0.3 Grundtypenbiologie
0.3.1.0 Biblische Grundlagen für die Biologie
Der biblische Schöpfungsbericht und andere biblische Schilderungen über Gott als Schöpfer und über sein Handeln in der Geschichte motivieren einige Postulate, die der Grundtypenbiologie und der Design-Theorie zugrunde gelegt werden können. Aus diesen Postulaten kann eine Reihe von Hypothesen abgeleitet werden, die wissenschaftlich prüfbar sind und Forschung anregen.
1.0 Inhalt
In diesem Artikel werden einige Postulate der Grundtypenbiologie und der Design-Theorie zusammengestellt, die vom biblischen Schöpfungsbericht und anderen biblischen Überlieferungen motiviert sind. Aus diesen Postulaten wird eine Reihe von Hypothesen abgeleitet, die wissenschaftlich prüfbar sind.
1.1 Einleitung
Im Artikel |0.1.1.2 Schöpfung und Wissenschaft| wird erläutert, wie unter der Vorgabe von Schöpfung fruchtbare, erkenntnisfördernde Wissenschaft betrieben werden kann. Dabei muss zwischen dem Rahmenparadigma einerseits und konkreten Hypothesen innerhalb dieses Rahmens andererseits unterschieden werden. Mit Hypothesen, die im Rahmen des Schöpfungsparadigmas aufgestellt werden, kann man genauso verfahren wie mit evolutionären Hypothesen: sie sind prüfbar, widerlegbar, regen Forschung an und führen zu neuen Erkenntnissen.
In diesem Artikel sollen einige Konkretisierungen des Schöpfungsparadigmas als Postulate der Grundtypenbiologie und der Design-Theorie formuliert werden. Diese Postulate ergeben sich aus biblischen Aussagen über die Schöpfung und die biblische Urgeschichte oder sind mit ihnen zumindest verträglich. Aus diesen Postulaten können testbare und grundsätzlich falsifizierbare Hypothesen abgeleitet werden, wie weiter unten gezeigt wird. Auf entsprechende Genesisnet-Artikel wird im Einzelnen verwiesen. Zur Grundtypenbiologie vergleiche die Artikel |0.3.2.1 Heutige Grundtypen|, |0.3.2.4 Genetisch polyvalente Stammformen von Grundtypen| und |0.3.2.6 Kritik an der Grundtypenbiologie|, zur Design-Theorie siehe Artikel |0.4.1.1 Einführung in „Intelligent-Design“| und |0.4.1.2 Kontroverse um „Intelligent-Design“|.
1.2 Postulate der Grundtypenbiologie
- Alle Lebewesen waren ursprünglich optimal konstruiert.
Dieses Postulat ergibt sich aus der biblischen Selbstoffenbarung Gottes über sein Schöpfungshandeln. Vielfach wird die Kraft und die Weisheit Gottes als Schöpfer gepriesen. Daraus darf man schließen, dass Gott keinen „Pfusch“ erschaffen hat. Außerdem bezeichnet Gott selber nach dem Zeugnis der Heiligen Schrift die Gesamtheit seiner Schöpfung als „sehr gut“ (Genesis 1,31). Kritiker der biblischen Schöpfungslehre haken an dieser Stelle oft ein; Artikel |0.4.2.1 Argumente gegen Design| befasst sich mit dieser Kritik. Theoretisch wäre ein Schöpfer natürlich frei, auch Pfusch zu erschaffen; doch dies soll in hier ausgeschlossen werden, da dies aufgrund zahlreicher biblischer Aussagen über Gottes Weisheit gepriesen als unwahrscheinlich gelten kann.
- Alle Lebewesen starteten ihre Existenz als polyvalente, abgegrenzte Grundtypen.
Das Postulat der Abgrenzbarkeit der Grundtypen (vgl. Abb. 82) ergibt sich aus dem biblischen Schöpfungsbericht, wonach Pflanzen und Tiere „nach ihrer Art“ erschaffen wurden (vgl. |0.1.1.1 Biblische Grundlagen der Schöpfungslehre| und Junker 1994, 218ff.). Die biblischen Texte definieren „Art“ nicht. In der Grundtypenbiologie wird dazu jedoch ein konkreter Vorschlag gemacht, der eine experimentelle Prüfung von Grundtypgrenzen erlaubt (|1.3.1.4 Artbegriffe|, |0.3.2.1 Heutige Grundtypen|). Demnach gehören alle biologischen Arten, die durch Kreuzungen direkt oder indirekt miteinander verbunden sind, zu einem Grundtyp, wenn nachgewiesen werden kann, dass während der Ontogenese das Erbgut beider Eltern ausgeprägt wird. Details dazu und Begründungen für diesen Ansatz werden in Scherer (1993) erläutert (vgl. auch Junker & Scherer 2001, Abschnitte II.3 und VII.17.3).
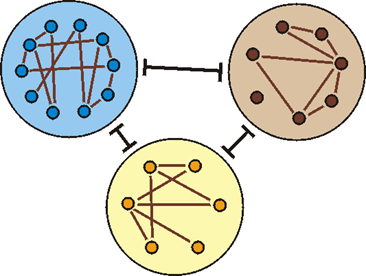
Abb. 82: Bei den bisher untersuchten Tier- und Pflanzengruppen konnten deutliche Grundtypgrenzen festgestellt werden. Innerhalb der Grundtypen (große Kreise) sind die verschiedenen Biospezies (kleine Kreise) jedoch eng verbunden.
Die Vorgabe der Polyvalenz ist zwar durch Kenntnisse der Evolutionsmechanismen motiviert, aber auch naheliegend, wenn man die biblischen Schilderungen voraussetzt, denn danach stammen alle Menschen von einem einzigen Menschenpaar ab. Das ist angesichts der heutigen Vielfalt nur bei einer Polyvalenz der Ursprungsform möglich. Polyvalenz beinhaltet darüber hinaus auch das Potential, zukünftigen Anforderungen durch entsprechende Flexibilität begegnen zu können, was als Design-Merkmal gewertet werden kann und daher im Rahmen eines Schöpfungsparadigmas naheliegend ist.
Wichtig zum Verständnis des Begriffs „Polyvalenz“ ist, dass er nicht nur einen anfänglichen ausgeprägten Polymorphismus [= Vielgestaltigkeit, genetische Vielseitigkeit] meint, sondern auch ein Variationspotential einschließt. Grundtypen besitzen daher eine aktuelle sowie eine potentielle Vielfalt.
Zur Polyvalenz von Grundtypen tragen bei:
- ein ursprünglicher genetischer (und damit auch phänotypischer) Polymorphismus
- ein großes Modifikationspotential. Ein Beispiel ist der Wasser-Hahnenfuß: er hat drei Blatttypen, die je nach Umgebung (Luft, Wasser, Wasseroberfläche) ausgebildet werden. Dies birgt auch ein Potential für Artbildung (vgl. Junker 1993, 42f.).
- programmierte Variabilität, z. B. ein vorprogrammiertes Mutationsspektrum. Interessant ist in diesem Zusammenhang das „Gesetz der rekurrenten Variation“ (nach Lönnig; vgl. |1.3.2.1 Mutation|).
- Die Lebewesen sind nur im Rahmen der vorgegebenen Polyvalenz zu Variation fähig.
Grenzen der Variationsfähigkeit zu postulieren, ist naheliegend, weil man davon ausgehen kann, dass die Schöpfungseinheiten des Lebens (also die Grundtypen) nicht nur am Anfang schöpfungsgemäß getrennt waren, sondern als solche auch nach vielen Generationen erkennbar bleiben. Bei einer beliebigen oder sehr weitgehenden Wandelbarkeit wäre das nicht der Fall. Hier liegt ein offenkundiger Gegensatz zum Evolutionsparadigma vor. Die Plausibilitäten der gegensätzlichen Erwartungen innerhalb von Evolutionstheorien und Grundtyptheorien kann anhand von Daten getestet werden.
Die Existenz fertiger, abgrenzbarer Grundtypen sowie Grenzen ihrer Veränderlichkeit wird aus biblischer Sicht auch indirekt dadurch begründet, dass eine evolutionäre Entstehung aus zahlreichen theologischen Gründen ausgeschlossen werden muss (vgl. |0.5.1.2 Die biblische Urgeschichte im Neuen Testament| und |0.5.1.3 Evolutionsmechanismen als Schöpfungsmethode?|).
- Es gibt in großem Ausmaß inkongruente Merkmalskombinationen bei den verschiedenen Grundtypen (modulares Baukastensystem)
Dieses Postulat ist naheliegend, weil der Urheber der Grundtypen frei ist, Merkmale beliebig zu kombinieren, solange dies funktionell Sinn macht. Die Wiederverwendung von Bauteilen ist sozusagen ingenieurstechnisch zweckmäßig und bietet darüber hinaus eine „unifying message“ (ReMine 1993), also einen Hinweis darauf, dass es nur einen einzigen Urheber gibt, was dem Monotheismus entgegenkommt. Andererseits macht die Wiederverwendung von Bauteilen aufgrund zahlloser Konvergenzen Evolutionskonzepten Probleme (vgl. |1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie|). Zwingend aus dem biblischen Schöpfungsbericht ergibt sich dieses Postulat jedoch nicht; es ist aber naheliegend, weil Gott als planvoll handelnder Schöpfer beschrieben wird.
Man könnte hier entgegnen, dass auch andere Postulate formuliert werden könnten. Das ist richtig, doch das ist kein Spezifikum des Schöpfungsparadigmas, wie in Artikel |1.1.3.4 Evolutionsparadigma und Naturwissenschaft| gezeigt wird. Die hier formulierten Postulate orientieren sich wie erläutert am biblischen Schöpfungszeugnis. Sie sind bei dessen Vorgabe nahelegend, wenn auch nicht alle gleichermaßen zwingend. Sie sind genügend konkret gefasst, um sie Prüfungen zugänglich zu machen, und darauf kommt es hier an. Es sei an die Unterscheidung „Schöpfungsparadigma“ bzw. „Evolutionsparadigma“ und die davon motivierten Hypothesen bzw. Theorien erinnert (s. o. und |0.1.1.2 Schöpfung und Wissenschaft|).
1.3 Schlussfolgerungen und Hypothesen
Aus diesen vier Postulaten können die nachfolgenden Schlussfolgerungen gezogen und als Hypothesen im Rahmen des Schöpfungsparadigmas aufgestellt werden, die testbar sind. Wie diese Tests durchgeführt werden, wird in den angegebenen Artikeln gezeigt.
Taxonomie:
- Grundtypen sind nach dem vorgelegten Kriterium abgrenzbar; es gibt unter den Lebewesen eine Diskontinuität auf Grundtyp-Ebene (siehe |0.3.2.6 Kritik an der Grundtypenbiologie|). Diese Schlussfolgerung ergibt sich aus dem zweiten Postulat.
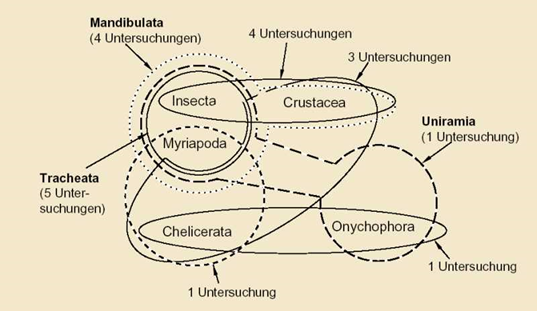
Abb. 206: Ähnlichkeitsbeziehungen unter Lebewesen können als Netzwerk dargestellt werden: Verwandtschaftsverhältnisse der Arthropoden (Gliederfüßer). Je nach zugrunde gelegten Merkmalen ergeben sich unterschiedliche Gruppierungen. Die entsprechenden Untersuchungen stammen alle aus den 1990er Jahren. (Nach Wägele 2001, 102). Wägele kommentiert diese Abbildung: „Ergebnisse neuerer Analysen sind untereinander nicht kompatibel, mehrere davon oder alle müssen demnach fehlerhaft sein.“
- Oberhalb der Grundtyp-Ebene gibt es inkongruente Mosaikformen; es sind teilweise Netzstrukturen der Ähnlichkeitsbeziehungen zu erwarten (das wäre eine Widerspiegelung eines Baukastensystems; Abb. 206 zeigt ein Beispiel), keine widerspruchsfreien Baumstrukturen (siehe |0.3.3.1 Mosaikformen als Grundtypen und Baukastensysteme|). (Viertes Postulat)
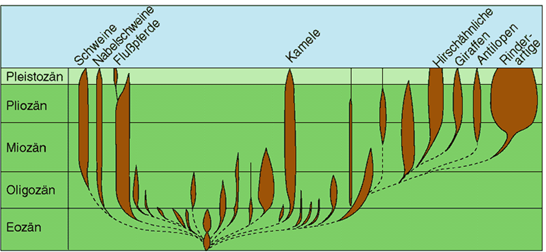
Abb. 87: Fossildokumentation der Paarhufer-Familien. Die Überlieferung fossiler Reste reißt ohne Verbindung durch Übergangsformen zwischen den Familien ab (gestrichelte Linien). Im Tierreich sind Familien häufig identisch mit Grundtypen. Die meisten Familien der Paarhufer sind ausgestorben (diese sind nicht beschriftet). (Nach Romer 1968)
Evolutionsmechanismen:
- Variationsmöglichkeiten bewegen sich innerhalb von Grundtypen. Es gibt nur Mikroevolution (vgl. |1.3.1.3 Mikro- und Makroevolution|. Dies passt zum dritten Postulat der Abgrenzbarkeit von Grundtypen.
- Innerhalb von Grundtypen sind aufgrund der Polyvalenz häufig netzartige Beziehungen unter den einzelnen Arten zu erwarten (zweites Postulat); wie ausgeprägt diese sind, hängt u. a. vom Ausmaß der ursprünglichen Polyvalenz ab.
Fossilien:
- Auf der Grundtyp-Ebene sind systematisch Fossillücken zu erwarten (vgl. |0.3.2.2 Fossile Grundtypen|; ein Beispiel zeigt Abb. 87). Oberhalb der Grundtypebene kann es wiederum inkongruente Mosaikformen geben. (Diese sind evolutionstheoretisch u. U. als Übergangsformen zwischen höheren Taxa interpretierbar, z. B. Archaeopteryx, vgl. |1.7.2.4 Entstehung der Vögel|.) (Zweites Postulat)

Abb. 186: Kambrische Explosion. Kein Baum, sondern ein Strauch ohne Hauptäste und ohne Verzweigungen: das fossil dokumentierte Auftreten der wichtigsten Tierstämme. Die Strichlierung steht für das Fehlen von Fossilfunden.
Intelligent Design:
- Es ist zu erwarten, dass Lebewesen Eigenschaften aufweisen, die auf einen schöpferischen Ursprung hinweisen (siehe |0.4.1.1 Einführung in „Intelligent-Design“|). (Erstes Postulat)
- Es ist zu erwarten, dass es keine primären Konstruktionsfehler bei den Lebewesen gibt. (Mit „primär“ ist gemeint, dass Mängel im Rahmen der Grundtypenbiologie nicht durch Degenerationen aufgrund schädlicher Mutationen erklärt werden können, sondern von vornherein – zu Beginn der Existenz der Grundtypen – vorhanden gewesen sein müssen; siehe |0.4.2.1 Argumente gegen Design|.) (Erstes Postulat)
1.4 Wann sind die Grundtypen entstanden?
Ein wichtiger Aspekt fehlt in den bisherigen Ausführungen, nämlich die Frage, wann die Grundtypen entstanden sind. Nach dem biblischen Schöpfungsbericht sind alle Grundtypen in der Schöpfungswoche entstanden. Geologisch gesehen bedeutet das Gleichzeitigkeit.
Wir formulieren daher als 5. Postulat:
- Alle Grundtypen sind geologisch gesehen gleichzeitig entstanden.
Im Rahmen der biblischen Schöpfungslehre kann daraus folgende Erwartung abgeleitet werden:
Geologie/Paläontologie
- Die Fossilüberlieferung der Grundtypen zeigt keine auffälligen Regelhaftigkeiten (vgl. |1.7.1.2 Die geologischen Systeme|, Abb. 186). Offenkundig entsprechen viele paläontologische Befunde nicht dieser Erwartung. Die hier formulierte Erwartung des Fehlens von Regelhaftigkeiten in der Fossilüberlieferung ist allerdings anfechtbar, wenn auch die weiteren biblischen Aussagen über die Geschichte der Lebewesen berücksichtigt werden, insbesondere die Schilderung über die weltweite Sintflut. Genau dies könnte in einer biblisch-urgeschichtlichen Geologie ein wesentlicher Schlüssel zur korrekten Deutung der Fossilüberlieferung sein. Daher wird im Rahmen der biblischen Sicht der Schöpfung und der Geschichte der Lebewesen nach Ursachen gesucht, die zu diesem zunächst nicht zu erwartenden Befund der fossilen Regelhaftigkeiten führen. Ansätze dazu diskutiert Stephan (2002). Zweifellos sind bei der Verknüpfung der Grundtypenbiologie mit der Erdgeschichte wesentliche Fragen offen. Eine plausible Gesamtdeutung der Daten im Rahmen einer biblisch-urgeschichtlichen Geologie steht bislang aus. Die damit verbundenen komplexen Fragen erfordern eine eigene Behandlung und werden in diesem Artikel nicht thematisiert.
Für Kritiker, die der biblischen Überlieferung keine Bedeutung beimessen, muss eine solche Vorgehensweise fremd und vielleicht auch unsinnig erscheinen. Werden die biblischen Schilderungen jedoch als Offenbarung Gottes betrachtet, dienen sie als Schlüssel zum Verständnis, als Leitschienen für die Rekonstruktion der Geschichte der Lebewesen.
1.5 Literatur
Junker R (1993) Prozesse der Artbildung. In: Scherer S (Hg) Typen des Lebens. Berlin, S. 31-45.
Junker R (1994) Leben durch Sterben? Schöpfung, Heilsgeschichte und Evolution. Neuhausen-Stuttgart.
Junker R & Scherer S (2001) Evolution – ein kritisches Lehrbuch. Gießen.
ReMine WJ (1993) The Biotic Message. Evolution versus Message Theory. Saint Paul, Minnesota.
Scherer S (1993) basic types of life. In: Scherer S (Hg) Typen des Lebens. Studium Integrale. Berlin, S. 11-30.
Stephan M (2002) Der Mensch und die geologische Zeittafel. Holzgerlingen.
Autor: Reinhard Junker, 16.07.2005
© 2005, https://www.genesisnet.info/schoepfung_evolution/i1221.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
0.3.1.1 Biblische Schöpfungslehre und Grundtypenbiologie
Die taxonomische Einheit des Grundtyps kann als „Schöpfungseinheit“ („geschaffene Art“) interpretiert werden. Tatsächlich zeigt sich, dass Arten desselben Grundtyps kreuzungsmäßig eng verbunden sind, zu Nachbargrundtypen dagegen eine „Kreuzungslücke“ auftritt. Dies passt zur Deutung von Grundtypen als Schöpfungseinheiten. Innerhalb der Grundtypen kann es erhebliche Variabilität geben.
0.3.2.1 Heutige Grundtypen
1.0 Inhalt
In diesem Artikel wird erklärt, weshalb Grundtypen als Schöpfungseinheiten interpretiert werden können. Einige Beispiele von Grundtypen werden kurz vorgestellt und Schwächen und Stärken des Grundtypmodells angesprochen. Außerdem wird die Variabilität innerhalb von Grundtypen thematisiert.
1.1 Einleitung
Im Artikel |1.3.1.4 Artbegriffe| wurde als taxonomische Einheit der Grundtyp eingeführt. Danach gehören alle Biospezies [= biologische Arten: dazu gehören alle Individuen, die unter natürlichen Bedingungen fruchtbar kreuzbar sind] zu einem Grundtyp, die durch Kreuzungen direkt oder indirekt miteinander verbunden sind, unabhängig davon, ob die Mischlinge fruchtbar sind oder nicht und ob sie im Freiland auftreten oder nach künstlicher Kreuzung erzeugt wurden. (Bei der Biospezies-Definition wird dagegen zusätzlich gefordert, dass die Mischlinge im Freiland auftreten und fruchtbar sind.)
1.2 Beispiele von Grundtypen
Beispielsweise bilden die Pferdeartigen – Pferd, zwei Eselarten und drei Zebraarten – einen Grundtyp (Abb. 80). Die sechs Arten wurden bis auf eine Ausnahme direkt miteinander erfolgreich gekreuzt (im Sinne des Grundtypkriteriums). Die beiden bisher nicht direkt gekreuzten Arten sind aber mehrfach indirekt durch Kreuzungen miteinander verbunden. Daher ist die Zusammengehörigkeit zu einem einzigen Grundtyp gut belegt.

Abb. 80: Der Grundtyp der Pferdeartigen in einer Kreuzungsmatrix zusammengefasst. Punkte stehen für gelungene Kreuzungen nach der Grundtypdefinition (d. h. es entstanden Mischlinge, in denen das Erbgut beider Eltern ausgeprägt wurde, meistens waren die Mischlinge steril). Fast alle Mitglieder sind durch Kreuzung direkt verbunden, alle sind mehrfach indirekt verbunden. Es gibt keine Mischlinge zwischen einem Pferdeartigen und einem anderen Säugetier. Damit ist der Grundtyp der Pferdeartigen deutlich erkennbar und gegen andere Grundtypen abgegrenzt.

Abb. 28: Einige Arten des Grundtyps der Entenartigen. Obere Reihe: Kanadagans, Rothalsgans, Trompeterschwan, Schwarzhalsschwan, untere Reihe: Mittelsäger, Stockente, Mandarinente, Laysan-Stockente, Afrikanische Zwergglanzente.
Grundtypen sind in vielen Fällen aber deutlich umfangreicher. Beispielsweise gehören zum Grundtyp der Entenvögel (Abb. 28) ca. 150 Biospezies. In der Literatur sind weit über 400 Kreuzungen zwischen den verschiedenen Arten, Gattungen und Unterfamilien der Entenartigen gemeldet worden, jedoch niemals über die Familiengrenze hinaus. Eine genauere Analyse zeigt, dass 126 der 149 Arten der Entenvögel direkt oder indirekt durch Kreuzungen verbunden sind. Dabei hängen mit Ausnahme der Spaltfußgänse alle Triben [= Gattungsgruppe, Klassifikationsebene zwischen Familie und Gattung] über Kreuzungen miteinander zusammen. Ein Beispiel für eine Kreuzung zwischen zwei verschiedenen Triben ist in Abb. 81 dargestellt. Da 23 Entenarten noch nicht mit anderen gekreuzt wurden, lassen sich diese Arten nach der Grundtypdefinition noch nicht endgültig dem Enten-Grundtyp zuordnen. Die gesamte Biologie dieser Arten lässt es aber mit Ausnahme von der Spaltfußgans als sehr wahrscheinlich gelten, dass sie mit zum Grundtyp der Entenvögel gerechnet werden können. Die Unterfamilie der Spaltfußgänse wird jedoch nicht zum Grundtyp der Entenvögel gezählt, weil nicht nur keine Kreuzungen bekannt sind, sondern auch morphologische (Gestalt), anatomische (Körperbau), verhaltensbiologische und molekularbiologische Befunde die Spaltfußgans scharf von allen anderen Entenvögeln abheben.

Abb. 81: Kreuzung zwischen Eiderente (unten) und Brandgans (Mitte). Diese Tiere sind innerhalb der Entenvögel nur entfernt verwandt. Der Mischling ist oben abgebildet.
1.3 Abgrenzbarkeit von Grundtypen
Bisher wurden etwa 20 Grundtypstudien durchgeführt (einige Grundtypen sind in Abb. 86 aufgelistet). Diese Untersuchungen haben bislang ausnahmslos gezeigt, dass sich Gruppen von Lebewesen anhand der Grundtypkriterien deutlich voneinander abgrenzen lassen. Grundtypen sind als solche identifizierbar und weisen aufgrund der Kreuzbarkeit eine erkennbare Lücke zu Nachbargrundtypen auf.

Abb. 86: Anzahlen von Biospezies pro Grundtyp in Auswahl. Innerhalb von Grundtypen zeigt sich eine enge Verbundenheit durch zahlreiche Mischlinge (s. o. Abb. 80). Zwischen „benachbarten“ (d. h. taxonomisch nahestehenden) Grundtypen hingegen wurden bisher keine zweifelsfreien Mischlinge beobachtet. Das heißt: Bezüglich der Kreuzbarkeit sind auffällige Einschnitte zwischen Gruppen von Lebewesen erkennbar, die gerade dadurch als Grundtypen erkannt werden können (Abb. 82).
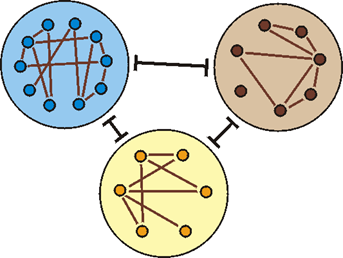
Abb. 82: Bei den bisher untersuchten Tier- und Pflanzengruppen konnten deutliche Grundtypgrenzen festgestellt werden. Innerhalb der Grundtypen (große Kreise) sind die verschiedenen Biospezies (kleine Kreise) jedoch eng verbunden.
Die Erkennbarkeit und Abgrenzbarkeit von Grundtypen wird also anhand von zwei Befunden festgemacht: 1. Innerhalb des Grundtyps treten zahlreiche Mischlinge auf (enge innere Verbindung). 2. Grundtypen, die von ihrer Morphologie (Bau, Gestalt) her benachbart sind, sind durch keine einzige Kreuzung verbunden (Abb. 82).
Die zahlreichen Mischlinge zeigen, dass innerhalb von Grundtypen gleiche Grundmuster morphogenetischer Gene [= Gene, die bei der Formbildung des Organismus besondere steuernde Funktion haben“] vorliegen. Dies sollte sich in grundtypspezifischen phänotypischen [= das äußere Erscheinungsbild betreffend] Merkmalen äußern. In der Tat zeichnen sich die Grundtypen durch typische morphologische Merkmale aus, beispielsweise die Entenartigen durch den typischen Entenschnabel (vgl. Abb. 28), die Kernobstgewächse durch die typische Apfelfrucht (Abb. 83) usw. Auch nach biochemischen Merkmalen lassen sich Grundtypen in manchen Fällen abgrenzen.
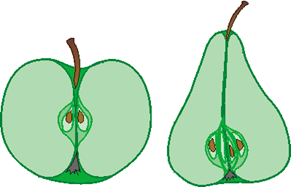
Abb. 83: Die typische Apfelfrucht als besonderes Kennzeichen der Kernobstgewächse.
1.4 Grundtypen als Schöpfungseinheiten
Im Rahmen von Schöpfungsvorstellungen werden diese empirischen Befunde in einen Deutungsrahmen gestellt, indem Grundtypen als Schöpfungseinheiten aufgefasst werden. Es bietet sich an, Grundtypen mit „geschaffenen Arten“ gleichzusetzen (Abb. 84). Jedoch hat eine solche Annahme nur hypothetischen Charakter. So ist nicht genau zu klären, was die biblischen Texte unter „geschaffenen Arten“ verstehen. Außerdem könnten neue Ergebnisse der biologischen Forschung eine andere Lösung nahelegen. Doch bislang lassen sich die Ergebnisse der Grundtypforschung befriedigend im Sinne der o.g. Gleichsetzung deuten. Die deutliche Abgrenzbarkeit von Grundtypen kann als Hinweis für die Existenz getrennt geschaffener Einheiten gewertet werden.

Abb. 84: Grundtypen als Schöpfungseinheiten, die sich als genetisch vielseitige Stammformen in zahlreiche Arten ausdifferenziert haben. Im Rahmen der Schöpfungslehre resultiert daraus ein Forschungsprogramm: Weitere Grundtypstudien müssen die Tragfähigkeit des Grundtypkonzepts und darüber hinaus die angesprochene Identifikation von Grundtypen mit Schöpfungseinheiten stützen oder in Frage stellen. Dazu gehört auch der Test, grundtypübergreifende Kreuzungen zu versuchen. Bei Gräsern wurde dies aus züchterischem Interesse schon vielfach durchgeführt; dabei sind fast alle Tribus-übergreifenden Kreuzungen (über ein Dutzend) fehlgeschlagen.
Wie verhalten sich Biospezies und Morphospezies zu Grundtypen? Die eingebürgerten und teilweise bewährten Artbegriffe werden durch den Grundtypbegriff keineswegs überflüssig. Vielmehr können genetisch und morphologisch begründete Arten als Unterteilungen von Grundtypen verwendet werden. Biospezies können nach wie vor als Grundeinheit der Taxonomie gelten, zu der alle Individuen gehören, die durch fruchtbare Kreuzbarkeit miteinander verbunden sind (genetischer Aspekt). Darüber steht die Gattung als Zusammenfassung der Biospezies, die einander deutlich ähneln (morphologischer Aspekt). Schließlich werden alle diese Arten zu Grundtypen zusammengefasst, wenn sie das gleiche Grundmuster der Embryonalentwicklung aufweisen (Formbildung; morphogenetischer Aspekt).
Nach bisherigen Studien liegt das Grundtypniveau auf dem Niveau der Familie oder zwischen Familie und Gattung.
1.5 Variation innerhalb von Grundtypen
Die Vielfalt innerhalb von Grundtypen ist erstaunlich. In Abb. 28 ist eine ganze Reihe von Entenvögeln abgebildet. Man kann sie trotz ihrer Verschiedenheit als Entenvögel erkennen, etwa am typischen Schnabelbau. Doch auf der anderen Seite fällt auch auf, wie verschieden in Größe, Form, Färbung oder Lebensweise die Arten dieses Grundtyps sind. Die Säger ernähren sich von Fischen und sind entsprechend angepasst, während die Schwimmenten kleinste Organismen durch eine siebartige Einrichtung des Schnabels aus dem Wasser seihen. Auch bei Pflanzen wird eine außerordentliche Verschiedenheit der Arten innerhalb eines Grundtyps beobachtet. Beispielsweise sind die Weiden (Gattung Salix) durch mehrfache indirekte Kreuzungen miteinander verbunden: vom hochalpinen Zwergstrauch bis zum üppigen Tieflandsbaum (Abb. 85).
Abb. 85: Enorme Variabilität innerhalb des Grundtyps der Weiden.
Die Quellen für diese Variabilität werden im Artikel |0.3.2.4 Genetisch polyvalente Stammformen von Grundtypen| besprochen.
Die bisherigen Untersuchungen zeigen, dass die Artenzahlen (in Biospezies gerechnet) von Grundtypen sehr unterschiedlich sein können (Abb. 86). Generell scheinen Pflanzengrundtypen deutlich artenreicher zu sein als Tiergrundtypen. Ob innerhalb der Tiergrundtypen weitere systematische Unterschiede in den Grundtypgrößen auftreten, ist noch nicht absehbar. Ein auch ökologisch sinnvoller Faktor könnte das Körpergewicht sein: Grundtypen mit einem durchschnittlich größeren Körpergewicht tendieren zu geringeren Artenzahlen. Doch die Prüfung dieser Hypothese wird erst durch die Erforschung zahlreicher weiterer Grundtypen möglich sein.
1.6 Schwächen und Stärken des Grundtypmodells“
Das Grundtypmodell hat drei Hauptschwächen. Zum einen ist es bisher erst an wenigen Organismengruppen wirklich fundiert getestet worden. Damit kann es vorerst auch nur als eine Arbeitshypothese betrachtet werden. Der weitere Gang der biologischen Forschung wird zur Bestätigung, zur Revision oder auch zur Verwerfung dieser Hypothese führen. Die Erarbeitung einer vollständigen Taxonomie auf Grundtypbasis steht aus.
Eine zweite Schwäche der Grundtypdefinition liegt in den zuweilen auftretenden praktischen Schwierigkeiten, im konkreten Fall eine Kreuzung zu vollziehen. Der Kreuzung verschiedener Arten sind bei Tieren von der Verhaltensbiologie her Grenzen gesetzt, und eine künstliche Befruchtung ist zwar manchmal leicht möglich, aber z. B. im Falle seltener oder schwer zu haltender Tiere außerordentlich schwierig.
Eine dritte Schwäche des Grundtypmodells liegt in der Definition. Die experimentelle Kreuzungsanalyse zeigt, dass die Embryonalentwicklung in manchen Fällen nicht über die ersten Zellteilungsstadien hinauskommt. In anderen Fällen bricht sie beim Stadium der Neurulation ab, in wieder anderen Fällen kommt es noch später zu Fehlentwicklungen. Wann genau kann man nun von einer erfolgreichen Kreuzung sprechen? Um dieses Problem zu lösen, wird ein weiterer Aspekt der Grundtypdefinition eingeführt:
„Zwei Individuen gehören zum gleichen Grundtyp, wenn die Embryogenese eines Mischlings über die maternale [= mütterliche] Phase der Entwicklung hinausführt und eine koordinierte Expression von väterlichen und mütterlichen morphogenetischen Genen beinhaltet.“
Damit sollen verschiedene Fälle ausgeschlossen werden. Beispielsweise können Eier von bestimmten Wirbellosen durch einen Nadelstich zur Teilung angeregt werden – dann entsteht ohne Beteiligung eines väterlichen Genoms ein lebensfähiger Organismus (Parthenogenese = Jungfernzeugung). Die Funktion eines Nadelstiches könnte auch durch ein Spermium übernommen werden, dessen DNS aber nicht an der weiteren Entwicklung des Eis teilnimmt. Dann spricht man von einer scheinbaren Befruchtung, doch werden die Gene der väterlichen Keimzelle niemals aktiviert und gehen schließlich im Laufe der Zellteilungen verloren. In diesen Fällen ist gar keine Aussage über die Grundtypzugehörigkeit der Eltern möglich. Durch die o.g. Ergänzung der Grundtypdefinition werden Fehlschlüsse bei der Grundtypbestimmung verhindert.
Die bisher erfolgten Grundtypbeschreibungen beruhen jedoch ausschließlich auf erfolgreichen Kreuzungen mit lebensfähigen Nachkommen. Die erwähnte Ergänzung der Definition wird wahrscheinlich nur in sehr speziellen Fällen zur Anwendung kommen.
Eine Stärke des Modells ist die prinzipielle Prüfbarkeit. Bei Tieren und Pflanzen liegen Tausende von Kreuzungsberichten in der Literatur vor, welche ausgewertet werden können. In kritischen Fällen kann künstliche Befruchtung trotz des o.g. experimentellen Aufwandes auch zur Entscheidung herangezogen werden, und damit ist die systematische Kategorie des Grundtyps als einzige Typdefinition der gesamten Biologie experimentell prüfbar. In den Fällen, in denen man die Grundtypdefinition an konkreten Beispielen geprüft hat, waren die Ergebnisse bisher ermutigend. Durch die Entwicklung der Molekularbiologie wird es vielleicht möglich sein, die einen Grundtyp kennzeichnenden Genmuster zu finden – doch ist dies derzeit nicht im Einzelnen absehbar.
1.7 Zusammenfassung
Die taxonomische Kategorie des Grundtyps kann im Gegensatz zu anderen Artbegriffen experimentell geprüft werden. Durch Kreuzungsanalysen konnten bisher rund 20 verschiedene Grundtypen im Tier- und Pflanzenreich bestimmt werden. Zwischen diesen Grundtypen und ihren nächststehenden Verwandten existieren klare Unterschiede.
Die Grundtypenbiologie harmoniert mit der herkömmlichen Taxonomie und ergänzt sie auf sinnvolle Weise: Angehörige einer Biospezies sind unter natürlichen Bedingungen durch Kreuzungen verbunden (genetische Ebene), alle Biospezies, welche einander deutlich ähneln, gehören zu einer Gattung (morphologische Ebene) und alle Gattungen, die grundsätzlich kreuzbar sind, gehören zu einem Grundtyp (morphogenetische Ebene).
Die Grundtypentaxonomie ist als Konzept der empirisch arbeitenden Biologie unabhängig von Ursprungsvorstellungen, eignet sich aber gut als systematische Kategorie im Rahmen der Schöpfungslehre.
Literaturhinweis
Junker R & Scherer S (2013) Evolution – ein kritisches Lehrbuch. Gießen, 7. Auflage, Kap. VII.16.3.
Autor: Reinhard Junker, 11.10.2013
© 2013, https://www.genesisnet.info/schoepfung_evolution/i1241.php
0.3.2.2 Fossile Grundtypen
Nach dem Kreuzungskriterium können heute lebende Grundtypen voneinander abgegrenzt werden. Viele Arten sind nur fossil bekannt. Lassen sich auch fossile Arten als Grundtypen zusammenfassen? Und bestätigen sich Grundtypgrenzen heutiger Grundtypen bei Berücksichtigung fossiler Formen?
1.0 Inhalt
In diesem Artikel wird erklärt, wie unter nur fossil erhaltenen Organismen Grundtypen erkannt werden können. Außerdem wird der Frage nachgegangen, ob sich die Grenzen zwischen heutigen Grundtypen bei Hinzunahme fossiler Formen bestätigen.
1.1 Einleitung
Im Artikel |0.3.2.1 „Heutige Grundtypen“| wurde beschrieben, dass und wie Grundtypen unter der heutigen Vielfalt der Arten erkannt werden können. Unter den heute lebenden Arten lassen sich Grundtypen mit Hilfe des Kreuzungskriteriums als zusammenhängende Einheiten erkennen und gegen Nachbargrundtypen abgrenzen.
Das definierende Kriterium für die Erkennung von Grundtypen ist die Kreuzbarkeit wie im Artikel über |1.3.1.4 „Artbegriffe“| beschrieben: Mischlinge aus verschiedenen Arten müssen nicht fruchtbar sein, jedoch das Erbgut beider Elternarten ausprägen. Bei Arten, die nur fossil [= wörtlich „ausgegraben“; gemeint ist: versteinert, als Abdruck oder als Ausguss einer Hohlform erhalten| bekannt sind, kann das definierende Grundtypkriterium nicht angewendet werden. Daher ist es schwieriger, bei Fossilien Grundtypgrenzen zu untersuchen. Dazu muss auf morphologische [= die Gestalt bzw. den äußeren Bau betreffend] Kriterien zurückgegriffen werden: In der Regel kann bei Fossilien nur der Bau der Lebewesen als Informationsquelle herangezogen werden, um eine Zugehörigkeit zum selben Grundtyp zu begründen.
1.2 Grundtypen, die nur fossil bekannt sind
Da man für die Bestimmung von nur fossil bekannten Grundtypen auf morphologische Kriterien zurückgreifen muss, bietet es sich an, die morphologischen Spannweiten heutiger Grundtypen als Vergleichsmaßstab heranzuziehen. Wie im Artikel |0.3.2.1 „Heutige Grundtypen“| geschildert, kann man bei heute lebenden Grundtypen einerseits eine erhebliche gestaltliche Vielfalt beobachten, andererseits aber auch grundtypspezifische morphologische Besonderheiten erkennen. Beides kann als Anhaltspunkt für die Erkennung und Abgrenzung von Grundypen unter Fossilien dienen.
Der Fossilbericht zeigt nun, dass es möglich ist, bis auf das Niveau der Familie hinunter morphologische Abgrenzungen zwischen fossil erhaltenen Gruppen vorzunehmen. Abb. 87 zeigt dies am Beispiel der Paarhufer. Darin ist das fossile Auftreten der verschiedenen Paarhuferfamilien dargestellt. Die heute noch lebenden Familien sind beschriftet. Die taxonomische Stufe der Familie fällt in den meisten der bislang untersuchten Tiergruppen mit den Grundtypen zusammen. Daher könnte die in Abb. 87 wiedergegebene Abgrenzung der Familien als ungefähre Widerspiegelung von Grundtypgrenzen interpretiert werden. Das regelhafte Fehlen von Zwischenformen beinhaltet gleichzeitig eine Schwierigkeit für evolutionäre Deutungen (vgl. Artikel über |1.7.5.1 „Punktualismus“|).
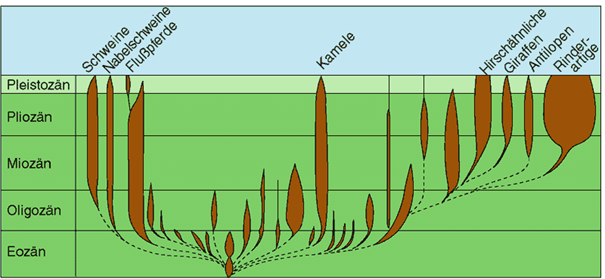
Abb. 87: Fossildokumentation der Paarhufer-Familien. Die Überlieferung fossiler Reste reißt ohne Verbindung durch Übergangsformen zwischen den Familien ab (gestrichelte Linien). Im Tierreich sind Familien häufig identisch mit Grundtypen. Die meisten Familien der Paarhufer sind ausgestorben (diese sind nicht beschriftet). (Nach Romer 1968)
1.3 Fossilien heutiger Grundtypen
Teilweise sind auch Fossilfunde bekannt, die zu den bislang untersuchten heute lebenden Grundtypen gehören, z. B. bei den Greifvögeln und den Entenartigen. Die Zuordnung zu den Grundtypen ist in diesen Fällen gut begründet möglich. So zeigt der Vergleich der Greifvogelfossilien mit den heutigen Formen, dass eine eindeutige Zuordnung zu den fünf Familien auch beim Fossilmaterial möglich ist. Übergangsformen zwischen den einzelnen Gruppen sind nicht bekannt.
Als fossile Form, die einerseits zu den Wat- und Möwenvögeln, andererseits zu den Entenvögeln überleiten soll, wird von manchen Autoren die Gattung Presbyornis sozusagen als „Urente“ diskutiert (Abb. 88). Doch ist diese Stellung umstritten, denn morphologische Merkmale und das Vorkommen eines älteren fossil überlieferten Restes eines deutlich entenartigen Vogels sprechen dafür, dass dieser Vogel viel näher zu den Wat- und Möwenvögeln als zu den Entenvögeln zu stellen ist.

Abb. 88: Rekonstruktion von Presbyornis nach Olson & Feduccia (1980). Das Tier war ca. 40-40 cm hoch.
Detaillierte Studien im Rahmen der Schöpfungslehre liegen in diesem Bereich kaum vor. Wegen Datenmangels sind sie oft auch noch nicht möglich.
1.5 Mosaikformen: Übergangsformen oder eigene Grundtypen?
Im Artikel |1.7.2.1 „Definition von Mosaikform und Übergangsform| wurde der Unterschied zwischen dem beschreibenden Begriff „Mosaikform“ (oder „Zwischenform“) und dem interpretierenden Begriff „Übergangsform“ erläutert. Zweifellos gibt es unter den Fossilien (aber auch bei den heute lebenden Formen) viele Arten, die als Zwischenformen bezeichnet werden können. Das heißt, sie weisen eine Kombination von Merkmalen auf, die sonst für verschiedene Gruppen typisch sind. Dies steht nicht im Gegensatz zur Deutung von Grundtypen als Schöpfungseinheiten. Denn das Grundtypkonzept lässt grundsätzlich verschiedenste Merkmalskombinationen bei verschiedenen Grundtypen zu. Vielmehr beinhaltet das Grundtypkonzept nur, dass nach geeigneten Kriterien Grundtypen voneinander abgrenzbar sind.
Beispielsweise ist der Urvogel Archaeopteryx (Abb. 89) als eigener Grundtyp erkennbar. Von diesem Grundtyp sind 8 Exemplare mit einer gewissen Variabilität bekannt (z. B. ist die größte Form etwa doppelt so groß wie die kleinste). Andererseits ist Archaeopteryx nach bestimmten Merkmalen deutlich gegen andere Vögel und gegen nahestehende Reptil-Gruppen abgrenzbar (Näheres dazu im Artikel |1.7.2.4 „Entstehung der Vögel“|).
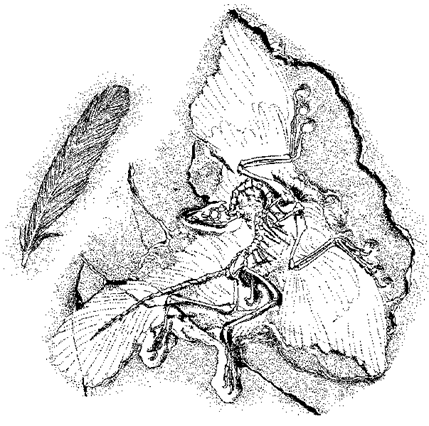
Abb. 89: Urvogel Archaeopteryx (Berliner Exemplar) und Abdruck einer fossilen Feder aus den gleichen Schichten der Solnhofener Plattenkalke.
Auf das Thema „Mosaikformen als Grundtypen“ wird in einem anderen Zusammenhang (s. Artikel |0.3.3.1 Mosaikformen als Grundtypen|) weiter eingegangen.
Literaturhinweis
Junker R & Scherer S (2013) Evolution – ein kritisches Lehrbuch. Gießen, 7. Auflage, Kap. VII.16.3.
Autor: Reinhard Junker, 11.10.2013
© 2013, https://www.genesisnet.info/schoepfung_evolution/i1242.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
0.3.2.4 Genetisch polyvalente Stammformen von Grundtypen
Neben der Frage nach der Abgrenzbarkeit von Grundtypen geht es in der Grundtypenbiologie auch um die Variabilität innerhalb von Grundtypen. Als „Pool“ für die Vielfalt der Grundtypen wird eine genetische Vielseitigkeit (Polyvalenz) der Ursprungsformen angenommen. Dafür gibt es eine Reihe von Hinweisen.
1.0 Inhalt
In diesem Artikel wird erklärt, was unter Polyvalenz von Grundtypen zu verstehen ist und welche Indizien es für eine ursprüngliche Polyvalenz gibt. Vielfalt innerhalb von Grundtypen entstand demnach nicht in erster Linie durch Mutationen, sondern auf der Basis einer vorgegebenen programmierten Variabilität.
1.1 Einleitung
Zum Grundtypkonzept der Schöpfungslehre gehört zum einen die Annahme getrennt erschaffener Grundtypen. Daraus folgt die Erwartung, dass auch heute Grundtypen nach geeigneten Kriterien voneinander abgegrenzt werden können. Dies wird im Artikel |0.3.2.1 Heutige Grundtypen| behandelt. Zum anderen beinhaltet das Grundtypkonzept auch die Annahme, dass die Grundtypen von Anfang an mit einer großen Variationsbreite ausgestattet waren. Sie waren genetisch [= in ihrem Erbgut] vielseitig.
1.2 Polyvalenz
Für diese Variabilität kann es mehrere Quellen gegeben haben. Zum einen kann man annehmen, dass die ursprünglichen Grundtypen in großem Ausmaß mischerbig waren. Die meisten Lebewesen besitzen bekanntlich einen doppelten Chromosomensatz. Auf den Chromosomen befinden sich die Gene. Diese wiederum können unterschiedlich ausgeprägt sein (z. B. Gen für blaue oder braune Augen). Ein Pärchen kann demnach pro Gen maximal vier Zustandsformen (= Allele) besitzen. Insgesamt besitzen die Lebewesen in der Regel mehrere zehntausend Gene, die anfangs alle mischerbig (also bei einem Pärchen mit je Gen bis zu vier Varianten) gewesen sein können. Bei der Fortpflanzung werden die Genvarianten gemischt (= Rekombination) (vgl. Artikel |1.3.2.3 Rekombination und Züchtung|). Da die Merkmale der Lebewesen von vielen Genen gleichzeitig beeinflusst werden, liegt durch die unterschiedlichen Kombinationsmöglichkeiten ein großes Potential an Variationen vor.
Zum anderen können Gene mutieren (vgl. Artikel |1.3.2.1 Mutation|, womit die anfänglich schon vorhandene Variationsbreite weiter ausgedehnt werden kann. Im Artikel über |1.3.2.1 Mutation| wurden Gründe dargelegt, weshalb Mutationen zwar zu Variabilität, nicht aber zu neuen Konstruktionen führen. Möglicherweise waren viele Mutationen vorprogrammiert. Diese und andere Variationsmöglichkeiten werden unter dem Begriff Polyvalenz zusammengefasst.
Die Variabilität von Grundtypen, also ihre mikroevolutive Aufspaltung in Arten und Gattungen, würde nach der Hypothese der polyvalenten Stammformen nicht in erster Linie auf der evolutionären Entstehung neuer Allele, sondern auf einem genetischen Variations-Programm beruhen. Das Ausmaß der Variationsmöglichkeiten wäre vorgegeben und damit begrenzt.
Im Folgenden werden einige Beobachtungen zusammengestellt, die auf eine ursprünglich große, im Laufe der Zeit tendenziell, aber abnehmende Polyvalenz der Grundtypen sprechen.
1.3 Variabilitätsverlust durch Artbildung

Abb. 72: Links: Modifikationspotential von Grundtypen (oben), beispielhaft bezogen auf die Höhenverteilung und Einschränkung des Potentials bei Pflanzen, die nur auf der alpinen Stufe leben können (unten). Rechts: Ein Beispiel liefern Nelkenwurzarten. Die Bach-Nelkenwurz (oben) besitzt Früchte, die (noch?) relativ vielseitig verbreitet werden können: sowohl durch den Wind (aufgrund der Behaarung), durch Tiere (mit Hilfe des Hakens) und durch Verkleben mittels Wasserhaftung. Damit korrespondiert eine weite Verbreitung von Tallagen bis weit über 2000 m Höhe auf Bergwiesen. Die Echte Nelkenwurz (unten) besitzt dagegen einen nur schwach behaarten Hakengriffel, der nur Tierverbreitung (Klettverbreitung) erlaubt. Entsprechend kommt diese Art nur in tieferen Lagen, an Waldrändern, in Gärten usw. vor. Diese Art ist stärker spezialisiert. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001.
Ein Vorgang, der zu einer Verringerung der Polyvalenz führt, ist Artbildung. Artbildung ist empirisch belegt. Im Artikel |1.3.2.4 Artbildung| wird gezeigt, dass fortgesetzte Artbildungsereignisse zu Spezialisierung führen; die so entstandene Tochterart verfügt fast immer über ein geringeres Variationspotential als die Elternart: Der Grad an genetischer Polyvalenz ist reduziert (Abb. 70, 72). Dieser Sachverhalt ist ein Ergebnis empirischer Forschung.

Abb. 70: Hintereinander erfolgende Abspaltungen von Teilpopulationen (Rn) mit dem Genpool rn führen zu Spezialisierungen und Genpoolverarmung, was bildlich durch die kleiner werdenden Ovale angedeutet wird. Quelle: R. Junker & S. Scherer: Evolution – ein kritisches Lehrbuch. Gießen 2001
Wenn man auf dieser Grundlage in die Vergangenheit extrapoliert, wird man den Vorfahren der heute beobachtbaren Arten ein höheres Variationspotential als ihren Nachfahren zuschreiben. Mit anderen Worten: Die der Artbildung zugrunde-
liegenden Vorgänge deuten auf genetisch polyvalente Vorfahren hin. Je weiter diese Vorfahren zeitlich zurückliegen, desto mehr Artbildungsereignisse wird man annehmen und desto höher muss wohl die genetische Polyvalenz ihres Erbmaterials gewesen sein. Aus dieser Argumentationslinie entwickelt sich nahezu zwangsläufig die Vorstellung eines komplexen statt eines primitiven Vorfahren (Abb. 90).

Abb. 90: Artbildung in der Evolutions- und Schöpfungsvorstellung. Die genetische Polyvalenz der Populationen ist durch die Breite der Stammbaumäste angedeutet. Der Evolutionstheoretiker nimmt an, dass der Verlust genetischer Polyvalenz bei der Artbildung durch im Einzelnen unbekannte Prozesse wieder ausgeglichen wird. Im Grundtypkonzept wird dagegen von anfänglich polyvalenten Grundtypen ausgegangen.
Es gibt nur einen Weg, diesen für die Makro-Evolutionslehre kaum akzeptablen generellen Schluss zu umgehen: Durch andere Evolutionsprozesse (etwa Mutation) muss die genetische Polyvalenz nach der Artbildung immer wieder neu erzeugt worden sein. Allerdings ist empirisch nicht zu beobachten, wie durch Evolutionsprozesse die verlorengegangenen Allele durch neuartige ersetzt werden können, und theoretische Modelle dazu liegen auch nicht vor (vgl. |1.3.2.1 Mutation| und |1.3.4.2 Beispiel Bakterienmotor|). Im Rahmen der Grundtypenbiologie der Schöpfungslehre ist ein genetisch polyvalenter Vorfahre zu erwarten.
1.4 Verteilung von Merkmalen innerhalb von Grundtypen
Die Verteilung von Merkmalen auf verschiedene Arten ist die Grundlage für die Erstellung von Hypothesen über Verwandtschaft. Merkmalsverteilungen innerhalb von Grundtypen können im Sinne genetisch polyvalenter Vorfahren der heutigen Grundtypen gedeutet werden. Einige Beispiele sollen dies verdeutlichen.
Aberrante Arten. Unter den Entenvögeln findet man einige Arten, die den Taxonomen schon immer Probleme bereitet haben; man fasst sie auch unter dem Begriff „aberrante Arten“ zusammen. Sie zeichnen sich dadurch aus, dass sie in sich Merkmale von ganz verschiedenen Entengattungen vereinigen. Deshalb können sie nicht ohne weiteres anderen Entenvögeln zugeordnet werden, und man stellt sie in eine eigene Tribus [= Klassifikationsstufe zwischen Gattung und Familie]. Die Hühnergans (Abb. 91) ist ein solcher aberranter Typ. Sie vereinigt in sich Merkmale der Halbgänse, der Gänse und der Enten. Die Dampfschiffenten gehören nach ihren Federproteinen in die Nähe der Halbgänse, nach den Skelettmerkmalen zu den Enten und nach der Zusammensetzung der Bürzeldrüsenwachse in die Verwandtschaft der Eiderente. Solche Merkmalswidersprüche innerhalb von Grundtypen sind allgemein häufig und werden im Evolutionsmodell normalerweise durch Konvergenz [= mehrfach unabhängige Entstehung baugleicher Merkmale] erklärt (vgl. dazu Artikel |1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie|). Das setzt jedoch die Existenz der Selektionsfaktoren und Mutationsmechanismen voraus, welche in verschiedenen Arten unabhängig zum Auftreten des gleichen Merkmals führten. Diese sind in der Regel unbekannt. Im Grundtypmodell der Schöpfungslehre vermutet man, dass in der Stammform der Enten die Kapazität zur Ausprägung der in Frage stehenden Merkmale vorhanden war. Die Merkmale haben sich auf dieser Basis in mehr oder weniger zufälligen Kombinationen auf die Nachfahren verteilt. Ähnliche Befunde gibt es auch hinsichtlich der Gefiederzeichnungen oder der Schnabelformen.

Abb. 91: Die Hühnergans (Cereopsis) kann nicht eindeutig in eine der Untergruppen der Entenartigen eingeordnet werden, weil sie in sich Merkmale von ganz verschiedenen Gruppierungen der Entenvögel vereinigt.
Parallelvariation bei Meisen. Bei Meisen wurde beobachtet, dass sogenannte Parallelvariationen auftauchen. Darunter versteht man das parallele Auftreten sehr ähnlicher Phänotypen [= äußere Erscheinungsform eines Organismus] in Rassen unterschiedlicher Arten. Die Parallelvariationen konnten nicht als Reaktion auf Selektionsdrücke gedeutet werden. S. Eck schreibt dazu (1988, S. 62; vgl. Abb. 92): „Der Zusammenhang von räumlicher Anordnung und Merkmalsanordnung in Proportionen und Pigmentierung der Nest- und Alterskleider bei den Kohlmeisen (Parus major) von major im engeren Sinne bis bokharensis wiederholt sich in räumlich anderer Reihung spiegelbildlich so genau bei den fernerstehenden Blau- und Lasurmeisen (Parus caeruleus und Parus cyanus), dass es so abwegig nicht ist, von Programmierung zu sprechen.“ Vergleichbare Phänomene wurden auch bei Getreidearten und den verwandten Wildgräsern und anderen Nutzpflanzen in zahlreichen Fällen beobachtet. Der russische Genetiker Vavilov formulierte bereits 1925 daraus sogar ein „Gesetz der Parallelvariationen“.

Abb. 92: Europäische Blaumeise (Parus caeruleus) und Kohlmeise (Parus major). Die unabhängig entstandenen Unterarten der beiden Arten entsprechen sich im Äußeren in derart vielen Details (Parallelvariation), dass der Schluss auf eine genetisch programmierte Variabilität nahe liegt.
Regel der rekurrenten Variation. Im Artikel über |1.3.2.1 Mutation| wird die „Regel der rekurrenten Variation“ erläutert. Danach bringen künstlich ausgelöste Mutationen zum einen auf Dauer kaum noch neue Varianten, zum anderen gleichen Labormutanten den im Freiland auftretenden Mutationen weitgehend. Auch dieser Befund weist darauf hin, dass es zwar eine große, aber letztlich begrenzte Zahl von Merkmalsausprägungen gibt, und passt gut zum Konzept der polyvalenten Stammformen.
Zusammenfassende Deutung. Die vorstehenden Beispiele für Merkmalsverteilungen innerhalb von Grundtypen sind sehr gut mit der Vorstellung genetisch polyvalenter Stammformen verträglich. Abb. 93 kann als Ansatz für eine Arbeitshypothese dienen, um die molekulargenetischen Grundlagen der besprochenen Strukturen zu erforschen (vgl. auch Abb. 94). Der Vorfahr verfügte über eine Reihe von Merkmalskomplexen (Aa,b,c,d,e,f…; Ba,b,c,d,e,f…; Ca,b,c,d,e,f… usw.), welche die Zustände a,b,c,… annehmen können. Diese Merkmalskomplexe lagen entweder in der Ursprungspopulation polymorph [= in verschiedenen Ausprägungen] vor oder sie konnten durch Regulationsmutationen angeschaltet und teilweise durch Mutationen erzeugt werden. Während der Artbildung verteilten sich die Ausprägungszustände auf die entstandenen Arten, deren Variationsfähigkeit mit zunehmenden Artbildungsereignissen immer mehr abnahm.
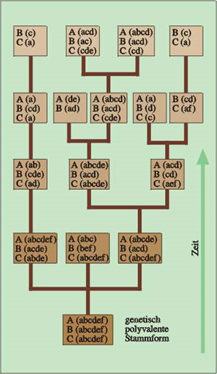
Abb. 93: Eine genetisch polyvalente Stammform birgt in sich viele Möglichkeiten der Ausprägung von Merkmalen in den nachfolgenden Generationen. A, B, C stehen für Merkmalskomplexe; a, b, c,… für deren unterschiedliche Ausprägungen.

Abb. 94: Deutung von Merkmalsverteilungen innerhalb von Grundtypen. Die kleinen Ovale symbolisieren Arten eines Grundtyps mit den Merkmalen a-f. A Von einer polyvalenten Stammform ausgehend (mit dem kompletten Merkmalsbestand) entstehen einzelne Arten mit eingeschränkter Vielfalt, wobei die Merkmale unsystematisch verteilt sind. Bei Zugrundelegung einer primitiven Stammform (B) kann diese Merkmalsverteilung nur unter der Annahme mehrerer Konvergenzen (unabhängige Entstehung baugleicher Merkmale) oder Reversionen (Rückentwicklungen) erklärt werden.
1.5 Ähnlichkeiten von Mischlingen mit dritten Arten
Kreuzungen zwischen verschiedenen Arten innerhalb eines Grundtyps sind häufig. Manchmal treten dabei Mischlinge auf, deren Phänotypen [= äußere Erscheinungsform eines Organismus] weniger ihren Eltern und mehr dritten Arten aus dem Grundtyp ähneln. Die in Abb. 95 oben abgebildete Reiherente kann mit der darunter abgebildeten Tafelente kreuzen und den an dritter Stelle gezeichneten Mischling hervorbringen. Dieser wird in Parkanlagen unweigerlich mit der nur in Amerika lebenden Veilchenente verwechselt (ganz unten abgebildet). Mehrere solcher Fälle sind bekannt, wo die Kreuzung zweier Arten des Grundtyps der Entenartigen Ähnlichkeiten mit einer dritten Art hervorbringt. Auch bei den Finkenvögeln sind derartige „Phänokopien“ bekannt geworden.

Abb. 95: Ähnlichkeit eines Mischlings mit einer dritten Art. Von oben nach unten: Reiherente (Aythya fuligula), Tafelente (Aythya ferina). Mischling aus beiden, Veilchenente (Aythya affinis). Zeichnung: Thomas HILSBERG.
Ein besonders eindrucksvolles Beispiel sind Mischlinge von Pudel und Wolf (Abb. 96), bei welcher in der zweiten Generation erstaunliche verborgene Variationspotentiale der Elternrassen offengelegt werden. Offenbar sind im Erbgut der beteiligten Arten Variationspotentiale vorhanden, welche durch eine artübergreifende Kreuzung aktiviert werden.

Abb. 96: „PuWos“ der zweiten Generation: Die Nachkommen von Mischlingen aus Pudel und Wolf. Sie zeigen eine reiche Vielfalt von Merkmalskombinationen.
1.6 Explosive Artbildung
In den letzten Jahren wurden vermehrt Beispiele für schnelle Artbildungsprozesse bekannt. Sie liefern Hinweise auf genetisch polyvalente Stammformen. Als Beispiel soll die sehr gut untersuchte Bildung von Artenschwärmen bei Buntbarschen dienen.
Eine Vielfalt von Buntbarscharten (vgl. Abb. 97) kommt in den großen ostafrikanischen Seen vor. Im Viktoriasee leben über 300 Buntbarscharten in etwa acht Gattungen. Mit molekularbiologischen Methoden konnte gezeigt werden, dass alle Arten des Viktoriasees von einer einzigen Stammgruppe abstammen. Das bedeutet, dass sich sämtliche Arten des Sees in diesem See entwickelt haben müssen; sie können daher nicht älter sein als der See. Dennoch sind sie sehr verschieden. Geologische Ergebnisse zeigen, dass der Viktoriasee vor rund 12.000 Jahren mit großer Wahrscheinlichkeit komplett austrocknete. Damit ist das Maximalalter der Buntbarsche in diesem See festgelegt. Folglich sind die Buntbarsche des Viktoriasees ein Beispiel für ein bemerkenswertes Tempo morphologischer [= die äußere Gestalt betreffend] Aufspaltungen.

Abb. 97: Beispiele für die Vielgestaltigkeit der Buntbarsche in ostafrikanischen Seen. Von oben: Pseudotropheus saulosi, Lamprologus spec. und Chalinochromis ndobhoi.
Die Mbuna-Fauna des Malawisees aus rund 200 Arten ist von besonderem Interesse. Sie umfasst nicht nur die farbenprächtigsten Arten, sondern fast jede felsige Erhebung des Malawisees besitzt ihre eigenen Fischarten, die nur auf den jeweiligen Inseln vorkommen. Die einzelnen Arten sind oft räumlich von denen anderer Inseln oder Uferbereiche isoliert. Viele der heutigen Inseln waren vor ca. 200 Jahren von trockenem Land umgeben, die Lebensräume der Fische existierten zeitweise dort also gar nicht. Daher muss die Entstehung dieser einzigartigen Faunen in den letzten rund 200 Jahren stattgefunden haben. Diese Aussage wird gestützt von gut belegten topographischen und historischen Veränderungen, Untersuchungen der Unterschiede des Erbguts und der Verbreitung der betreffenden Fischarten.
Arten mit vielen potenziellen Entfaltungsmöglichkeiten werden als Generalisten bezeichnet. Früher wurde die Vermutung geäußert, dass die Besiedlung des Viktoria- und Malawisees durch solche Generalisten erfolgt sei. Die heute vorliegenden Daten stützen diese Hypothese, denn eine derart schnelle Artbildung lässt für die Erzeugung neuer Gene durch Mutation keinen genügenden zeitlichen Spielraum. Explosive Artbildung ist nur noch mit genetisch polyvalenten Stammformen zu erklären. Das heißt, dass die Stammformen über genetisches Material verfügten, welches unterschiedliche Merkmalsausprägungen zuließ.
1.7 Literatur
Eck S (1988) Ist Otto Kleinschmidts Formenkreis-Konzept wieder aktuell? Proc. Int. 100. DO-G Meeting. Current Topics in Avian Biology. Bonn, S. 61-66.
Fehrer J (1997) Explosive Artbildung bei Buntbarschen der ostafrikanischen Seen. Studium Integrale Journal 4, 51-55.
Junker R (1993) Prozesse der Artbildung. In: Scherer S (Hg) Typen des Lebens. Berlin, S. 31-45.
Junker R & Scherer S (2001) Evolution – ein kritisches Lehrbuch. Gießen, 5. Auflage, Kap. VII.17.3
Lönnig W-E (1995) Mutationen: Das Gesetz der rekurrenten Variation. In: Mey J, Schmidt R & Zibulla S (Hg) Streitfall Evolution. Kontroverse Beiträge zum Neodarwinismus. Stuttgart, S. 149-166.
Scherer S (1998) Abnehmender Sexappeal von männlichen Buntbarschen durch Umweltverschmutzung des Viktoriasees. Studium Integrale Journal 5, 85-86.
Autor: Reinhard Junker, 12.01.2004
© 2004, https://www.genesisnet.info/schoepfung_evolution/i1244.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
0.3.2.6 Kritik an der Grundtypenbiologie (Interessierte)
An den Ansätzen der Grundtypenbiologie wurde verschiedentlich Kritik geäußert. Dabei geht es vor allem um wissenschaftstheoretische Fragen, aber auch um konkrete Sachkritik und um Behauptungen über inkonsequente Deutungsweisen in der Grundtypenbiologie. Durch die Entgegnungen auf diese Kritikpunkte wird ein vertieftes Verständnis des Grundtypen-Ansatzes ermöglicht: dieses Konzept ist prüfbar und fördert Forschung.
1.0 Inhalt
In diesem Artikel wird Kritik vorgestellt, die an der Grundtypenbiologie geübt wurde. Es wird gezeigt, dass entgegen der Ansicht der Kritiker die Ansätze der Grundtypenbiologie prüfbar sind und Forschung anregen. Vermeintliche Widersprüche innerhalb der Grundtypenbiologie werden aufgelöst.
1.1 Einleitung
Der Ansatz der Grundtypenbiologie wurde bisher von der Fachwelt nur wenig beachtet. Hier und da wurde in den letzten Jahren aber Kritik publiziert. Manchmal zeigte sich, dass der Kritiker mit den Inhalten der Grundtypenbiologie praktisch überhaupt nicht vertraut war (ein Beispiel schildert [#NEWS 23 Grundtypenmodell der Schöpfungslehre widerlegt?|). Es wurde aber auch seriösere Kritik geäußert, zum Beispiel von Korthof (2004). Auf diese wird in diesem Artikel eingegangen. Eine wesentlich detaillierte Auseinandersetzung mit Kritikpunkten wird im Expertenteil geführt.
1.2 Ist das Grundtypmodell testbar?
Zur Beantwortung muss auf die wichtige Unterscheidung zwischen „Schöpfungsparadigma“ und Hypothesen bzw. Theorien der „Grundtypenbiologie“ hingewiesen werden. Dieser Unterschied wird im Expertenteil des Artikels |1.1.3.2.2 Methodik der historischen Forschung| näher erläutert. Unter Schöpfungsparadigma wird ein von Naturgesetzen nicht ableitbares ins-Dasein-Kommen (der ursprünglichen Lebewesen) durch Befehle Gottes („durch das Wort“) verstanden. Dieser paradigmatische Rahmen ist nicht direkt prüfbar, sondern kann nur auf Plausibilität getestet werden. In diesem Rahmen können aber konkrete prüfbare Hypothesen und Theorien aufgestellt werden. Sie werden als „Grundtypenbiologie“ zusammengefasst (|0.3.2.1 Heutige Grundtypen|, |0.3.2.4 Genetisch polyvalente Stammformen von Grundtypen|) bzw. es wird vom „Grundtypmodell“ gesprochen. Eine ähnliche Unterscheidung zwischen paradigmatischem Rahmen und konkreten, prüfbaren Theorien muss auch in der Evolutionsbiologie vorgenommen werden; auch dort gilt, dass nicht das Paradigma, sondern nur die untergeordneten Hypothesen und Theorien prüfbar sind (|1.1.3.4 Evolutionsparadigma und Naturwissenschaft|).
Das Grundtypmodell erlaubt folgende Testmöglichkeiten:
Grundtypen müssen nach dem definierenden Kreuzungskriterium von den nächst ähnlichen Grundtypen abgrenzbar sein. (Die Grundtypdefinition wird im Artikel |0.3.2.1 Heutige Grundtypen| vorgestellt; die Kenntnis dieses Artikels wird hier vorausgesetzt.) Das Kriterium der indirekten Kreuzbarkeit (Kreuzungsverbindung mittels einer dritten Art) ist hier wichtig, um nachträglich aufgetretene Lücken und primäre (von Anfang an bestehende, schöpfungsgemäße) Grundtypgrenzen besser auseinanderhalten zu können. Erst ein Gesamtbild von den Kreuzungsmöglichkeiten kann zeigen, ob sich Kreuzungen innerhalb mutmaßlicher Grundtypgrenzen häufen und ob es scharfe Grenzen gegenüber anderen mutmaßlichen Grundtypen gibt (vgl. Abb. 82). Die Bestimmung von Grundtypgrenzen ist hypothetisch; weitere Überprüfungen können diese Grenzen verschieben.
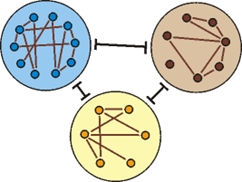
Abb. 82: Bei den bisher untersuchten Tier- und Pflanzengruppen konnten deutliche Grundtypgrenzen festgestellt werden. Innerhalb der Grundtypen (große Kreise) sind die verschiedenen Biospezies (kleine Kreise) jedoch eng verbunden.
Weiter kann geprüft werden, ob es Merkmale im Bau, im Verhalten, in der Biochemie, in der Genetik usw. gibt, die eine Abgrenzbarkeit von Grundtypen nahelegen. Auch Fossilfunde können herangezogen werden. Dies führt in ein weites Forschungsfeld hinein. Die Grundtypenbiologie regt also Forschung an.
Es sei darauf hingewiesen, dass deutliche Lücken zwischen benachbarten Grundtypen die Evolutionstheorie nicht widerlegen; sie schwächen aber ihre Plausibilität, wenn sie systematisch auftreten, und sie bestätigen eine Vorhersage des Grundtypmodells.
Eine weitere Testmöglichkeit ist das Aufdecken von Befunden, die für eine große Polyvalenz der Stammformen sprechen (siehe dazu |0.3.2.4 Genetisch polyvalente Stammformen von Grundtypen|).
Auf den Einwand, diese Prüfmöglichkeiten seien nicht spezifisch für das Grundtypmodell wird am Ende des Expertenteils eingegange (Abschnitt „Ist der Test des Grundtypmodells ein Test auf Schöpfung?“ in |0.3.2.6.2 Kritik an der Grundtypenbiologie|).
Bisher liegen knapp 20 Grundtypstudien vor, die zeigen, dass sich das Konzept bewährt hat. Zweifellos ist das eine sehr geringe Zahl und weitere Grundtypstudien wären wünschenswert.
Grundtypgrenzen fallen nicht generell mit einem bestimmten taxonomischen Niveau zusammen. Zwar ist der Grundtyp bei Wirbeltieren häufig mit der Familie identisch, doch muss dies nicht immer so sein, sondern muss von Fall zu Fall geprüft werden. Daher ist es falsch, den Menschen mit Menschenaffen in denselben Grundtyp zu stellen, nur weil beide von vielen Taxonomen in dieselbe Familie gestellt werden.
1.3 Wie könnte die Existenz schöpfungsgemäß getrennter Grundtypen falsifiziert werden?
Das Grundtypkonzept, so wie es derzeit formuliert ist (|0.3.2.1 Heutige Grundtypen|), könnte wie folgt falsifiziert werden:
- Würden Kreuzungen zwischen Menschen und Tieren zweifelsfrei gelingen, wäre das Grundtypkonzept in der vorgelegten Form nicht haltbar.
- Würden sich keine Gruppen von Lebewesen bei Anwendung des Grundtypkriteriums abgrenzen lassen, wäre das Grundtypkonzept falsifiziert.
Die Existenz anfänglich getrennter Grundttypen wäre damit zwar nicht widerlegt, wohl aber die derzeitige konkrete Ausgestaltung des Grundtypkonzepts. (Hier muss der Unterschied zwischen allgemeinem „Schöpfungsparadigma“ und konkretem Grundtyp beachtet werden, s. o.)
Daten, die im Rahmen des Grundtypkonzepts plausibel gedeutet werden können, stützen dagegen diesen Ansatz: Hier kann vor allem an die zahlreichen Belege für die Existenz polyvalenter Stammformen eines Formenkreises verwiesen werden (vgl. dazu |0.3.2.4 Genetisch polyvalente Stammformen von Grundtypen|). Außerdem ist der Befund, dass fortgesetzte Artaufspaltungen durch zunehmende Spezialisierung in evolutionäre Sackgassen führen, im Rahmen des Grundtypkonzepts plausibel deutbar (|1.3.2.4 Artbildung|). Entsprechend besteht Forschungsinteresse, mehr über Variationsmechanismen herauszufinden; auch hier gilt also: die Grundtypenbiologie regt Forschung an.
1.4 Gemeinsamkeiten verschiedener Grundtypen
Verschiedene Grundtypen weisen eine mehr oder weniger große Zahl von Gemeinsamkeiten (Homologien) auf. Diese Gemeinsamkeiten sollen auf gemeinsame Vorfahren hinweisen, aus denen verschiedene Grundtypen entstanden sind. Dies stehe nach Ansicht mancher Kritiker dem Grundtypen-Ansatz entgegen. Doch hier wird mit Ähnlichkeiten argumentiert. Ähnlichkeiten an sich sprechen aber nicht für gemeinsame Abstammung, wie im Artikel
(|1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie|) gezeigt wird.
Arten desselben Grundtyps gehen nach dem Grundtypkonzept auf eine gemeinsame (geschaffene) Stammart zurück, Arten verschiedener Grundtypen dagegen nicht. Dennoch weisen verschiedene Grundtypen vergleichbare Ähnlichkeiten auf wie Arten innerhalb desselben Grundtyps. Es gibt also keine einheitliche Erklärung für Ähnlichkeit; dies erscheint Kritikern inkonsequent. Das Fehlen einer einheitlichen Erklärung für Ähnlichkeit ist jedoch kein Gegenargument gegen das Grundtypkonzept. Auch im Rahmen des Evolutionsparadigmas gibt es keine einheitliche Erklärung für Ähnlichkeiten, auch nicht für solche, die nach vergleichend-biologischen Kriterien als homolog eingestuft werden
(|1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie|). Viele nach biologischen Kriterien als homolog einzustufende Ähnlichkeiten können aufgrund widersprüchlicher Merkmalsverteilungen nicht auf gemeinsame Vorfahren zurückgeführt werden, sondern werden als Konvergenzen interpretiert. Das gilt auch für viele Komplexmerkmale (Beispiele in Junker 2002, 85ff.). Im Rahmen der Grundtypenbiologie wird der Unterschied zwischen abstammungsbedingten und nicht-abstammungsbedingten Ähnlichkeiten an den Grundtypgrenzen festgemacht, die anhand eines von den Merkmalen unabhängigen Kriteriums – dem Kreuzungskriterium – erkannt werden.
1.5 Grundtyp-Diversifikation und Makroevolution
Die Diversifikation [= Verschiedenwerden] innerhalb von Grundtypen erfolgt durch mikroevolutive Prozesse. Diese würden aber aber nach Auffassung von Kritikern bei weitem nicht ausreichen, um die Vielfalt innerhalb von Grundtypen in kurzer Zeit zu erzeugen. Diese Kritik übersieht, dass das Grundtypmodell damit rechnet, dass die Grundtypen zu Beginn ihrer Existenz polyvalent waren. Die Hauptquelle der Variabilität steckt demnach bereits in den (geschaffenen) Stammformen (vgl. |0.3.2.4 Genetisch polyvalente Stammformen von Grundtypen|).
Im Rahmen der Grundtypenbiologie besteht großes Interesse, Polyvalenz und Variationsmechanismen nachzuweisen. Dies macht einmal mehr deutlich, dass das Schöpfungsparadigma Wissenschaft nicht verhindert – im Gegenteil: Je mehr über die Variationsmechanismen bekannt ist, desto aussagekräftiger kann die Plausibilität des Grundtypmodells eingeschätzt werden.
Polyvalenz schließt auch das Mutationspotential ein. Aber alleine schon die Kombinationen verschiedener Allele [= Zustandsformen eines Gens] könnten ein erhebliches Variationspotential bergen. Abb. 96 gibt hierzu modellhaft (!) einen Eindruck. Es handelt sich dabei um die vielfaltige Nachkommenschaft von äußerlich einheitlichen F1[= erste Tochtergeneration]-Mischlingen aus Pudel und Wolf (Abb. 213 zeigt die einheitliche Elterngeneration der in Abb. 96 gezeigten Tiere der F2-Generation). In der F2-Generation tritt die genetische Vielfalt durch unterschiedliche Kombinationen zutage.

Abb. 213: Äußerlich einheitliche F1-Mischlinge aus Pudel und Wolf, die genetisch
vielfach mischerbig sind.

Abb. 96: „PuWos“ der zweiten Generation: Die Nachkommen von Mischlingen aus Pudel und Wolf. Sie zeigen eine reiche Vielfalt von Merkmalskombinationen.
Hier wird beispielhaft sichtbar, dass eine morphologisch [= gestaltlich] einheitliche Elterngeneration (hier: die F1-Generation der PuWos) in kürzester Zeit (in wenigen Generationen) eine erhebliche Vielfalt zeigen kann, wenn sie anfangs sehr polymorph [= genetisch vielseitig] war. Die PuWos der F1-Generation können als Modell (!) für einen ursprünglichem Grundtyp betrachtet werden. Im Grundtypmodell wird davon ausgegangen, dass die ursprünglichen Grundtypen noch deutlich mehr Variationpotential besaßen als die F1-Generation der PuWos. Ein großes anfängliches Variationspotential bietet auch die Basis für hohe Artbildungsraten, die im Rahmen des Grundtypmodells zu erwarten sind.
Die Grundtypdiversifikation erfordert keine Makroevolution. Unter der Voraussetzung polyvalenter Stammformen ist nur Variation erforderlich. Evolution oberhalb des Biospezies-Level ist noch keine „Makroevolution“. Der Unterschied zwischen Mikro- und Makroevolution wird im Grundtypmodell nicht an Grenzen zwischen Gattungen, Familien oder anderen Gruppierungen festgemacht, sondern an Qualitäten (Entstehung neuer Konstruktionen bzw. neuer Basisfunktionszustände; vgl. |1.3.1.3 Mikro- und Makroevolution|). Geht man von poylvalenten Stammformen aus, sind neue Konstruktionen bei der Grundtypaufspaltung und mithin Makroevolution nicht erforderlich.
1.6 Die Grundtypen tauchen im Fossilbericht nicht gleichzeitig auf
Dieser Einwand trifft im Wesentlichen zu. Das zeitlich gestaffelte fossile Auftreten verschiedener Taxa [= Einheiten der Klassifikation] mit verschiedenen Grundtypen im Fossilbericht ist in der Tat ein schwerwiegendes Problem für die biblische Schöpfungslehre, die dem biblischen Schöpfungsbericht folgend von einer gleichzeitigen Erschaffung aller Grundtypen ausgeht. Stephan (2002) diskutiert Lösungsansätze, die aber noch viele Fragen offen lassen. Allerdings ist innerhalb von einzelnen geologischen Systemen ein nahezu gleichzeitiger erster fossiler Nachweis von Grundtypen regelmäßig anzutreffen, besonders markant bei der kambrischen Explosion, aber auch bei vielen Tier- und Pflanzengruppen, die in jüngeren Systemen fossil überliefert sind (siehe |1.7.2.2 Kambrische Explosion|).
1.7 Die Entstehung der Grundtypen ist ein Geheimnis
Manche Kritiker behaupten, die Grundtypenbiologie ersetze eine bereits vorliegende gute Erklärung für die Entstehung der Grundtypen durch ein Mysterium. Es gibt seit Darwin jedoch nur die Behauptung einer Erklärung, aber der Beweis fehlt. Darüber kann man kontrovers diskutieren (was im Evolutionsteil von Genesisnet geschieht). Die Kritik am Grundtypmodell basiert hier demnach auf einer falschen Voraussetzung, nämlich dass Darwin bereits eine Erklärung für die Entstehung der Lebewesen gehabt habe. Diese Erklärung hatte er nicht und niemand hat sie bis heute.
Literatur
Junker R (2002) Ähnlichkeiten, Rudimente, Atavismen. Holzgerlingen.
Korthof G (2004) Common Descent. It’s All or Nothing. In: Young M & Edis T (eds) Why Intelligent Design Fails. A Scientific Critique of the New Creationism. New Brunswick, pp. 32-47.
Stephan M (2002) Der Mensch und die geologische Zeittafel. Holzgerlingen.
0.3.2.6 Kritik an der Grundtypenbiologie (Experten)
In diesem Artikel wird Kritik vorgestellt, die an der Grundtypenbiologie geübt wurde. Es wird gezeigt, dass entgegen der Ansicht der Kritiker die Ansätze der Grundtypenbiologie prüfbar sind und Forschung anregen. Vermeintliche Widersprüche innerhalb der Grundtypenbiologie werden aufgelöst.
Siehe auch den PDF-Artikel: https://www.wort-und-wissen.org/wp-content/uploads/Die_Grundtypenbiologie_in_der_Kritik.pdf
2.1 Einleitung
Der Ansatz der Grundtypenbiologie wurde bisher von der Fachwelt nur wenig beachtet. Über die Gründe soll hier nicht spekuliert werden. Hier und da wurden in den letzten Jahren aber vermehrt Kritikpunkte geäußert. Manchmal zeigte diese Kritik, dass der Kritiker mit den Inhalten der Grundtypenbiologie praktisch überhaupt nicht vertraut war (ein Beispiel schildert [#NEWS 23 Grundtypenmodell der Schöpfungslehre widerlegt?|). Es wurde aber auch seriösere Kritik geäußert. Auf diese wird in diesem Artikel eingegangen.
Vorneweg soll festgestellt werden, dass die Existenz dieser Kritik zeigt, dass die Grundtypenbiologie zu Recht einen Platz in der Wissenschaft hat. Die Aussagen und Begründungen der Grundtypenbiologie können rational diskutiert werden, auch wenn man ihre dogmatische Motivation nicht teilt oder gar nicht kennt. Außerdem macht die Grundtypenbiologie testbare bzw. falsifizierbare [= widerlegbare] Aussagen. Mit diesem Thema soll die Liste von Einwänden begonnen werden.
2.2 Testmöglichkeiten des Grundtypmodells
Zunächst muss auf die wichtige Unterscheidung zwischen „Schöpfungsparadigma“ und Hypothesen bzw. Theorien der „Grundtypenbiologie“ hingewiesen werden. Dieser Unterschied wird im Expertenteil des Artikels |1.1.3.2 Methodik der historischen Forschung| näher erläutert. Unter Schöpfungsparadigma wird ein von Naturgesetzen nicht ableitbares ins-Dasein-Kommen (der ursprünglichen Lebewesen) durch Befehle Gottes („durch das Wort“) verstanden. Dieser paradigmatische Rahmen ist nicht direkt prüfbar, sondern kann nur auf Plausibilität getestet werden. In diesem Rahmen können aber konkrete prüfbare Hypothesen und Theorien aufgestellt werden. Sie werden als „Grundtypenbiologie“ zusammengefasst (|0.3.2.1.1 Heutige Grundtypen|, |0.3.2.4.1 Genetisch polyvalente Stammformen von Grundtypen|) bzw. es wird vom „Grundtypmodell“ gesprochen. Eine ähnliche Unterscheidung zwischen paradigmatischem Rahmen und konkreten, prüfbaren Theorien gilt auch für die Evolutionsbiologie; auch dort gilt, dass nicht das Paradigma, sondern nur die untergeordneten Hypothesen und Theorien prüfbar sind (|1.1.3.4 Evolutionsparadigma und Naturwissenschaft|).
Das Grundtypmodell erlaubt folgende Testmöglichkeiten:
Grundtypen müssen nach dem definierenden Kreuzungskriterium von Nachbargrundtypen abgrenzbar sein. (Die Grundtypdefinition wird im Artikel |0.3.2.1.1 Heutige Grundtypen| vorgestellt; die Kenntnis dieses Artikels wird hier vorausgesetzt.) Unter Nachbargrundtypen sind solche zu verstehen, die morphologisch-anatomisch oder nach anderen Kriterien dem untersuchten Grundtyp am nächsten stehen. Das Kriterium der indirekten Kreuzbarkeit (Kreuzungsverbindung mittels einer dritten Art) ist dabei wichtig, um sekundär aufgetretene Lücken und primäre Grundtypgrenzen auseinanderhalten zu können. Mit Hilfe des definierenden Kreuzungskriteriums alleine können nur Zugehörigkeiten zum selben Grundtyp sicher festgestellt werden, nicht aber die Nicht-Zugehörigkeit, da das Ausbleiben von Kreuzbarkeit durch eine einzige Mutation von zuvor noch kreuzbaren Individuen verursacht sein kann. Daher ist es erforderlich, ein Gesamtbild von den Kreuzungsmöglichkeiten zu bekommen. Aus diesem Grund können Grundtypgrenzen immer nur auf Vorbehalt bestimmt werden. Die Bestimmung von Grundtypgrenzen ist hypothetisch; weitere Überprüfungen können diese Grenzen verschieben. Im Idealfall sollte sich jedoch folgendes zeigen:
- Grundtypen sollten intern vielfältige Kreuzungs-Beziehungen zwischen den zugehörigen Arten aufweisen. Wichtig ist also der Gesamteindruck: Gibt es auffällige Häufungen von Kreuzungen, denen markante Grenzen gegenüberstehen? (vgl. Abb. 82).
- Es sollten Befunde aufgedeckt werden, die für eine große Polyvalenz der Stammformen sprechen, sowohl in genetischer als auch entwicklungsbiologischer und evtl. noch anderer Hinsicht (siehe dazu |0.3.2.4.1 Genetisch polyvalente Stammformen von Grundtypen|).
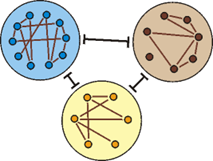
Abb. 82: Bei den bisher untersuchten Tier- und Pflanzengruppen konnten deutliche Grundtypgrenzen festgestellt werden. Innerhalb der Grundtypen (große Kreise) sind die verschiedenen Biospezies (kleine Kreise) jedoch eng verbunden.
Neben dem definierenden Kreuzungskriterium können auch noch andere Kriterien zur Abgrenzung von Grundtypen zum Einsatz kommen, z. B. morphologische, verhaltensbiologische, biochemische oder molekularbiologische. In der Paläontologie muss notgedrungen auf solche weicheren Kriterien zurückgegriffen werden (was aber kein Spezifikum für die Grundtypenbiologie ist). Dies führt in ein weites Forschungsfeld hinein, denn um Grundtypgrenzen plausibel machen zu können, sind möglichst viele Kenntnisse über vermutete Grundtypen erforderlich. Interessant wäre es beispielsweise, zu prüfen, ob verschiedene Grundtypen sich durch den Besitz oder das Fehlen eines irreduzibel komplexen Merkmals gegen nächstähnliche Grundtypen abgrenzen lassen. Damit könnte auch eine interessante Verbindung zum Design-Argument hergestellt werden (|0.4.1.1 Einführung in „Intelligent-Design“|).
Auch im molekularen Bereich könnte es Kriterien zur Aufdeckung oder Bestätigung von Grundtypgrenzen geben. Es ist zu prüfen, ob das weitere Anwachsen des Datenmaterials auf Grundtypebene zu mehr Klärung der Beziehungen beiträgt; im Fall von Vergleichen außerhalb der Grundtypen könnte der umgekehrte Effekt eintreten: je mehr Daten, desto mehr Widersprüche (Fehrer 2000, 31). Mittlerweile liegen jedoch Beispiele vor, dass dieser Effekt der zunehmenden Widersprüche bei Zuwachs an Datenmaterial auch auf Grundtypebene vorkommt (Fehrer, persönl. Mitt.).
Evolutionstheoretisch würde man zwischen enger verwandten Formen primär eher ein Formenkontinuum erwarten, welches nicht abrupt bei Grundtypgrenzen abbricht. Ein ursprüngliches Formenkontinuum kann natürlich nachträglich durch Aussterben vermittelnder Formen unterbrochen werden. Diskontinuitäten zwischen benachbarten Grundtypen widerlegen daher die Evolutionstheorie nicht, schwächen aber, wenn sie systematisch auftreten, ihre Plausibilität. Wichtig ist hier der Nachweis sowohl von vielfältigen grundtypinternen Kreuzungen als auch einer markanten Kreuzungsgrenze zu Nachbargrundtypen. Es sollten sich also einerseits möglichst deutliche Grenzen zu Nachbargrundtypen zeigen, andererseits innerhalb von Grundtypen keine klaren Grenzen auftreten (s. o. Abb. 82).
Bisher liegen knapp 20 Grundtypstudien vor (Scherer 1993, Fehrer 1994, Neuhaus 1995), die zeigen, dass sich das Konzept bewährt hat. Zweifellos ist das eine sehr geringe Zahl und weitere Grundtypstudien wären wünschenswert. Die Kritik von Korthof (2004, 35, 3.), es handle sich nur um ein Fragment von einer Theorie, ist so gesehen richtig. Aber diese Theorie ist grundsätzlich auf weite Teile der Organismenwelt anwendbar und sie regt daher Forschung an.
Grundtypgrenzen fallen nicht generell mit einem bestimmten taxonomischen Niveau zusammen. Zwar ist der Grundtyp bei Wirbeltieren häufig mit der Familie identisch, doch muss dies nicht immer so sein, sondern muss von Fall zu Fall geprüft werden. Korthof, der sich vergleichsweise ausführlich mit dem Grundtypkonzept befasst, hat diesen Sachverhalt falsch verstanden, vermutlich durch Verallgemeinerung weniger Grundtypbeispiele (Korthof 2004, 35, 37; 1., 2. und 9). Insbesondere ist es falsch, den Menschen mit Menschenaffen in denselben Grundtyp zu stellen, nur weil beide von vielen Taxonomen in dieselbe Familie gestellt werden. Die darauf aufgebaute Kritik von Korthof (2004, 37, 9.) geht an den Aussagen des Grundtypmodells vorbei. Denn Grundtypen werden primär durch Kreuzbarkeit erkannt, nicht durch Merkmale oder anderweitig definierte Taxongrenzen (siehe dazu auch Kritikpunkte weiter unten).
Manchmal wird argumentiert, Grundtypengrenzen seien Symptome für constraints („strukturelle Zwänge“), doch ist dieser Verweis nur eine black box: Denn was ist ein constraint? Weshalb gibt es constraints? Woran werden sie erkannt? Hier lauert die Gefahr des Zirkelschlusses: Strukturelle Gleichheit (das Grundtypspezifische) wird zunächst durch constraints erklärt, die ihrerseits mit dem Hinweis auf strukturelle Gleichheit begründet würden. Rieppel (1994, 67f.) meint dazu: Solange kein kausaler Mechanismus für die Invarianz der Entwicklung gefunden würde, sei der Verweis auf „Entwicklungszwänge“ empirisch leer und nur eine Umschreibung für die strukturelle Gleichheit (vgl. auch Junker 2002, 60f.). Evolution erklärt die Ursache für constraints bislang nicht.
2.3 Wie könnte die Existenz primär getrennter Grundtypen falsifiziert werden?
Hier muss zunächst darauf hingewiesen werden, dass in historischen Fragen kaum strenge Falsifikationen möglich sind. Die Begründung dazu findet sich im Artikel |1.1.3.2 Methodik der historischen Forschung|. Dort wird gezeigt, dass sich die Wissenschaftler in der Praxis um Verifizierungen [= Bestätigungen] bemühen, indem sie nach passenden Daten suchen. Es ist nur möglich, eine Zuordnung von Daten zu dem zugrundeliegenden Ursprungskonzept herzustellen, um eine möglichst große Plausibilität der Dateninterpretation zu erzielen. Dennoch könnte das Grundtypkonzept auf der Basis der Grundtypdefinition sensu Scherer (1993) wie folgt falsifiziert werden:
- Würden Kreuzungen zwischen Menschen und Tieren zweifelsfrei gelingen, wäre das Grundtypkonzept in der vorgelegten Form nicht haltbar.
- Würden sich keine Gruppen von Lebewesen bei Anwendung des Grundtypkriteriums abgrenzen lassen, wäre das Grundtypkonzept falsifiziert.
Daten, die im Rahmen des Grundtypkonzepts plausibel gedeutet werden können, stützen diesen Ansatz:
- Hier kann vor allem an die zahlreichen Belege für die Existenz polyvalenter Stammformen eines Formenkreises verwiesen werden (vgl. dazu |0.3.2.4.1 Genetisch polyvalente Stammformen von Grundtypen|).
- Es gibt Hinweise auf eine entwicklungsbiologische Polyvalenz.
So gibt es Fälle, wo nah verwandte Formen (innerhalb derselben Grundtypen oder sogar Gattungen) dieselben Organe oder ontogenetischen Stadien auf verschiedenen ontogenetischen Entwicklungswegen erreichen (Hall 1995; vgl. Wells & Nelson 1997; Junker 2002, 53f.). Evolutionstheoretisch ist das schwer zu erklären. Das Polyvalenz-Konzept könnte hier hilfreich sein. Hier besteht großer Forschungsbedarf, der ausdrücklich durch ein Postulat im Rahmen des Schöpfungsparadigmas motiviert ist. Das „Polyvalenz-Konzept“ könnte hier auch insofern wertvoll sein, als es die Frage nach dem Zweck einer solchen Polyvalenz nahelegt. Es könnte funktionelle Gründe dafür geben: vielleicht sind manche ontogenetische Entwicklungswege unter bestimmten Umweltbedingungen besser. Das wäre für einen Designer ein Grund, diese Möglichkeit einzurichten. Solche Zwecke zu suchen ist heuristisch wertvoll.
- Die zahlreichen empirischen Belege, dass fortgesetzte Artaufspaltungen durch zunehmende Spezialisierung in evolutionäre Sackgassen führen, ist ebenfalls im Rahmen des Grundtypkonzepts plausibel deutbar (|1.3.2.4.1 Artbildung|).
Die gegenteilige Auffassung, Evolution führe durch Artaufspaltungen und Spezialisierung nicht immer in Sackgassen, wird mit dem Fossilbericht oder anderen indirekten Indizien begründet. Doch dies ist eine völlig andere Begründungsschiene. Man verlässt bezüglich der Mechanismen (um die es hier geht) den empirischen Bereich und muss zur Argumentation Makroevolution voraussetzen. Denn die Fossilabfolge als solche beweist Makroevolution nicht.
Aus diesen Punkten ergibt sich das Forschungsinteresse, Grundtypgrenzen aufzudecken sowie Variationsmechanismen und ihre Ursachen aufzuklären. Damit ist klar: Die Grundtypenbiologie regt Forschung an und wirft prüfbare Fragen auf. Außerdem ist nicht jedes Forschungsergebnis mit der Grundtypenbiologie kompatibel. Zudem sind manche Fragen für die Grundtypenbiologie interessant, die sonst nicht verfolgt werden. Daher sind folgende Behauptungen über Wissenschaft im Rahmen des Schöpfungsparadigmas falsch:
- Die Behauptung, man könne eine Aussage und zugleich auch ihr Gegenteil ableiten,
- die Behauptung, es gebe keine Falsifizierungs- oder wenigstens Prüfmöglichkeiten,
- die Behauptung, die Ergebnisse stünden von vornherein fest und Forschung würde verhindert,
- die Behauptung, Ansätze im Rahmen des Schöpfungsparadigmas seien heuristisch wertlos.
2.4 Grundtypen und Evolutionsbiologie
Nun könnte an dieser Stelle weiter eingewendet werden, alle die oben genannten Fragen könnten auch im Rahmen des Evolutionsparadigmas gestellt werden. Teilweise trifft dies in der Tat zu; so gibt es gemeinsame Interessen an der Aufklärung von Variationsmechanismen. Andere Fragen werden von Evolutionstheoretikern jedoch nicht einmal gestellt. Welcher Evolutionstheoretiker sucht nach primären Grundtypgrenzen? Nach deren Verständnis gibt es diese nicht, daher wird er auch nicht danach suchen. Vor allem aber ändert der Einwand, es gebe die gleichen Fragestellungen auch im Rahmen der Evolutionsbiologie, nichts daran, dass die oben genannten vier Behauptungen über Wissenschaft im Rahmen des Schöpfungsparadigmas falsch sind – im Gegenteil: offenkundig ist erkenntnisgewinnende Forschung unter der Vorgabe von „geschaffenen Grundtypen“ sehr wohl möglich.
2.5 Gemeinsamkeiten verschiedener Grundtypen
Verschiedene Grundtypen weisen eine mehr oder weniger große Zahl von Gemeinsamkeiten (Homologien, Synapomorphien) auf. Diese Gemeinsamkeiten sollen auf gemeinsame Vorfahren hinweisen, aus denen verschiedene Grundtypen entstanden sind. Dies stehe nach Ansicht mancher Kritiker dem Grundtypen-Ansatz entgegen. In diesem Sinne ist auch das Argument von van Dongen & Vossen (1984, 41f.) zu verstehen: „Without common descent there is no reason why the chromosomes of apes and man should be so strikingly similar as they are.“
Darauf kann wie folgt entgegnet werden: Zum einen handelt es sich hierbei um das Ähnlichkeitsargument, wonach Ähnlichkeit als Indiz für gemeinsame Abstammung gewertet wird. Das Ähnlichkeitsargument erweist sich jedoch bei genauerer Analyse als nicht stichhaltig, denn es gibt keine objektiven Kriterien zur Bestimmung abstammungsbedingter Ähnlichkeiten (|1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie|). Zum anderen wird nicht gefordert, dass sich Grundtypen in möglichst vielen Merkmalen unterscheiden. Das definierende Kreuzungskritierium hat zunächst mit Merkmalen und Ähnlichkeiten nichts zu tun. Allerdings stellt sich durchaus die Frage, ob grundtypspezifische Eigenschaften auch an geeigneten äußeren Merkmalen erkennbar sind (dazu können auch Sequenzunähnlichkeiten von Proteinen und DNA gehören). Die oben angeführte Behauptung von van Dongen & Vossen trifft auf das Grundtypkonzept jedoch nicht zu. Selbstverständlich haben verschiedene Grundtypen mehr oder weniger zahlreiche Gemeinsamkeiten (vgl. dazu Artikel |0.3.2.5 Grundtypen und Makrotaxonomie|).
Da Grundtypgrenzen nicht primär an Merkmalen festgemacht werden, geht auch der Hinweis von Korthof (2004, 37, 9.), die Chromosomenvariation innerhalb der Hominoiden sei geringer als die innerhalb der Hundeartigen (Canidae), am Grundtypkonzept vorbei. Es ist zudem zu kurz gegriffen, einzelne Merkmale oder Merkmalskomplexe herauszugreifen, andere dagegen zu übergehen. Im Bezug auf die Abgrenzbarkeit des Grundtyps Mensch heißt das: Es wird ein Gesamtbild von Gemeinsamkeiten und Unterschieden zwischen Menschen und Menschenaffen benötigt, um beurteilen zu können, ob auch auf morphologisch-anatomischer Ebene oder in anderer Hinsicht Grundtypgrenzen plausibel sind. Oben wurde darauf hingewiesen, dass es außer dem definierenden Kriterium Hilfskriterien zur Grundtypabgrenzung gibt. Im Falle des Menschen ist die Grundtypenbiologie auf diese Kriterien besonders angewiesen, da sich Hybridisierungsexperimente mit Tieren aus ethischen Gründen verbieten. Aus der Vergleichenden Biologie gibt es zahlreiche Befunde, die eine deutliche Trennung zwischen Menschen und Menschenaffen hinweisen (aus der neueren Forschung sei z. B. auf Smith 2005, Jorde 2005, Neufeld & Conroy 2004 oder Weissenbach 2004 hingewiesen).
Hinter diesen Argumenten steht noch ein grundsätzlicherer Einwand, der im Folgenden besprochen wird.
2.6 Nur manche Ähnlichkeiten sind abstammungsbedingt
Arten desselben Grundtyps gehen nach dem Grundtypkonzept auf eine gemeinsame (geschaffene) Stammart zurück, Arten verschiedener Grundtypen dagegen nicht. Dennoch weisen – so Korthof (2004, 38) – verschiedene Grundtypen vergleichbare Ähnlichkeiten auf wie Arten innerhalb desselben Grundtyps. Es gibt also keine einheitliche Erklärung für Ähnlichkeit; dies erscheint inkonsequent (vgl. Neukamm 2005). Diese Feststellung ist richtig, aber sie ist kein Gegenargument gegen das Grundtypkonzept. Auch im Rahmen des Evolutionsparadigmas gibt es keine einheitliche Erklärung für Ähnlichkeiten, auch nicht für solche, die nach vergleichend-biologischen Kriterien als homolog eingestuft werden (|1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie|). Viele nach biologischen Kriterien als homolog einzustufende Ähnlichkeiten können aufgrund widersprüchlicher Merkmalsverteilungen nicht auf gemeinsame Vorfahren zurückgeführt werden, sondern werden als Konvergenzen interpretiert. Das gilt auch für viele Komplexmerkmale (Beispiele in Junker 2002, 85ff.). Unter evolutionären Prämissen gibt es kein objektives Kriterium zur Unterscheidung von Homologie und Konvergenz (Junker 2002, Kapitel 2) und auch keine einheitliche Erklärung für (strukturell homologe) Ähnlichkeiten.
Im Rahmen der Grundtypenbiologie wird der Unterschied zwischen abstammungsbedingten und nicht-abstammungsbedingten Ähnlichkeiten an den Grundtypgrenzen festgemacht, die anhand eines von den Merkmalen unabhängigen Kriteriums – dem Kreuzungskriterium – erkannt werden. Daraus resultiert auch die Antwort auf die Frage Korthofs, weshalb zwar die Ähnlichkeiten innerhalb der Hundeartigen auf einen gemeinsamen Vorfahren zurückgeführt werden, nicht aber die Ähnlichkeiten von Hunde- und Katzenartigen. Sie lautet: Hunde- und Katzenartige gehören nach dem Grundtypkriterium der Kreuzbarkeit verschiedenen Grundtypen an.
An dieser Stelle könnte eine interessante Untersuchung ansetzen. Man könnte postulieren, dass es mindestens eine Eigenschaft oder einen Merkmalskomplex gibt, der Nachbargrundtypen voneinander unterscheidet und der nicht durch mikroevolutive Prozesse entstehen kann. Wer geht solchen Fragen nach? Nur diejenigen, die die Existenz primär getrennter Grundtypen zugrundelegen. Die Grundtypenbiologie regt Forschung an.
Daraus ergibt sich eine weitere Antwort auf die Frage, weshalb die Ähnlichkeiten verschiedener Grundtypen nicht auf Evolution zurückgeführt werden: Die betreffenden Grundtypen sind (neben evtl. zahlreichen Gemeinsamkeiten) in bestimmten Merkmalen so sehr verschieden, dass diese Unterschiede nur makroevolutiv überwunden werden könnten. Die Nichteinheitlichkeit der Deutung von Ähnlichkeit wird im Grundtypkonzept also auch mit der Mechanismenfrage begründet: Für Makroevolution fehlen graduell wirkende Mechanismen, daher sind die Ähnlichkeiten von Grundtypen, die durch makroevolutive Sprünge voneinander getrennt sind, nicht durch Evolution zu begründen, es sei denn, man nimmt größere Evolutionssprünge an. Diese würden aber dennoch nach allem, was man heute weiß, latent graduelle Änderungen erfordern, die sich im Laufe der Zeit ansammeln und dann irgendwann (z. B. durch eine homöotische Mutation) plötzlich in Erscheinung treten könnten. Für einen solchen Vorgang gibt es allerdings keine empirischen Belege. Wenn Evolution tatsächlich auch genotypisch makroevolutive Sprünge vollziehen könnte, würde sie sich freilich an dieser Stelle nicht mehr von der Grundtypenbiologie unterscheiden
An dieser Stelle sei im Vorgriff auf einen weiter unten zu besprechenden Einwand darauf hingewiesen, dass der Unterschied zwischen Mikro- und Makroevolution nicht an einer Taxongrenze, sondern an Qualitäten festgemacht wird (siehe Einwand „Erfordert die Grundtypdiversifikation Makroevolution?“ weiter unten). Die Grenze zwischen Mikro- und Makrovolution kann aber mit einer Taxongrenze zusammenfallen, die aber sicher oberhalb der Biospeziesgrenze liegt.
2.7 Darwinscher Mechanismus einer Diversifikation bis zum Familien-Level?
Korthof (2004, 36, 5.) fragt in seiner Kritik des Grundtypmodells, wie die Diversifikation [= Verschiedenwerden] innerhalb von Grundtypen geschehe und stellt fest, dass sie durch mikroevolutive Prozesse erfolgen müsse. Das ist richtig, denn nach der Grundtypenbiologie erfolgt die Diversifikation tatsächlich allein auf der Basis von Mikroevolution. Weiter behauptet Korthof, „the mechanism must be standard Darwinian mutation and natural selection. … genera and species are created by natural processes as described by the textbooks.“ Das ist nur teilweise richtig. Natürlich sind Mutation und Selektion Mechanismen der Mikroevolution und tragen zur Grundtypendiversifikation bei. Alle bekannten Variationsmechanismen sind auch Bestandteil der Grundtypenbiologie. Es mag aber sein, dass es noch weitere Mechanismen gibt, die erst noch zu entdecken sind. Daran besteht im Rahmen der Grundtypenbiologie großes Interesse, was einmal mehr deutlich macht, dass das Schöpfungsparadigma Wissenschaft nicht verhindert – im Gegenteil: Je mehr über die Variationsmechanismen bekannt ist, desto aussagekräftiger kann die Plausibilität des Grundtypmodells eingeschätzt werden.
Vor allem aber rechnet das Grundtypmodell damit, dass die Grundtypen zu Beginn ihrer Existenz polyvalent waren. Die Hauptquelle der Variabilität steckt demnach bereits in den (geschaffenen) Stammformen (vgl. |0.3.2.4.1 Genetisch polyvalente Stammformen von Grundtypen|). Es gibt Möglichkeiten, eine ursprünglich große Polyvalenz aufzudecken. Einige davon sind im Artikel |0.3.2.4.1 Genetisch polyvalente Stammformen von Grundtypen| beschrieben (ausführlicher in Junker & Scherer [2001|, Kap. VII.17.3). Auch hier dürfte es noch viel zu entdecken geben, und auch hier regt das Grundtypmodell Forschung an.
Grundtypenbiologie impliziert also (aktuelle oder potentielle) Polyvalenz der Stammformen. Dieser Aspekt scheint Korthof (2004) nicht bekannt zu sein, wenn er schreibt: „The beautiful ornamentation of the Argus pheasant … was not created by God but by selection.“ Korthofs Ansicht nach folgt dies aus dem Grundtypkonzept. Nein, selbstverständlich gehört das Potential zur Ausprägung von „Schmuckstücken“ wie Schillerfarben, Krönchen, Pfauenauge und dergleichen zum ursprünglichen Bestand der polyvalenten Stammform des Fasanenartigen-Grundtyps! Um dieses Potential auszuschöpfen, spielen Mutation, Selektion und Gendrift sicher auch eine wichtige Rolle, aber kaum die einzige. Polyvalenz schließt auch das Mutationspotential ein. Aber alleine schon die Kombinationen verschiedener Allele könnten ein erhebliches Variationspotential bergen. Abb. 96 gibt hierzu modellhaft (!) einen Eindruck. Es handelt sich dabei um die vielfältige Nachkommenschaft von äußerlich einheitlichen F1[= erste Tochtergeneration]-Mischlingen aus Pudel und Wolf (s. Abb. 213) zeigt die einheitliche Elterngeneration der in Abb. 96 gezeigten Tiere der F2-Generation). In der F2-Generation tritt die genetische Vielfalt durch unterschiedliche Kombinationen zutage. Hier wird beispielhaft sichtbar, dass eine morphologisch einheitliche Elterngeneration (hier: die F1-Generation der PuWos) in kürzester Zeit (in wenigen Generationen) eine erhebliche Vielfalt zeigen kann, wenn sie anfangs sehr polymorph [= genetisch vielseitig] war. Die PuWos der F1-Generation können als Modell (!) für einen ursprünglichen Grundtyp betrachtet werden. Im Grundtypmodell wird davon ausgegangen, dass die ursprünglichen Grundtypen noch deutlich mehr Variationspotential besaßen als die F1-Generation der PuWos.

Abb. 213: Äußerlich einheitliche F1-Mischlinge aus Pudel und Wolf, die genetisch
vielfach mischerbig sind.

Abb. 96: „PuWos“ der zweiten Generation: Die Nachkommen von Mischlingen aus Pudel und Wolf. Sie zeigen eine reiche Vielfalt von Merkmalskombinationen.
Korthof schreibt weiter: „If the common ancestor of that group did not possess the eyespots on its tail, mutation and natural selection must have created the eyespots.“ Dieser Satz stimmt zwar, aber er beschreibt eine Voraussetzung, die im Rahmen der Grundtypenbiologie gar nicht gegeben ist: „If the common ancestor … did not possess…“. Das ist die Voraussetzung der Evolutionstheorie, nicht aber der Grundtypenbiologie. Nach dem Grundtypkonzept besaß der gemeinsame Vorfahre dieses Merkmal (Polyvalenz), was allerdings nicht bedeutet, dass es phänotypisch ausgeprägt sein musste. Die Evolutionsbiologie muss erklären, wie durch Mutation und Selektion das Pfauenauge entstand und welche Selektionsbedingungen die Entstehung dieses Merkmals überhaupt ermöglichten.
Korthof (2004, 36) meint, es würde im Rahmen des Grundtypmodells keinen Sinn machen, die Effizienz der natürlichen Selektion in Frage zu stellen und zu behaupten, Selektion sei keine kreative Kraft. Diese Kritik erübrigt sich, wenn die anfängliche Polyvalenz berücksichtigt wird. Denn das Grundtypkonzept benötigt keine wirklich kreativen Faktoren, auch keine kreative Selektion. Die Vielfalt der Grundtypen steckt vielmehr aktuell oder potenziell in den ursprünglichen Grundtyp-Stammformen. Deren Ausprägung erfolgt durch Rekombination und Mutationen (und anderen Mechanismen).
Daher ist die Behauptung, dass Selektion nicht kreativ sei, nicht widersprüchlich zum Grundtypmodell. Die Kritik von Korthof hängt an dieser Stelle mit einem weiteren Missverständnis zusammen, auf welches im Folgenden Kritikpunkt eingegangen wird.
2.8 Erfordert die Grundtypdiversifikation Makroevolution?
Auch diese Kritik hat Korthof (2004, 36, 6. und 7.) formuliert. Er hält die Bezeichnung „Variation“ für die Grundtypdiversifikation für unangemessen. Doch dies ist zum einen Definitionssache. Zum anderen ist unter der Voraussetzung polyvalenter Stammformen in der Tat nicht mehr als Variation erforderlich. Und es muss auch keine Makroevolution ablaufen. Hier begeht Korthof einen schwerwiegenden Fehler, wenn Evolution oberhalb des Biospezies-Level bereits als „Makroevolution“ deklariert. Wenn er sich mit dem Grundtypkonzept auseinandersetzt, muss er jedoch berücksichtigen, dass in dessen Rahmen der Unterschied zwischen Mikro- und Makroevolution nicht an Taxongrenzen festgemacht wird, sondern an Qualitäten (Entstehung neuer Konstruktionen bzw. neuer Basisfunktionszustände; vgl. Junker & Scherer 2001, II.4.3, S. 53f. sowie IV.7.4; zu Basisfunktionszuständen siehe |1.3.4.2.1 Beispiel Bakterienmotor|), sonst diskutiert er an den eigentlichen Fragen vorbei. Hier wird übrigens sichtbar, dass es in der Auseinandersetzung um Schöpfung und Evolution auch um die inhaltliche Füllung von Begriffen geht. Die Diskussion führt in die Irre, wenn die verwendeten Begriffe nicht einheitlich definiert werden. Und es ist irreführend, wenn Begriffe so verwendet werden, dass man den Gegner besser kritisieren kann.
Wenn der Unterschied zwischen Mikro- und Makroevolution an Taxongrenzen festgemacht wird, wird das eigentliche Problem verschleiert, nämlich die evolutive Entstehung echter Neuheiten. Dies gilt insbesondere dann, wenn Entwicklungsvorgänge bereits dann als Makroevolution definiert werden, wenn sie über die Biospezies-Grenzen hinausgehen. Dann entsteht nämlich der falsche Eindruck, Makroevolution sei bereits nachgewiesen, wenn die Entstehung neuer Biospezies beobachtet wurde (Korthof 2004, 44), und es sei damit im Prinzip geklärt, wie die Evolution der Lebewesen mechanismisch verlaufen sei. Davon kann aber nicht die Rede sein.
Der eigentliche Knackpunkt ist, welche Qualität von Veränderungen durch Mutation (im Zusammenwirken mit anderen Evolutionsfaktoren) möglich sind, nämlich ob auf diesem Wege die Entstehung echter Neuheiten möglich ist. Natürlich ist hier eine möglichst genaue Bestimmung dessen erforderlich, was unter „echten Neuheiten“ zu verstehen ist. Letztlich steht die Forschung hier vor dem Problem der Quantifizierung, ein Problem, mit dem alle Beteiligten zu tun haben und an dessen Lösung auch Grundtypforscher interessiert sind – wieder ein Beispiel, dass die Grundtypenbiologie nicht aus der Forschung herausführt.
Wird die Definition von Mikro- und Makroevolution an Qualitäten festgemacht und nicht an Taxongrenzen, löst sich auch eine vermeintliche „Inkonsistenz“ des Grundtypmodells auf, die Korthof (2004, 39) wie folgt beschreibt: „Why is common descent a good explanation up to the family level and a bad explanation a higher levels?“ Antwort: Weil für ersteres Evolutionsmechanismen gefunden wurden, für zweiteres nicht, und weil an dieser Grenze häufig die Grenze zwischen Mikro- und Makroevolution liegt. (Dabei ist daran zu erinnern, dass die Grundtypgrenze nicht automatisch die Familiengrenze ist, sondern von Fall zu Fall ermittelt werden muss! Korthofs Frage müsste also lauten: Warum wird gemeinsame Abstammung innerhalb von Grundtypen als plausibel erachtet, darüber hinaus jedoch nicht? Dass hier ein Problem liegt, räumen auch manche Evolutionsbiologen ein, vgl. Leigh [1999| und Carroll [2000|.)
2.9 Hohe Artbildungsraten
Ebenfalls von Korthof (2004, 36, 8.) wird der Einwand vorgebracht, das Grundtypmodell erfordere unrealistisch hohe Artbildungsraten. Dieser Einwand erscheint gemessen am derzeitigen Kenntnisstand berechtigt, wenn man von einer jungen Schöpfung ausgeht, wonach die Organismenwelt allenfalls größenordnungsmäßig 10.000 Jahre alt ist. Mittlerweile liegt eine zunehmende Zahl von Studien vor, die zeigen, dass Artbildungsprozesse um Größenordnungen schneller verlaufen können, als im Rahmen von Evolutionstheorien früher vermutet worden war (Literaturhinweise in Junker & Scherer 2001, 291). Berühmt geworden ist die oft als „explosiv“ bezeichnete Artbildung von Buntbarschen in den ostafrikanischen Seen (Fehrer 1997) (vgl. |0.3.2.4.1 Genetisch polyvalente Stammformen von Grundtypen|). Diese Geschwindigkeit lässt sich wahrscheinlich nur auf der Basis von Polyvalenz verstehen (wobei von Evolutionstheoretikern der Zeitrahmen immer noch größer eingeschätzt wird als von Vertretern der Grundtypenbiologie). Hier liegt also zugleich auch eines der Argumente für die Polyvalenz der Grundtyp-Stammformen.
Wie schnell Artbildung ablaufen kann, ist durch weitere Forschung zu klären. Hier sind viele Fragen offen und auch hier hat die Grundtypenbiologie wieder ein ausgeprägtes Forschungsinteresse. Es deutet sich bisher an, dass diese Frage eng mit der Frage nach der Polyvalenz der Stammformen gekoppelt ist.
2.10 Die Grundtypen tauchen im Fossilbericht nicht gleichzeitig auf
Dieser Einwand von Korthof (2004, 37, 10. und 11.) trifft im Wesentlichen zu. Das zeitlich gestaffelte fossile Auftreten verschiedener Taxa mit verschiedenen Grundtypen im Fossilbericht ist in der Tat ein schwerwiegendes Problem für die biblische Schöpfungslehre, die dem biblischen Schöpfungsbericht folgend von einer gleichzeitigen Erschaffung aller Grundtypen ausgeht. Stephan (2002) diskutiert Lösungsansätze, die aber noch viele Fragen offen lassen. Man muss bei diesem Einwand von Korthof beachten: Das Grundtypkonzept an sich impliziert zunächst keine Gleichzeitigkeit der Entstehung der Grundtypen. Erst in einem zweiten Schritt kommt dann der biblische Aspekt der (annähernden) Gleichzeitigkeit ihrer Erschaffung in der Schöpfungswoche ins Spiel.
Allerdings ist innerhalb von einzelnen geologischen Systemen ein nahezu gleichzeitiger erster fossiler Nachweis von Grundtypen regelmäßig anzutreffen, besonders markant bei der kambrischen Explosion, aber auch bei vielen Tier- und Pflanzengruppen, die in jüngeren Systemen fossil überliefert sind (Valentine 2004; siehe |1.7.2.2 Kambrische Explosion|).
2.11 Die Entstehung der Grundtypen ist ein Geheimnis
Aus der Sicht eines konsequenten Naturalisten ist dieser Satz (Korthof 2004, 38) natürlich eine schwerwiegende Kritik. Im Rahmen des Schöpfungsparadigmas liegt dieser Sachverhalt dagegen in der Natur der Sache. „Schöpfung“ bedeutet nun einmal die unableitbare Herkunft von Entitäten. Eine solche Voraussetzung kann aber empirisch nicht a priori ausgeschlossen werden, vielmehr ergibt sich die Aufgabe, wie man methodisch mit der Möglichkeit einer unableitbaren Schöpfung umgeht (siehe dazu |0.1.1.2 Schöpfung und Wissenschaft|).
In diesem Zusammenhang wird auch behauptet, die Grundtypenbiologie ersetze eine bereits vorliegende gute Erklärung für die Entstehung der Grundtypen durch ein Mysterium. „But when a good explanation is available, it is unacceptable to fall back into the mysteries of pre-Darwinian times“ (Korthof 2004, 40). Diesem Zitat ist zwar zuzustimmen; die Frage ist aber, ob die Prämisse stimmt. Genau darüber geht aber die Kontroverse. Welches ist die „gute Erklärung“? Es gibt seit Darwin nur die Behauptung einer Erklärung, aber der Beweis fehlt. Darüber kann man kontrovers diskutieren (was im Evolutionsteil von Genesisnet geschieht). Aber man kann den Verfechtern der Grundtypenbiologie nicht den Vorwurf machen, sie würden Wissen ignorieren und 200 Jahre in der Forschungsgeschichte zurückfallen. Die Kritik am Grundtypmodell basiert hier demnach auf einer falschen Voraussetzung, nämlich „Darwin already had an elegant explanation for the similarities between taxonomic groups“ (Korthof 2004, 47). Die Ähnlichkeiten konnte er wohl evolutionstheoretisch deuten, aber eine Erklärung für ihre Entstehung hatte er nicht und niemand hat sie bis heute. Davon lenken viele Kritiker des Schöpfungsparadigmas ab, z. B. indem sie versuchen, den Zugang zu dieser entscheidenden Frage schon im wissenschaftstheoretischen Vorfeld abzusperren (vgl. dazu auch |1.1.3.2 Methodik der historischen Forschung|).
2.12 Ist der Test des Grundtypmodells ein Test auf Schöpfung?
In vielen Fällen wird man dieselben Befunde erwarten oder auch nicht erwarten, sowohl wenn man von Schöpfung als auch wenn man von Evolution ausgeht. Evolutionstheoretiker reklamieren viele Befunde als Bestätigungen ihrer Ansichten, die mit einer Bestätigung ihrer Theorien gar nichts zu tun haben (vgl. |1.1.3.2 Methodik der historischen Forschung|; dort wird auch festgestellt, dass das Evolutionsparadigma gegensätzliche Befunde verkraftet und dass damit alle diese Befunde nicht als eingetroffene Vorhersagen gelten können). In diesem Sinne kann man auch Tests am Grundtypmodell damit relativieren, dass dessen Bestätigung keine Bestätigung für Schöpfung sei (Neukamm 2005). Ist also Grundtypenbiologie überhaupt eine Forschungsrichtung, die im Schöpfungsparadigma anzusiedeln ist? Das wäre nur der Fall, wenn die Grundtypenbiologie zwingend aus dem (biblischen) Schöpfungsparadigma ableitbar wäre. Doch trifft dies zu?
Im Artikel |1.1.3.2 Methodik der historischen Forschung| wurde dargelegt, dass es in historischen Fragen nur um Plausibilitäten gehen kann. „Schöpfung“ kann prinzipiell nicht direkt getestet werden, genausowenig wie „Makroevolution“ direkt getestet werden kann. Wird „Schöpfung“ aber dahingehend konkretisiert, dass eine getrennte Entstehung polyvalenter Stammformen vorausgesetzt wird, dann können Befunde ermittelt werden, die diese Vorgabe stützen oder eher unplausibel erscheinen lassen. Mehr ist bei der Prüfung historischer Theorien methodisch nicht möglich, und dies ist kein Spezifikum für die Grundtypenbiologie. Getestet wird nur das, was aus dem Paradigma gefolgert wurde (vgl. Mahner 2000; Neukamm 2005). Im Artikel |0.3.1.1 Biblische Schöpfungslehre und Grundtypenbiologie| wird gezeigt, dass sich die Existenz abgrenzbarer Grundtypen aus dem biblischen Schöpfungsparadigma ergibt (jedenfalls dass ihre Existenz mindestens sehr nahe liegt). Ein auf jeder taxonomischen Ebene gleichmäßiges Formenkontinuum würde dazu nicht passen. Wenn also das Grundtypkonzept getestet wird, wird damit auch das Schöpfungsparadigma auf Plausibilität getestet.
Allerdings sind Grundtypgrenzen auch im Rahmen des Evolutionsparadigmas möglich. Es könnte evolutionäre Mechanismen geben, die dazu führen, dass es klar abgrenzbare Grundtypen gibt. Vielleicht lässt sich sogar argumentieren, dass solche Grenzen und abgrenzbare Cluster von niederrangigenTaxa zu erwarten sind (allerdings dürfte dergleichen evolutionstheoretisch nicht vorhergesagt worden sein). Kurzum: Die Existenz abgrenzbarer und in sich geschlossener Grundtypen testet nicht unbedingt spezifisch das Schöpfungsparadigma. Daher muss der Test auf Abgrenzbarkeit mit anderen Tests (s.u.) kombiniert werden.
Zunächst aber können wir zweierlei festhalten:
- Wenn sich regelmäßig nach geeigneten Kriterien deutliche Grundtypgrenzen und innerhalb dieser Grenzen enge Beziehungen nachweisen lassen, ist ein solcher Befund ein starker Beleg auf primäre Grenzen, also auf solche, die es von Anfang der Existenz der Grundtypen gab. Genau das ist eine Schlussfolgerung aus dem biblischen Schöpfungsparadigma.
- Könnte man evolutionstheoretisch tatsächlich abgrenzbare Grundtypen vorhersagen, dann würde an dieser Stelle auf das Evolutionsparadigma zutreffen, was Kritiker dem Schöpfungsparadigma vorhalten: Die Befunde könnten sein wie sie wollen: sie würden immer passen. Im Rahmen des Evolutionsparadigmas wäre bezüglich der Abgrenzbarkeit oder Nicht-Abgrenzbarkeit von Taxa alles erklärbar, und an dieser Stelle gäbe es dann auch keine Möglichkeit, das Evolutionsparadigma zu testen (vgl. |1.1.3.2 Methodik der historischen Forschung|). Man kann durchaus die komplette Grundtypforschung auch im Rahmen des Evolutionsparadigmas betreiben (wie auch umgekehrt Evolutionsforschung im Rahmen des Schöpfungsparadigmas), aber viele Fragen, denen sich die Grundtypenbiologie widmet, sind durch die Vorgabe einer allgemeinen Evolution aller Lebewesen nicht gerade motiviert. Daher zeigt die Tatsache, dass im Rahmen der Grundtypenbiologie erforschbare Fragen nur dadurch aufgeworfen werden, dass das Grundtypkonzept seinen Platz sehr wohl zurecht im Rahmen des Schöpfungsparadigmas hat.
Es gibt aber weitere Testmöglichkeiten, bei denen unterschiedliche Erwartungen vorliegen, je nachdem, ob man vom Evolutions- oder Schöpfungsparadigma ausgeht. Es geht ja in der Grundtypenbiologie nicht nur um die Frage der Abgrenzbarkeit geeigneter systematischer Taxa, sondern auch um ihre Eigenschaften und Fähigkeiten. Hier ist an das Stichwort „Polyvalenz“ zu erinnern. Wenn plausibel gemacht werden kann, dass die Lebewesen mit einem großen genetischen Potential ihre Existenz starteten (Anhaltspunkte gibt es dafür: |0.3.2.4.1 Genetisch polyvalente Stammformen von Grundtypen|), und wenn eine Abnahme der Polyvalenz im Laufe der Generationen plausibel gemacht werden kann, passt dieser Befund letztlich nicht ins Evolutionskonzept, ist aber im Rahmen des Schöpfungsparadigmas zu erwarten. Denn er würde darauf hinauslaufen, in der Vergangenheit auf zunehmend polyvalentere Formen zu stoßen. Zu primitiven Vorfahren, die im Evolutionsparadigma zu fordern sind, gelangte man auf diesem Wege nicht. Dagegen könnte der Befund der nahezu unbegrenzten Variabilität und makroevolutionären Entwicklung der Arten nicht Teil des Grundtypkonzepts sein. Eine unbegrenzte Variabilität könnte aber Teil irgendeines Schöpferplans sein, wie Neukamm (2005) richtig feststellt. Hier wird deutlich, dass zwischen allgemeinem Paradigma und konkreten Theorien im Rahmen des Paradigmas unterschieden werden muss. Denn unbegrenzte programmierte (geschaffene) Variabilität wäre für ein allgemeines Schöpfungsparadigma vorstellbar, verbietet sich aber aus dem biblischen Befund und passt nicht zum Grundtypmodell.
Man kann weiter argumentieren: Wenn gezeigt werden könnte, dass Evolution nur im Rahmen von Grundtypen möglich ist, dann wäre Deszendenz zwischen den Grundtypen widerlegt. Ein solcher Nachweis ist allerdings sehr schwierig, strenggenommen nicht zu führen. Denn ein „nicht möglich“ kann immer als ein „noch nicht möglich“ interpretiert werden. Auch hier kann man wieder nur mit Plausibilitäten arbeiten: Wenn trotz aller Forschungsbemühungen keine Anhaltspunkte erkennbar werden, wie Grundtypgrenzen durch natürliche Prozesse überschritten werden können, wird eine primäre Abgrenzbarkeit von Grundtypen immer wahrscheinlicher.
Die Problematik der logischen Verknüpfung des biblischen Schöpfungsparadigmas und daraus gefolgerter Hypothesen soll durch einen Vergleich mit der Situation im Rahmen des Evolutionsparadigma verdeutlicht werden. Neukamm (2005) meint im Sinne der eingangs dieses Abschnitts genannten Kritik, dass das Schöpfungsparadigma mit keinem Postulat und mit keiner Folgerung des Grundtypmodells in logischer Hinsicht verbunden sei. „Keine der Hypothesen des Grundtypmodells lässt sich aus der Schöpfungshypothese ableiten oder schlüssig begründen.“ Wird lediglich ein allgemeines Schöpfungsparadigma zugrunde gelegt, so trifft diese Einschätzung Neukamms zu. Das gilt aber auch für das Evolutionsparadigma und sogar für manche untergeordneten Theorien im Rahmen dieses Paradigmas. In seinem Beitrag nennt Neukamm (2005) das Beispiel der abgestuften Ähnlichkeit: „So lässt sich z. B. aus dem Darwinschen Postulat, dass alle Arten von einem gemeinsamen Vorfahren abstammen (in Verbindung mit dem Wissen um die Vererbung, Variation und Selektion) die Folgerung ableiten, dass zwischen den Arten eine abgestufte Ähnlichkeit bestehen muss; eine Folgerung, die man testen kann und die bis zur molekularen Ebene immer wieder bestätigt wird“ (Hervorhebung im Original). In zahlreichen Fällen lassen sich die Organismengruppen jedoch nur mit sehr vielen Merkmalswidersprüchen in Baumdiagrammen darstellen, in einer Reihe von Fällen (sowohl in der Mikro- als auch in der Makrotaxonomie) ist die Baumdarstellung kaum sinnvoll (vgl. beispielhaft Abb. 206, 207 und 201 (s. u.) sowie |0.3.2.5 Grundtypen und Makrotaxonomie|).

Abb. 206: Ähnlichkeitsbeziehungen unter Lebewesen können als Netzwerk dargestellt werden: Verwandtschaftsverhältnisse der Arthropoden (Gliederfüßer). Je nach zugrunde gelegten Merkmalen ergeben sich unterschiedliche Gruppierungen. Die entsprechenden Untersuchungen stammen alle aus den 1990er Jahren. (Nach Wägele 2001, 102). Wägele kommentiert diese Abbildung: „Ergebnisse neuerer Analysen sind untereinander nicht kompatibel, mehrere davon oder alle müssen demnach fehlerhaft sein.“

Abb. 207: Netzartige Ähnlichkeitsbeziehungen bei Devonpflanzen. Ähnlichkeitsbeziehungen als Netzwerk am Beispiel des vielfältigen Beziehungsgefüges devonischer Landpflanzengruppen aufgrund unterschiedlicher gemeinsamer Merkmale. (Nach Junker 1996. Dort werden die Details erläutert und sind weitere Beispiele dieser Art abgebildet)
Abb. 210: Bunt gemischtes Merkmalsspektrum. Das Schnabeltier ist eine ausgeprägte Mosaikform. Seine Merkmalsverteilung erscheint bunt zusammengewürfelt. Evolutionstheoretisch muß vielfache Konvergenz angenommen werden.
Die Folgerung der abgestuften Ähnlichkeit hat sich keineswegs immer wieder bestätigt; und molekulare Dendrogramme [= Ähnlichkeitsbäume] passen oft nur teilweise (wenn überhaupt) zu den Stammbäumen, die auf der Basis morphologisch-anatomischer Merkmale ermittelt wurden. Vor diesem Hintergrund kann man hier kaum sagen, dass bestimmte Hypothesen eine logische Deduktion aus dem Evolutionsparadigma darstellen (vgl. auch |1.1.3.2 Methodik der historischen Forschung|). Es liegt offenbar in der Natur historischer Theorien, dass konkrete Schlussfolgerungen aus dem zugrundliegenden Paradigma kaum möglich sind. Die diesbezügliche Kritik am Verhältnis zwischen dem Grundtypkonzept und dem Schöpfungsparadigma trifft also nicht spezifisch das Schöpfungsparadigma, sondern alle Ursprungsparadigmen.
Schlussfolgerungen. Wird durch Prüfung des Grundtypmodells auch das Schöpfungsparadigma getestet? Strenggenommen ist dies nicht der Fall. Denn wenn wir uns auf der Ebene des allgemeinen Schöpfungsparadigmas bewegen, gibt es keine eindeutigen Vorhersagen und damit auch keine strikten Testmöglichkeiten. Dies gilt aber auch für das Evolutionsparadigma (|1.1.3.2 Methodik der historischen Forschung| ). Die spezifischeren Hypothesen der Grundtypenbiologie können jedoch durch empirische Befunde getestet werden, da aus ihnen keine beliebigen Erwartungen folgen. Da das Grundtypmodell im Rahmen des Schöpfungsparadigmas eine naheliegende Konkretisierung von Schöpfung bedeutet, erhöhen erfolgreiche Tests der Grundtypenbiologie die Plausibiliät des dahinterstehenden Schöpfungsparadigmas.
2.13 Literatur
Carroll R (2000) Towards a new evolutionary synthesis. Trends Ecol. Evol. 15, 27-32.
Fehrer J (1994) Schnelle Mikroevolution beim Grundtyp der Kraniche? Stud. Int. J 1, 38-39.
Fehrer J (1997) Explosive Artbildung bei Buntbarschen in ostafrikanischen Seen. Stud. Int. J. 4, 51-55.
Fehrer J (2000) Von der Schwierigkeit, Evolution zu rekonstruieren. Stud. Int. J. 7, 30-31.
Hall BK (1995) Homology and Embryonic Development. Evol. Biol. 28, 1-37.
Jorde LB (2005) Where we‘re hot, they‘re not. Science 308, 60-62.
Junker R (2002) Ähnlichkeiten, Rudimente, Atavismen. Holzgerlingen.
Junker R & Scherer S (2001) Evolution – ein kritisches Lehrbuch. Gießen.
Korthof G (2004) Common Descent. It’s All or Nothing. In: Young M & Edis T (eds) Why Intelligent Design Fails. A Scientific Critique of the New Creationism. New Brunswick, pp. 32-47.
Leigh EG (1999) The modern synthesis, Ronald Fisher and creationists. Trends Ecol. Evol. 14, 495-498.
Mahner M (2000) Stichwort Theorie. Naturwiss. Rdsch. 53, 157-158.
Neufeld AH & Conroy GC (2004) Human head hair is not fur. Evol. Anthropol. 13, 89.
Neuhaus K (1995) Die Familie der Bromeliaceen – ein oder mehrere Grundtypen? Stud. Int. J. 2, 15-19.
Neukamm M (2005) Die Grundtypenbiologie in der Kritik. Schöpfungstheorien und ihre prüfbaren Modelle. [#LINK 3 grundtyp.html| (Zugriff am 14. 3. 2005)
Rieppel O (1994) Homology, topology, and typology: the history of modern debates. In: Hall BK (ed) Homology: The hierarchical basis of comparative biology. San Diego, pp 63-100.
Scherer S (1993) basic types of life. In: Scherer S (Hg) Typen des Lebens. Studium Integrale. Berlin, S. 11-30.
Smith N (2005) Don‘t talk to the animals. Nature 434, 702-703.
Stephan M (2002) Der Mensch und die geologische Zeittafel. Holzgerlingen.
Valentine JW (2004) On the origins of Phyla. Chicago.
van Dongen PAM & Vossen JMH (1984) Can the theory of evolution be falsified? Acta Biotheoretica 33, 35-50.
Wägele JW (2001) Grundlagen der Phylogenetischen Systematik. München (2. Aufl.).
Weissenbach J (2004) Differences with the relatives. Nature 429, 353-355.
Wells J & Nelson PA (1997) Homology – a concept in crisis. Origin & Design 18, 12-21.
Autor: Reinhard Junker, 07.07.2005
© 2005, https://www.genesisnet.info/schoepfung_evolution/e1246.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/
0.3.3.1 Mosaikformen als Grundtypen und Baukastensysteme
Die Merkmalsverteilung verschiedener Grundtypen und höherer systematischer Gruppen sind oft so verteilt, dass netzartige – weniger baumförmige – Beziehungen resultieren. Die verschiedenen Grundtypen scheinen nach einem „Baukastensystem“ konstruiert zu sein.
1.0 Inhalt
In diesem Artikel wird erklärt, was unter „Mosaikformen“ zu verstehen ist und wie die Existenz von Mosaikformen im Rahmen des Grundtypmodells der Schöpfungslehre interpretierbar ist.
1.1 Was sind „Mosaikformen“?
Im Artikel |1.7.2.1 Definitionen| von |1.7.2.0 Mosaikformen/Übergangsformen| wird der Unterschied zwischen dem beschreibenden Begriff „Mosaikform“ (oder „Zwischenform“) und dem interpretierenden Begriff „Übergangsform“ erläutert. „Mosaikformen“ sind Lebewesen, die eine Kombination von Merkmalen besitzen, welche sonst in der Regel zu verschiedenen Gruppen von Lebewesen gehören. Eine exakte Definition dieses Begriffes ist etwas aufwendig. Daher soll im Folgenden umschrieben werden, was damit gemeint ist: Die meisten Tier- und Pflanzengruppen lassen sich anhand mehrerer typischer Merkmale charakterisieren. So haben alle Säugetiere z. B. Haare, Milchdrüsen oder nur ein Gehörknöchelchen, niemals aber Federn. Ein Säugetier mit Federn wäre eine Mosaikform, da Federn für Säugetiere nicht typisch sind, wohl aber für Vögel (wenn auch neuerdings wahrscheinlich nicht mehr ausschließlich, da befiederte Dinosaurier gefunden wurden). Die meisten Säugetiere haben auch einen bezahnten Kiefer; man kann sagen, dass dieses Merkmal ebenfalls typisch für Säugetiere ist, aber nicht ausnahmslos. Das Schnabeltier (Abb. 18) besitzt als Säugetier nämlich einen breiten Hornschnabel. Einen ähnlichen Hornschnabel besitzen aber auch die Entenvögel; für diese ist ein solcher Hornschnabel typisch. Das Schnabeltier ist wegen des Besitzes eines Hornschnabels eine Mosaikform. Außerdem legt das Schnabeltier Eier; das ist vogel- oder reptiltypisch, aber nicht säugertypisch. Der Mosaikcharakter wird schon mit diesen wenigen Merkmalen deutlich.

Abb. 18: Das Schnabeltier
1.2 Mosaikformen als Grundtypen
Viele Grundtypen – ob heute lebend oder fossil – weisen ausgeprägte Merkmalsmosaike auf. Der Mosaikcharakter von Grundtypen widerspricht nicht einer Deutung des Grundtypkonzepts im Sinne der Schöpfungslehre. Entscheidend für das Grundtypkonzept ist die Abgrenzbarkeit von Grundtypen, und zwar in erster Linie aufgrund der Kreuzbarkeit bzw. Kreuzungsbarrieren, und in zweiter Linie (bzw. wenn wenig Kreuzungsversuche durchgeführt wurden oder bei Fossilien) anhand bestimmter komplexer morphologischer [= die Gestalt bzw. den äußeren Bau betreffend] Merkmale. Das Grundtypkonzept lässt grundsätzlich verschiedenste Merkmalskombinationen bei verschiedenen Grundtypen zu.

Abb. 100: Widersprüchliche (inkongruente) Verteilung von Merkmalen A-D. Häufig sind Merkmale so verteilt, dass in einer Baumdarstellung eine mehrfach unabhängige Entstehung baugleicher Merkmale (hier B und C) angenommen werden muss.
Es ist sogar oft so, dass Mosaikformen weniger im Rahmen des Grundtypkonzepts als im Rahmen der Evolutionstheorie Probleme bereiten. Abb. 100 zeigt die Situation an einem theoretischen Beispiel. Vier Grundtypen sind darin als dicke Punkte mit kleinen Buchstaben a, b, c und d dargestellt. A, B, C und D seien komplexe (!) Merkmale, durch die sich die Grundtypen unterscheiden. (Diese Merkmale sollten mikroevolutiv nicht ineinander überführbar sein.) In diesem Beispiel sind die Grundtypen b und c Zwischenformen zwischen a und d bzw. können als Mosaikformen angesehen werden. Dennoch sind sie aufgrund ihrer besonderen komplexen Merkmale jeweils voneinander als Grundtypen abgrenzbar.
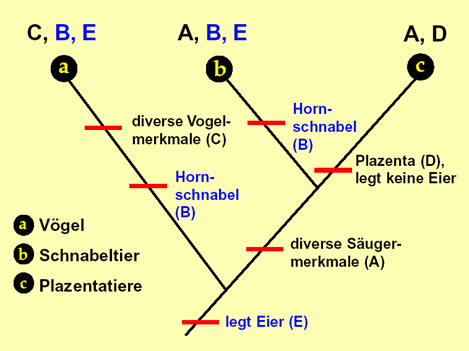
Abb. 101: Ein sehr vereinfacht dargestelltes konkretes Beispiel zu der in Abb. 100 präsentierten theoretischen Situation einer widersprüchliche (inkongruente) Verteilung von Merkmalen A-D.
Diese theoretische Situation kann auf das Schnabeltier angewendet werden. Abb. 101 zeigt dies in sehr vereinfachter Form, um den Grundgedanken deutlich zu machen. a stehe für Vögel, c für plazentabesitzende Säugetiere. Das Schnabeltier b steht in gewissem Sinne dazwischen; es ist zwar ein Säugetier (es besitzt Milchdrüsen und Haare, dafür steht A), es hat jedoch keine Plazenta (D), sondern legt Eier (E). Mit den Vögeln hat es außerdem den Hornschnabel gemeinsam (dafür steht B). Damit ist das Schnabeltier einerseits in gewisser Hinsicht eine Zwischenform zwischen Vögeln und Säugetieren, bzw. es ist eine Mosaikform mit Merkmalen, die teils typisch für Vögel, teils typisch für plazentabesitzende Säugetiere sind. Andererseits ist aber klar gegen andere Vogel- und Säugergrundtypen abgegrenzt.
Verschiedene Grundtypen können also bunte Merkmalsmosaike aufweisen. Entscheidend für das Grundtypkonzept im fossilen Bereich oder wenn keine Kreuzungsergebnisse bekannt sind ist nur die deutliche Abgrenzbarkeit bezüglich wenigstens einem komplexen Merkmal.
Dagegen bereiten solche Mosaikformen Probleme für das Evolutionsmodell. Denn man kommt hier nicht umhin, eine mindestens zweimalige unabhängige Entstehung eines Hornschnabels anzunehmen. Dies führt in die Problematik der Konvergenzen, die im Artikel |1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie| behandelt wird.

Abb. 102: Stark vereinfachte Darstellung einer netzförmigen Beziehung am Beispiel von Vögeln, Schnabeltier und plazentabesitzenden Säugetieren.
1.3 Baukastensystem
Man kann ein „Baukastensystem“ als Grundlage für die Merkmalsverteilungen hypothetisch zugrundelegen. Das heißt: Die Lebewesen sind nach einem gedachten Baukastensystem konstruiert. Einzelne Merkmalskomplexe können daher in verschiedenen Grundtypen sehr unterschiedlich kombiniert sein. Dies würde sich in der Darstellung der Ähnlichkeitsbeziehungen in Form eines Netzwerks (Abb. 102) äußern. Eine solche Netz-Darstellung ist häufig gut begründet (viele Beispiele finden sich im Artikel |1.3.5.1 Ähnlichkeiten in der Morphologie und Anatomie| im dortigen |1.3.5.1.3 PDF|.). Dagegen würden Baumdarstellungen (die evolutionär als Stammbäume interpretierbar wären) Inkongruenzen beinhalten. D. h.: je nach zugrundegelegtem Merkmal bzw. Merkmalskomplex ergeben sich verschiedene Verwandtschaftsbeziehungen. In einer Stammbaumdarstellung äußert sich diese Situation im Auftreten von Konvergenzen [= mehrfach unabhängiges Auftreten von Merkmalen] oder Reversionen [= Rückentwicklungen zu einem angenommenen früheren Zustand].
Autor: Reinhard Junker, 10.01.2004
© 2004, https://www.genesisnet.info/schoepfung_evolution/i1261.php
Zurück zur Artikel-Übersicht: https://www.wort-und-wissen.org/publikationen/genesisnet/