ihren Vorschlag, dass die als flexible Gewebereste interpretierten Komponenten aus den T. rex-Fossilien auf Biofilme* zurückzuführen seien, legen die Autoren ebenfalls entsprechende Befunde vor. Kaye et al. interpretieren rissartige Strukturen, die in mikroskopischen Aufnahmen an im Labor aufgebrochenen Fossilien dokumentiert sind, als von Mikroorganismen in einem Biofilm verursachte Rinnen, die auch Verzweigungen aufweisen können. Auch sie konnten durch Auflösen der Mineralbestandteile der Fossilien z.T. flexible, verzweigte, röhrenartige Strukturen isolieren. Eine radiometrische Datierung (14C) ergab jedoch ein modernes Alter. Nach den Vorstellungen von Kaye et al. (2008) sind solche Biofilme, die Kanäle von Gefäßen auskleiden, Ursache für den Befund von flexiblen gewebeartigen Rückständen in den Fossilien nach Auflösung ihrer mineralischen Anteile.
*Biofilme (= eine dünne Schleimschicht, die sich an Grenzflächen in wässrigen Systemen bildet. Sie besteht neben verschiedenen Mikroorganismen (Bakterien, Algen, Pilze, …) aus von diesen ausgeschiedenen Polymeren (Polysaccharide, Proteine, Lipide, Nukleinsäuren), die eine kompakte Matrix bilden in der die Mikroben auch widrige Umweltbedingungen überleben können.)
Sind Framboide mit Biofilmen überzogen, so ist deren Identifikation erschwert.
Die Beteiligung von Biofilmen beim Prozess der Fossilisation wird bereits seit langer Zeit diskutiert, wobei die komplexen Abläufe nach wie vor wenig verstanden sind.
Mit dieser jüngsten Arbeit präsentieren also Kaye et al. mit ihren Befunden den Vorschlag, dass diese hauptsächlich durch mikrobiologische Aktivität (Biofilm) und anorganische geochemische Prozesse (Framboide) zu erklären sind. Mary Schweitzer dagegen verteidigt in ersten Stellungnahmen (Zimmer 2008, Hecht 2008) ihre ursprüngliche Interpretation der flexiblen Bestandteile als erhaltenes Gewebe von Dinosauriern. Sie hält Kaye et al. entgegen, dass mikrobielle Biofilme nicht erklären, warum vogelähnliche Proteinfragmente nachgewiesen werden könnten; auch bezweifelt sie, dass Bakterien die mikroskopischen Röhrchen gebildet haben können.
Man darf also gespannt auf Resultate weiterer Untersuchungen warten. Selbst wenn sich mikrobielle Aktivität als Ursache für die flexiblen Bestandteile der Fossilien in zukünftigen Untersuchungen bestätigen sollte, bleiben viele offene Fragen, z. B.: Wie können sich unter solchen Bedingungen überhaupt ausnehmend gut erhaltene Fossilien bilden? Warum sind Fragmente (ursprünglicher?) Proteine noch nachweisbar? Wenn Pyrit kristallisieren (Framboidbildung) und anschließend mit Biofilm überzogen werden kann, wie kann unter solchen Bedingungen fossile Erhaltung erfolgen?
Die gegenwärtige Diskussion zeigt jedoch einmal mehr, dass man bei der Interpretation fossiler Befunde – auch gerade dort, wo modernste und hochempfindliche Analysenmethoden zum Einsatz kommen – sehr vorsichtig vorgehen und viele Aspekte berücksichtigen muss. Die Diskussion belegt ebenso aufs Neue, wie wenig wir im Grunde genommen die an der Fossilisation beteiligten Prozesse verstanden haben. Hier ist ein unabsehbar weites Feld für weitere Forschung offen.
Quellen
Binder H (2005) Elastisches Gewebe aus fossilen Dinosaurierknochen. Stud. Int. J. 12, 72-73. (online: http://www.si-journal.de/index2.php?artikel=jg12/heft2/sij122-5.html)
Binder H (2007) Proteine aus einem fossilen Oberschenkelknochen von Tyrannosaurus rex. Stud. Int. J. 14, 72-73.
Hecht J (2008) T. rex ‘tissue’ may be bacterial scum. New Scientist.com news service.
http://www.newscientist.com/article/dn14427-t-rex-tissue-may-just-be-bacterial-scum.html?DCMP=ILC-hmts&nsref=news4_head_dn14427
Kaye TG, Gaugler G, Sawlowicz Z (2008) Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms. PLoS ONE 3(7): e2808. doi:10.1371/journal.pone.0002808
Schweitzer MH, Wittmeyer JL, Horner JR & Toporski JK (2005) Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science 307, 1952-1955.
Stokstad E (2006) Soft Tissue in dinosaur fossils? The evidence hardens. Science 314, 920.
Zimmer C (2008) Is dinosaur ‘soft tissue’ really slime? Science 321, 623.
Autor dieser News: Harald Binder
© 2008, http://www.genesisnet.info/schoepfung_evolution/n117.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
21.06.08 Dinohand zu Vogelflügel?
Ein kontroverses Thema der Evolutionsforschung ist die hypothetische stammesgeschichtliche Verbindung zwischen Theropoden-Dinosauriern und Vögeln. Dass die heutigen Vögel der letzte übriggebliebene Ast des Dinosaurier-Stammbaums sind, gilt für viele als nahezu bewiesen. Doch manche Merkmale sperren sich gegen diese Deutung. Dazu gehört der Bau der Dinohand, aus der der Vogelflügel evolutionstheoretisch abgeleitet wird. Henrik Ullrich hat die einschlägigen Arbeiten zu diesem Thema gesichtet und die Ergebnisse kürzlich in der Zeitschrift Studium Integrale Journal (http://www.wort-und-wissen.de/sij) veröffentlicht. Er kommt zum Ergebnis, dass die Befunde eine vielfältige und sehr kontroverse Deutung erlauben und kein klares Bild ergeben. Einmal mehr zeigt sich, dass je nach Perspektive verschiedene, sich widersprechende Homologiezuweisungen resultieren. (Zum Thema „Homologie“ siehe „Ähnlichkeiten in der Morphologie und Anatomie“, https://www.genesisnet.info/index.php?Artikel=41301&Sprache=de&l=1) Ein Schlüssel zur Bestimmung der „korrekten“ stammesgeschichtlich relevanten) Homologie liegt aber nicht vor. Damit liefert der Vergleich von Dinohand und Vogelflügel keinen unabhängigen Beleg für eine gemeinsame Abstammung.
Wir veröffentlichen nachfolgend die Zusammenfassung seines Artikels.
„Zusammenfassung: Die phylogenetische Ableitung der Vögel aus einer Gruppe der Theropoden-Dinosaurier zählt zu einem häufig zitierten und wichtigen Baustein in evolutionären Modellvorstellungen. Betrachtet man aber im Detail die dafür relevanten Befunde, trübt sich das scheinbar klare Bild schnell ein, denn die Befunde erlauben eine vielfältige und kontroverse Deutung. In diesem Artikel werden relevante Daten aus der Embryologie zur Gliedmaßen-Entwicklung bei Wirbeltieren vorgestellt. Deren Bedeutung für Homologie-Betrachtungen und für kausale Erklärungsansätze im Rahmen moderner Evo-Devo Konzepte werden diskutiert. Die große Spannbreite der diesbezüglich in der Literatur äußerst kontrovers vertretenen hypothetischen Vorstellungen führt beim gegenwärtigen Stand des Wissens zu folgenden Schlussfolgerungen:
- Es fehlt ein methodisch sicheres Kriterium, um aus mehreren möglichen Homologie-Zuweisungen auf der Basis embryologischer und paläontologischer Daten die phylogenetisch relevante zu extrahieren. Keine der vorgestellten Ansätze kann deshalb für sich ein höheres Maß an Plausibilität beanspruchen als andere. Die Homologie-Beziehungen zwischen dem Vögelflügel und der vorderen Extremität von Dinosauriern bleiben unter phylogenetischer (= stammesgeschichtlicher) Perspektive auch nach 150 Jahren Evolutionsforschung unklar.
- Die kausalen Erklärungsversuche zur Entstehung des Grundbauplans des Vogelflügels auf der Grundlage der Synthetischen Evolutionstheorie sowie unter Rückgriff auf neue Ansätze der evolutionären Entwicklungsbiologie („Evo-Devo“, https://www.genesisnet.info/index.php?Artikel=41266&Sprache=de&l=2) sind als spekulative Extrapolationen von Details des verfügbaren embryologischen bzw. molekulargenetischen Wissens zu werten.
- Alle vorgelegten Hypothesen zur Evolution des Vogelflügels sehen sich drei defizitären Situationen ausgesetzt: Fehlendes Wissen über Wirkmechanismen bzw. die Wechselbeziehungen von äußeren und inneren Selektionsdrücken auf den evolutionären Umbau der vorderen Extremität und die vielfach noch unverstandenen genetischen und epigenetischen Regulationskaskaden der Gliedmaßen-Entwicklung während der Individualentwicklung bei Wirbeltieren. Drittens muss offen gelassen werden, welche Kausalitäten den phylogenetischen Wandel ontogenetischer Entwicklungsprogramme bedingen und ermöglichen.
- Die wissenschaftliche Klärung der homologen Beziehungen zwischen den Fingerstrahlen an Vogelflügeln und denen bei anderen Landlebewesen (insbesondere Echsen und Säugetieren) und ihre kausale Begründung bleibt eine der größten Herausforderungen für die Evolutionsforschung.
- Das ontogenetische und phylogenetische Modellobjekt „Flügelentwicklung“ ist exemplarisch für grundsätzliche Probleme und Grenzen der evolutionsgeleiteten Hypothesenbildung sowie darauf aufbauender Homologiebetrachtungen.“
Die neue Ausgabe von Studium Integrale Journal kann bei der Studiengemeinschaft Wort und Wissen bezogen werden (https://www.wort-und-wissen.org/publikationen/studium-integrale-journal/).
Autor dieser News: Studiengemeinschaft Wort und Wissen
© 2008, http://www.genesisnet.info/schoepfung_evolution/n114.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
22.05.07 Proteine aus einem fossilen Oberschenkelknochen von Tyrannosaurus rex
In Fossilien, denen ein hohes Alter (ca. 70 Millionen Jahren) zugeordnet wird, haben Wissenschaftler wiederholt flexibles Gewebe isoliert und auch zellähnliche Strukturen nachgewiesen. Jetzt wurden mit leistungsfähigen Analysemethoden Fragmente von Proteinen festgestellt. Dieser Befund steht im Widerspruch zu den etablierten Erfahrungen über die chemische Stabilität von Proteinen. Danach sollten diese biologisch bedeutsamen Makromoleküle nach einer Zeitspanne, die deutlich unter einer Million Jahre liegt, nicht mehr analytisch nachweisbar sein. Zur Lösung dieser Diskrepanz könnten bisher unbekannte Mechanismen zur Langzeitstabilisierung von Polypeptiden beitragen. Solange solche nicht aufgezeigt werden können, stehen die hier vorgestellten Befunde im Widerspruch zu den konventionellen Altersangaben der Fossilien.
Einleitung. Nachdem Schweitzer et al. (2005) aus fossilen Dinosaurierknochen flexible Gewebefragmente isolieren konnten (s. auch Binder 2005), legten sie jetzt Untersuchungen zum Nachweis von Proteinfragmenten aus demselben Material vor (Schweitzer et al. 2007, Asara et al. 2007). Wenn sich die Befunde und deren Interpretation bestätigen, bedeutet das, dass Proteine – zumindest Teile davon – innerhalb von fossilen Knochen unter besonderen Bedingungen aus der unteren Kreide (hier konkret von der Basis der Hell Creek Formation in Ost Montana / USA) bis heute erhalten geblieben und nachweisbar sind. Angesichts des Alters der Probe ist dieser Befund für die Wissenschaftler überraschend.
Die fossilen Überreste stammen von Tyrannosaurus rex (Museum of Rockies specimen, MOR 1125) und umfassen u.a. Schädel, Wirbel, beide Oberschenkel- (Femur) und Schienbeinknochen (Tibia) in außergewöhnlich guter Erhaltung. Die Fossilien waren in mittelkörnigem Sandstein einsedimentiert, der mit feinkörnigem Schlamm (muds) verzahnt ist. Aufgrund dieser Befunde wird als Ablagerungssystem ein Strömungskanal (stream channel) angenommen.
Die Proben. Für die Untersuchungen wurden Proben aus beiden Oberschenkelknochen teilweise demineralisiert, d.h. die Mineralbestandteile wurden durch Zugabe eines Komplexbildners (EDTA) in Lösung gebracht. Die dadurch freigelegten Gewebe würden bei frischen Knochen der extrazellulären Matrix (Osteoid) entsprechen, deren Hauptbestandteil Kollagen I ist.
Als Strukturprotein des Bindegewebes hat Kollagen einen hohen Anteil am Proteingehalt des Körpers; beim Menschen sind ca. 30 % der Eiweißmasse Kollagen. Kollagen besteht typischerweise aus drei Polypeptidketten, die je aus einigen hundert bis mehreren tausend Aminosäuren aufgebaut sind. Typisch für Kollagen ist der hohe Anteil an Glycin – jede dritte Aminosäure ist ein Glycin; auch Prolin und Lysin kommen verhältnismäßig häufig vor.
Nachweis von Kollagen aus Tyrannosaurus-Material. Die Aminosäuresequenz von Kollagen I ist stark konserviert, d.h. die Sequenzen zeigen beim Vergleich zwischen verschiedenen Lebewesen nur geringe Abweichungen. Daher können Antikörper, welche sich in immunologischen Reaktionen spezifisch an Kollagen binden können, mit Kollagenen aus verschiedenen Organismen reagieren.
Bereits 2005 hatten Schweitzer et al. (supporting online material) erste Hinweise auf positive Befunde von immunologischen Tests veröffentlicht. Dabei hatten sie mit Antikörpern gegen Kollagen I vom Huhn und gegen Osteocalcin vom Rind – einem weiteren Protein, das einen hohen Anteil der organischen Knochensubstanz ausmacht – schwache, aber signifikante Reaktionen mit Präparationen aus Dinosaurierknochen erhalten. Kontrollexperimente mit dem umgebenden Sediment oder mit Pflanzenresten, die mit den Fossilien zusammen eingebettet waren, ergaben dagegen keine Reaktion. Das ist ein starker Hinweis dafür, dass das Kollagen aus dem Fossil stammt.
In den jüngsten Arbeiten konzentrierten sich Schweitzer et al. (2007) auf Nachweise für Kollagen, da dies in großen Mengen in Knochenmaterial lebender Organismen vorkommt, vergleichsweise stabil gegen Abbauprozesse ist und mit verschiedenen Methoden analytisch nachgewiesen werden kann. Die Extrakte aus den fossilen Knochen reagierten mit Antikörpern gegen Kollagen I vom Huhn. Die Reaktionen waren im Vergleich zu Extrakten aus frischen Knochen schwach und variierten stark in der Intensität, waren aber mindestens doppelt so stark wie negative Kontrollen. Diese Ergebnisse konnten durch weitere immunohistochemische Tests am fossilen Material bestätigt werden, die an Dünnschnitten von demineralisierten Proben aus dem äußeren Bereich des Knochen (Cortex) und dem Bereich, in dem sich bei lebenden Tieren das Knochenmark (Medulla) befindet, vorgenommen wurden. Wiederholt wurden schwache Antikörperreaktionen nachgewiesen. Diese konnten durch Zugabe von Kollagen vom Huhn unterbunden werden. Wurden die fossilen Proben mit Kollagenase – einem Kollagen abbauenden Enzym – vorbehandelt, war die Immunreaktion deutlich abgeschwächt. Die mittels Fluoreszenzaufnahmen dokumentierten Immunreaktionen waren verglichen mit Negativkontrollen deutlich stärker und im Gewebe lokalisiert. Die Verteilungsmuster waren vergleichbar mit entsprechenden Proben aus frischen Emu-Knochen. Wie bereits bei den ersten Untersuchungen waren die Kontrollexperimente an Sedimentproben negativ. Diese Befunde zeigen, dass die Konzentration von Erkennungsbereichen für die Antikörper (Epitope) in den Tyrannosaurus-Fossilien (MOR 1125) sehr gering ist. Der in situ-Nachweis an Dünnschnitten ist deutlicher im Vergleich zu den Ergebnissen, die mit Extrakten erhalten werden konnten. Insgesamt stützen die Ergebnisse den Befund, dass das Kollagen aus dem Fossil stammt.
Absicherung der Befunde. Zur weiteren Absicherung der Befunde wurden massenspektrometrische Untersuchungen (time-of-flight secondary ion mass spectrometry; TOF-SIMS) an demineralisiertem fossilem Gewebe vorgenommen. Diese Analyse, bei der sowohl Ionen von der Probenoberfläche als auch deren Fragmente mit hoher Auflösung analysiert werden können, ergab Signale für die Aminosäuren Glycin (mit 33 % häufigste Aminosäure in Kollagen), Alanin (ca. 10 % in Kollagen), Prolin, Lysin und Leucin bzw. Isoleucin. Mit anderen MS-Techniken konnten auch hydroxylierte Aminosäuren nachgewiesen werden, welche mit TOF-SIMS nicht nachweisbar waren (Asara et al. 2007). Das Verhältnis von Glycin : Alanin für Kollagen a1 Typ 1 des Huhns wird in der Literatur mit 2,5:1 angegeben. Schweitzer et al. (2007) fanden in ihrer fossilen Probe ein Verhältnis für Glycin : Alanin von 2,6:1. In Sandsteinproben aus dem Sediment, in dem das Fossil eingeschlossen war, konnten keine Aminosäuren oder nur Spuren davon nachgewiesen werden. Es gelang auch noch der Nachweis anderer Stickstoff-(N)-haltiger Molekülfragmente in Fossilproben wie z. B. C7H18N2, welche in Proben aus dem umgebenden Sediment fehlten. In den Proben aus den T. rex-Fossilien waren außerdem Eisen-(Fe)-haltige Ionen enthalten (FeCH, FeCH2, FeCH3). Die Autoren konnten solche Ionen auch in Sedimentproben in der Matrix der Fossilien finden, nicht jedoch in frischen Knochenproben. Sie vermuten, dass es sich dabei um Produkte von Mikroorganismen handelt und interpretieren diese als Hinweise darauf, dass Fe-Ionen an der fossilen Erhaltung von Gewebe durch Auslösen von intra- und intermolekularen Quervernetzungen beteiligt sind.
Weitere Ergebnisse. Die Menge an Protein oder proteinähnlicher Substanz in MOR 1125 ist nach Aussagen der Autoren minimal (0,62 % aus dem Cortex und 1,3 % aus dem Medulla-Bereich, jeweils nach Extraktion und Gefriertrocknung). Schweitzer et al. berichten auch von sehr unterschiedlichen Mikrobereichen in den Fossilien, d.h. die einzelnen Proben unterscheiden sich in ihrem Gehalt an extrahierbarem organischem Material erheblich. Ein interessanter Befund ist auch, dass über den Zeitraum der Untersuchungen die jeweilige Intensität der Signale für die nachgewiesenen organischen Bestandteile immer geringer wurden, d.h. Extraktionen in jüngster Zeit lieferten geringere Mengen an organischem Material. Dies zeigt einen fortschreitenden Abbau des Knochenmaterials seit der Bergung der Fossilien an.
Aus diesem Grund haben Schweitzer et al. umfangreiche Vorsichtsmaßnahmen für die Untersuchungen getroffen. Ein Resultat wurde nur als positiv in die Publikation aufgenommen, wenn es mehrfach (mindestens dreimal) gemessen werden konnte. Außerdem wurden die Untersuchungen in mindestens drei verschiedenen Labors von zahlreichen Wissenschaftlern durchgeführt.
Mögliche Ursachen der außergewöhnlichen Konservierung. Schweitzer et al. (2007a) stellen Befunde vor, die belegen, dass in Sandstein eingebettete Fossilien häufiger Weichteile und Zellstrukturen erhalten. Die Autoren vermuten, dass durch die Poren in der Sandsteinmatrix die flüssigen, den Abbau fördernde Komponenten abgeführt werden können und so aufgrund der Porosität des Sandsteins die Kontaktzeit des fossilisierenden Kadavers mit zerstörenden Flüssigkeiten im Vergleich zu in Ton eingebetteten Organismen reduziert wird (Schweitzer et al. 2007).
Befunde und bisherige Erfahrungen. Schweitzer und Mitarbeiter haben in ihrer jüngsten Publikation (Schweitzer et al. 2007) mit umfangreichen, sorgfältig durchgeführten und durch entsprechende Kontrollexperimente gut abgesicherten Studien deutliche Hinweise für die Erhaltung von Proteinfragmenten aus Fossilien der Kreide vorgelegt. Die Autoren weisen darauf hin, dass ihre Befunde und Interpretationen aufgrund verschiedener Untersuchungen, wie z. B. Lindahl (1993), Bada et al. (1999), Stankiewicz et al. (2000) unerwartet sind. Deren Veröffentlichungen (und auch andere Studien) unterstützen die Vermutung, dass ursprüngliche Gewebebestandteile von Organismen durch Prozesse während der Fossilisierung innerhalb einer relativ kurzen Zeitspanne (die deutlich unter 1 Million Jahren liegt) zerstört oder mindestens so stark verändert sein sollten, dass sie nicht mehr analytisch nachweisbar sind. In diesen Studien waren u. a. Untersuchungen zur Kinetik der Hydrolyse von Proteinen und Nukleinsäuren in wässrigen Lösungen unter verschiedenen Bedingungen extrapoliert worden. Bereits 1992 hatten Muyzer et al. mit verschiedenen immunologischen Reaktionen Hinweise auf Osteocalcin in Knochen von Dinosauriern und anderen fossilen Wirbeltieren aus verschiedenen geologischen Schichten vorgelegt. Der analytische Nachweis sowohl von Glutaminsäure als auch von Peptidbindungen bestätigte das Vorkommen von Protein (-fragmenten). Die Erhaltung des Proteins in den fossilen Knochen war nach Ansicht der Autoren stark von den Umständen der Ablagerung abhängig und nicht einfach vom Alter. Als Reaktion auf den kritischen Einwand, dass aufgrund kinetischer Untersuchungen von Abbaureaktionen ein solcher Befund nicht zu erwarten sei, legten die Autoren eine weitere experimentelle Arbeit vor (Collins et al. 2000). Frisches Knochenmaterial wurde pulverisiert und erhitzt (75 °C, 85 °C, 95 °C) und mittels monoklonaler Antikörper die Abbaukinetik von darin enthaltenem Osteocalcin untersucht. Die Extrapolation der erhaltenen Daten zeigt, dass bei Ablagerungstemperaturen von durchschnittlich unter 10 °C eine Erhaltung von Peptidfragmenten aus dem Mesozoikum möglich erscheint. Eine Kontrolle an fossilen Knochen aus Neolithikum (ca. 6000 Jahre) zeigt jedoch, dass die Erhaltung über weite Bereiche streut und stark vom Zustand des Mineralanteils abhängt. Die Zulässigkeit der Extrapolation erscheint daher zur Zeit noch wenig belastbar und bedarf der Bestätigung durch weitere Studien. Osteocalcin ist in jüngerer Zeit wiederholt in fossilen Knochen unterschiedlichen Alters nachgewiesen worden, nicht zuletzt aus fossilen Knochen von Neandertalern (Nielsen-Marsh et al. 2005).
Schlussfolgerungen. Die widersprüchlichen Befunde, einerseits reproduzierbar nachgewiesene Proteinfragmente in Fossilien mit einem hohen Alter von ca. 70 Millionen Jahren, andererseits die Erfahrung, dass Makromoleküle wie Proteine oder Nukleinsäuren unter kontrollierbaren Laborbedingungen in vergleichsweise kurzer Zeit abgebaut werden, weisen auf die dringende Notwendigkeit weiterer Untersuchungen hin. Solange die Wechselwirkungen zwischen Makromolekülen und Mineralien bzw. deren Oberfläche nicht im Detail verstanden sind und deren Beitrag zur Konservierung unter extremen Bedingungen bzw. über extrem lange Zeiträume mechanistisch aufgezeigt werden können, hinterlässt die Behauptung vom Nachweis ursprünglicher Proteinfragmenten in sehr alten Fossilien (einige Zehner Millionen Jahre) viele offene Fragen. Die Vermutung von Muyzer et al. (1992), dass die fossile Erhaltung von Fragmenten organischer Makromoleküle (Osteocalcin) nicht einfach vom Alter abhängig ist, sollte ebenfalls zu weiterer Forschung motivieren. Ein detailliertes Verständnis der Wechselwirkung zwischen Mineralien und Makromolekülen während der Ablagerungsgeschichte ist dringend erforderlich, um die offenen Fragen gründlicher diskutieren und vielleicht beantworten zu können.
Derzeit kann jedenfalls die zeitliche Zuordnung der geologischen Systeme aufgrund der Diskrepanz zwischen etablierten Erfahrungen aus der Proteinchemie und der behaupteten extremen Stabilität von Proteinen in Fossilien beim gegenwärtigen Kenntnisstand begründet in Frage gestellt werden.
Literatur
Asara JM, Schweitzer MH, Freimark LM, Phillips M & Cantley LC (2007) Protein Sequences from Mastodon and Tyrannosaurus Rex revealed by Mass Spectrometry. Science 316, 280-285.
Bada JL, Wang XS & Hamilton H (1999) Preservation of key molecules in the fossil record: current knowledge and future challenges. Philos. Trans. R. Soc. London Ser. B 354, 77-87.
Binder H (2005) Elastisches Gewebe aus fossilen Dinosaurierknochen. Stud. Int. J. 12, 72-73.
Collins MJ, Gernaey AM, Nielsen-Marsh CM, Vermeer C & Westbroek P (2000) Slow degradation of osteocalcin: Green light for fossil bone protein? Geology 28, 1139-1142.
Lindahl T (1993) Recovery of antediluvial DNA. Nature 365, 700.
Muyzer G, Sandberg P, Knappen MHJ, Vermeer C, Collins M & Westbroek P (1992) Preservation of the bone protein osteocalcin in dinosaurs. Geology 20, 871-874.
Nielsen-Marsh CM, Richards MP, Hauschka PV, Thomas-Oates JE, Trinkaus E, Pettitt PB, KaravaicI, Poinar H & Collins MJ (2005) Osteocalcin protein sequences of Neanderthales and modern primates. Proc. Nat. Acad. Sci USA 102, 4409-4413.
Schweitzer MH, Wittmeyer JL, Horner JR & Toporski JK (2005) Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science 307, 1952-1955. (supporting online material: www.sciencemag.org/cgi/content/full/307/5717/1952/DC1)
Schweitzer MH, Suo Z, Avci R, Asara JM, Allen MA, Arce FT & Horner JR (2007) Analyses of Soft Tissue from Tyrannosaurus rex Suggest the Presence of Protein: Science 316, 277-280.
Schweitzer MH, Wittmeyer JL, Horner JR (2007a) Soft tissue and cellular preservation in vertebrate skeletal elements from the Cretaceous to the present. Proc. R. Soc. London Ser. B 274, 183-197.
Stankiewicz BA, Briggs DEG, Michels R, Collins ME, Flannery MB & Evershed RP (2000) Alternative origin of aliphatic polymer in kerogen. Geology 28, 559-562.
Autor dieser News: Harald Binder
© 2007, http://www.genesisnet.info/schoepfung_evolution/n82.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/